Abstract
We investigated dietary variation during three winters in female Svalbard reindeer (Rangifer tarandus platyrhynchus) on the basis of rumen samples from animals culled in early and late winter during 2000–2002. The winter diet was compared to a late summer diet on the basis of samples from 1999, with variations in dietary composition in early and late winter among years also investigated. We conducted our assessment by identifying plant fragments in rumen samples using a point identification method. The effects of season and year on dietary composition of females were tested using non-parametric multifactorial MANOVA. During early winter, the diet seemed to be a function of both plant availability and plant quality, with the latter also highly important for the summer diet. During late winter, restricted forage availability due to snow cover seemed to have influenced dietary composition. We also found dietary variation among years both in early and late winter. We hypothesize that climatic effects previously found on the growth rate of Svalbard reindeer populations may be mediated through interannual variation in their diet where increased summer precipitation and winter rain may affect the dietary composition through decreased availability of forage biomass.
Introduction
The Arctic is characterized by extreme seasonality with long cold winters and short summers. The combination of low temperatures, low precipitation, low soil nutrient status, and short growing season serves to limit plant growth (CitationCrawford, 1989) and the amount of plant biomass is small (CitationWalker et al., 2002). These limitations impose strong pressure on arctic herbivores to use vegetation in a highly efficient manner to meet their energy requirements for survival, maintenance, and reproduction (CitationWhite, 1983).
The phenology of a plant depends on the start and the length of the snow-free period in its habitat (CitationKudo, 1991). Generally, fast-growing plants in early phenological stages have high digestibility and high nitrogen content per unit dry weight (CitationJefferies et al., 1994). During the growing period, a plant's biomass increases while digestibility (CitationStaaland, 1986) and nitrogen concentration decrease (CitationChapin et al., 1975; CitationStaaland, 1986; CitationAlbon and Langvatn, 1992; CitationVan der Wal et al., 2000b). Plant quality is therefore negatively correlated with plant quantity within species. For herbivores, there is consequently a trade-off between the amount of forage obtained (i.e. quantity) and the nutritional value and digestibility (i.e. quality) of the forage. A plant's phenological state can have a significant impact on cervid body weight as they gain from a prolonged period with access to plants in early phenological stages (CitationAlbon and Langvatn, 1992; CitationMysterud et al., 2001), a time which is consistent with peak nitrogen levels (CitationChapin et al., 1975). In temperate zone ungulate populations, seasonal migration is a common strategy to prolong the ability to obtain forage with high levels of protein (CitationAlbon and Langvatn, 1992). Svalbard reindeer (Rangifer tarandus platyrhynchus) on the other hand, are regarded as non-migratory (CitationStaaland and Røed, 1986; CitationTyler and Øritsland, 1989). During a three-month summer period, these animals must consume enough forage to replenish body reserves that are of critical importance for winter survival (CitationReimers et al., 1982; CitationTyler, 1986). However, body reserves that are built up during the summer alone are not sufficient for winter survival (CitationTyler, 1986) and fetus development (CitationTyler, 1987). Winter diet is therefore important in the life history of Svalbard reindeer.
Previous studies of Svalbard reindeer indicate that they select forage by discriminating among plant species to eat during summer (CitationEkern and Kildemo, 1978; CitationPunsvik et al., 1980; CitationStaaland and Punsvik, 1980; CitationStaaland, 1986) and the vegetation types to visit during both summer (CitationPunsvik et al., 1980; CitationStaaland and Punsvik, 1980; CitationStaaland et al., 1993; CitationHenriksen et al., 2003) and winter (CitationPunsvik et al., 1980; CitationStaaland and Punsvik, 1980; CitationStaaland et al., 1993; CitationLindner, 2002). Unlike cervids in temperate regions, which seem to select for highest plant quality (CitationAlbon and Langvatn, 1992; CitationLangvatn and Hanley, 1993), CitationVan der Wal et al. (2000b) found that Svalbard reindeer select for biomass before quality in terms of nitrogen content during the growing season, when forage availability is high. This may be the outcome of generally low levels of plant biomass (CitationBrattbakk, 1986) and high plant quality (CitationStaaland et al., 1983) during the arctic growing season. However, less is known about habitat use and forage selection during the eight- to nine-month-long High Arctic winter season, although snow characteristics and distribution seem to be the main factors determining the use of different vegetation types by Svalbard reindeer (CitationLindner, 2002).
In this article we focus on the winter diet of female Svalbard reindeer by analyzing rumen samples from culled animals. Additional data on late summer diet were used for comparison. During the growing season the availability of plants with high biomass and/or quality is high. During autumn and early winter, before snow has started to accumulate, the availability of forage is relatively high, although plant quality has decreased. In late winter, plant quality is low and snow and ice cover most of the ground, making plants less accessible. However, when snow starts to melt early in exposed habitats, nutritious plants that already have started to grow may be available.
Since the availability and quality of arctic plant species vary greatly through the year, we tested whether there is a different diet in early winter compared to late winter, and compared early and late winter diet with late summer diet. When the availability of forage is high, as is true during the growing season and early winter, high quality species and high biomass species are expected to dominate in the diet. When the availability of forage is low, as is the case during late winter, species growing in exposed habitats are expected to dominate in the diet. In addition, climatic conditions vary greatly among years, which may have consequences for forage availability. The phenomenon of ground icing or rain-on-snow and its effect on reindeer survival has been described by several authors (CitationReimers, 1982; CitationAanes et al., 2000; CitationSolberg et al., 2001), but the reports are mainly anecdotal, and little has been recorded about how ground icing influences foraging behavior. Therefore, we investigated whether there was an interannual variation in dietary composition, and discussed this variation in relation to meteorological data.
Methods
Study Area
The study area includes Colesdalen with adjacent side valleys on the Nordenskiöld Land peninsula (78°N, 15°E) on the western part of Spitsbergen, Svalbard. This part of Svalbard is influenced by a mild ocean current from the south, and has therefore a long period free of sea ice in late summer/early autumn (CitationElvebakk, 1997). The topography is of great importance as it influences snow distribution and therefore the availability of food for reindeer during winter. There is no detailed vegetation map that covers all of Svalbard, but CitationVan der Wal et al. (2000a) provided a general plant community description for the study area. Ridge communities are mostly snow free during winter, have low species diversity, and are characterized by the presence of the dwarf shrubs Dryas octopetala and Salix polaris. Extensive heathland areas occur in two forms. Luzula heaths are dominated by Luzula arcuata subsp. confusa and S. polaris, with Alopecurus borealis also found in more moist areas, forming Luzula-Alopecurus heaths. Patches of graminoid vegetation are found on shallow slopes, including Poa pratensis subsp. alpigena, P. alpina, Festuca rubra subsp. arctica, and A. borealis. Moist moss vegetation is characterized by A. borealis and Equisetum arvense, whereas wet areas are dominated by Dupontia spp. and Eriophorum scheuchzeri. Lichens are of minor importance, possibly due to overgrazing in earlier years (CitationVan der Wal et al., 2001).
Study Species
The Svalbard reindeer is highly suitable for studying simple biological mechanisms, as it is an endemic subspecies considered to be sedentary in a simple ecosystem. It is the only large herbivore present on Svalbard, but small herbivores, such as several goose species and the Svalbard ptarmigan (Lagopus mutus hyperboreus), among others, are widespread (CitationJónsdóttir, 2005). Although the Svalbard reindeer does not suffer from insect harassment or true predation, parasitic nematodes are suggested to have a strong impact on reindeer populations (CitationAlbon et al., 2002). In addition, approximately 300 animals are culled annually by local hunters and for research. Furthermore, the population size is strongly limited by climatic conditions (CitationAanes et al., 2000, Citation2002, Citation2003; CitationSolberg et al., 2001), especially during winter, as the snow depth and hardness restrict forage availability. It has been shown that the amount of precipitation during winter (CitationAanes et al., 2000; CitationSolberg et al., 2001) and ice crust formation (CitationReimers, 1982; CitationAanes et al., 2000; CitationSolberg et al., 2001) are negatively correlated with population growth rates. The total population on Svalbard was approximately 11,000 animals in 1982/1983, but population size fluctuates from year to year (CitationØritsland and Alendal 1986). Together with the adjacent Semmeldalen and Reindalen valleys, the study area supports one of the largest populations of Svalbard reindeer, with a population size that fluctuated between 300 and 665 animals during the period 1979–1999 (CitationSolberg et al., 2001).
Rumen Samples
For the winter diet, rumen content was sampled in October and April/May from 50 female Svalbard reindeer that were culled during 2000–2002 for scientific purposes. Twenty-six rumen samples were collected in October, which represents early winter, when a thin snow cover often occurs. Twenty-four rumen samples were collected in April/May, which represents late winter, which is characterized by a thicker and denser snow cover, except on exposed ridges. Even though culling took place in different years, rumen samples were collected approximately at the same time within each season. Early winter samples were collected between 19 and 26 October and late winter samples between 27 April and 3 May. For comparison with late summer diet we used 22 rumen samples collected in Colesdalen and the connected Semmeldalen between 27 July and 4 August 1999. Because we were not able to compare the late summer diet with the early and late winter diet within years, the late summer samples were excluded from the statistical analyses.
Animals were gutted immediately after shooting and the rumen was cut open. Then 300–500 mL of rumen content was collected by sampling spoonfuls from various locations in the rumen. The samples were kept frozen at −20°C for later analysis.
To determine the botanical composition of the rumen samples, we used a point identification method (CitationGolley and Buechner, 1968; CitationEkern and Kildemo, 1978; CitationStaaland and Punsvik, 1980; CitationAlbon and Langvatn, 1992; CitationStoreheier et al., 2002). We thawed each sample and homogenized it by stirring before we spread out smaller subsamples on a circular sieve (diameter of 19 cm, mesh size of 2 mm), which was washed for approximately one minute. A 1.6 cm grid was marked on the sieve, which was used to guide 100 plant fragment identifications at the grid intersections through a 0.63×–5.00× stereo microscope and a 10×–20× microscope if necessary. For each rumen sample, we analyzed 4 subsamples with approximately 50 mL rumen content resulting in 400 fragment identifications. We reported the dietary composition as percentages of forage species or species groups observed in rumen samples. Due to differences in plant digestibility and throughput time, actual dietary composition may be different to a certain extent; however, such analyses provide a good indication of the importance of different species or species groups in the diet (CitationGaare et al., 1977).
We based the identification of plant fragments in the rumen samples on comparisons with identified plant material collected close to the study area during August 2003. This collection contained dried plants, plants stored in 96% ethanol, and microscope slides of each plant's epidermis. We identified each fragment preferably to species level. In case we had problems making such precise identification, we identified fragments to genus, family, or growth form category. The growth form categories we used were graminoids, forbs, evergreen cushion plants, deciduous shrubs, evergreen shrubs, sphenophytes (i.e. horsetails—Equisetum), bryophytes, and lichens. No attempt was made to identify lichens to either species or genus due to their low occurrences. However, the genus Stereocaulon was identified and separated from other lichens as it emerged repeatedly in the rumen samples. Neither fungi nor algae were observed. Some species/genera were hard to distinguish on a macroscopic level, and were therefore pooled into groups (e.g. Festuca rubra subsp. arctica, Poa arcticum subsp. vivipara, and Poa were all included in “F. rubra/Poa”). We also recorded items such as insect eggs, bird feathers, rocks, and hairs from reindeer that were “hit” by an intersection in the sieve grid. This meant that 0.2% ± 0.5% of the fragments were classified as “other items” and 6.6% ± 1.3% of the plant fragments remained unidentified. Nomenclature follows CitationElvebakk and Prestrud (1996).
Statistical Analyses
To test whether the botanical composition of the winter diet varied among seasons (S, fixed) and years (Y, random), we performed distance-based, non-parametric multifactorial MANOVA based on Euclidian distances (CitationLegendre and Anderson, 1999; CitationAnderson, 2001) using the software DISTLM. DISTLM uses permutations of data to obtain the distribution of the test statistics, so-called pseudo F-values, which are non-parametric multivariate analogues to Fisher's F-ratios (CitationAnderson 2004). Season and year, both nominal factors, were represented by orthogonal dummy variables constructed by the software XMATRIX (CitationAnderson, 2003), transforming the multifactorial MANOVA to multivariate, multiple regression analyses. For each test we performed 999 permutations of residuals under the full model, i.e. permutations of residuals obtained after fitting the regression model to the data (CitationAnderson and ter Braak, 2003). To test for effects of season and year on dietary composition, we based the analyses on the following linear model,
1 where yi,jk
is the percentage of species i in the diet in season j and year k. Terms combining two factors represented interaction effects. We found that the interaction was significant (MANOVA: pseudo-F = 8.84, df = 2/49, p < 0.001). We therefore split the data with respect to both factors and reanalyzed them according to adjusted models without the split factor and the interaction term. We further applied family-wise Bonferroni correction (CitationUnderwood, 1997, p. 333) for adjusting the significance levels.
Results
Identified Plant Species and Species Groups
In the rumen sample analysis, 23 different plant species from 20 genera were identified in addition to 9 genera of plants and lichens where identifications at the species level were not achieved. Altogether, 60.2% ± 15.9% of the fragments could be identified as one of these 32 taxa while 33.0% ± 16.0% of the fragments could be identified as belonging to one of six more integrated groups encompassing several genera. Graminoids constituted the most frequent plant group (52.3% ± 18.0%), followed by deciduous shrubs (14.5% ± 9.4%), bryophytes (13.2% ± 11.6%), and forbs (8.3% ± 9.4%). Evergreen shrubs (subsequently called Dryas octopetala because this was the only species in the group) (1.5% ± 3.0%), sphenophytes (subsequently called Equisetum) (1.3% ± 1.5%), lichens (1.3% ± 1.4%), and evergreen cushion plants (subsequently called Saxifraga oppositifolia) (0.9% ± 3.4%) were observed less frequently.
Seasonal Variation in Dietary Composition
There was a difference in the botanical composition of the rumen samples between early winter (October) and late winter (April/May) during 2000 (one-way MANOVA: pseudo-F = 22.55, df = 1/16, p < 0.001), 2001 (one-way MANOVA: pseudo-F = 13.30, df = 1/16, p < 0.001), and 2002 (one-way MANOVA: pseudo-F = 35.60, df = 1/12, p < 0.001).
The largest recognizable parts of the rumen samples from the early winter consisted of graminoids (63.7% ± 9.9%) (; in Electronic Appendix [available free of charge at MetaPress: http://instaar.metapress.com/content/120707 or at BioOne: http://www.bioone.org/perlserv/?request=get-archive&issn=1523-0430]). F. rubra/Poa (21.3% ± 10.1%) had the highest proportion, and A. borealis (7.6% ± 4.4%) was also found frequently. The presence of S. polaris (11.2% ± 5.8%) and various bryophytes (10.6% ± 4.5%), with Polytrichum (6.2% ± 2.3%) as the most frequent genus, was relatively high in the early winter diet. In addition, Stellaria longipes (2.0% ± 2.0%), Saxifraga oppositiofolia (2.3% ± 5.4%), and Equisetum (1.3% ± 1.5%) occurred infrequently.
Figure 1 Percentages of species/species groups ± SD across individuals in the diet in the early winter (n = 26), the late winter (n = 24), and the late summer (n = 22) based on rumen samples from female Svalbard reindeer (Rangifer tarandus platyrhynchus) culled during 1999–2002. The different growth forms are displayed using different shadings; graminoids (from left), forbs and evergreen cushion plants, deciduous and evergreen shrubs, sphenophytes, bryophytes, lichens, and unidentified plant fragments.
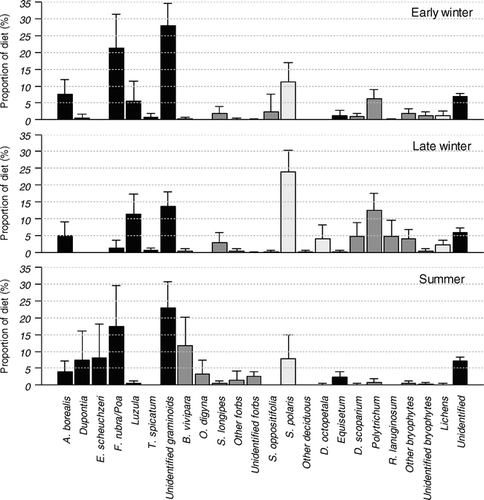
In the late winter samples, the same dominant species/species groups were found, but there were relatively more S. polaris (23.9% ± 6.5%) and bryophytes (26.4% ± 8.9%), and less graminoids (32.3% ± 9.1%) (; in Electronic Appendix [available free of charge at MetaPress or BioOne]). Within the graminoid group, there was a switch from F. rubra/Poa to Luzula (11.5% ± 5.9%). Other species infrequently present were S. longipes (2.3% ± 3.0%), Dryas octopetala (4.2% ± 4.1%), and the bryophytes Racomitrium lanuginosum (4.8% ± 4.8%) and Dicranum scoparium (4.7% ± 4.3%). Lichens (2.4% ± 1.4%) made only a small contribution to the late winter diet, although they were more frequent than in the early winter samples (1.3% ± 1.3%).
In the late summer samples, graminoids (60.7% ± 13.8%) were prevalent, with F. rubra/Poa (17.5% ± 12.0%) having the highest proportion, as was found in the early winter diet (; in Electronic Appendix [available free of charge at MetaPress or BioOne]). While Luzula (0.6% ± 0.7%) made hardly any contribution to the late summer samples, Eriophorum scheuchzeri (8.2% ± 10.0%) and Dupontia (7.4% ± 8.7%) were found frequently, which is contrary to what was found in the early and late winter samples, where this species and genus were virtually absent. Although graminoids dominated almost as much as in the early winter samples, forbs (19.6% ± 9.4%) represented a much larger proportion in the late summer samples compared to the early (2.6% ± 2.3%) and late (4.0% ± 3.0%) winter samples. Among the two dominant forb species, Bistorta vivipara (11.8% ± 8.3%) was consistently found while Oxyria digyna (3.2% ± 4.2%) represented a high percentage in a few samples. S. polaris (7.9% ± 6.9%) also represented a high proportion in the late summer samples, although not as high as in the winter samples. Equisetum (2.4% ± 1.7%) represented a small, but still higher contribution than in the winter samples, while bryophytes (1.9% ± 2.2%) and lichens (0.2% ± 0.4%) were less frequent in the late summer samples than in the winter samples.
Interannual Variation in The Winter Diet
Interannual variation in the botanical composition of the rumen samples was found for both early winter (one-way MANOVA: pseudo-F = 13.16, df = 2/23, p < 0.001) and late winter (one-way MANOVA: pseudo-F = 4.15, df = 2/21, p < 0.001). As a measure of among-year variation in botanical composition we calculated
2 for early winter samples and late winter samples separately. This revealed that the variation among years was larger in early winter (89.8) than late winter (35.4).
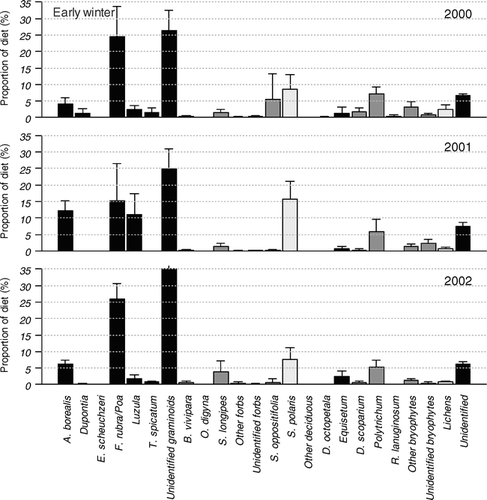
In the recognizable parts of the rumen samples from early winter in 2000 (; in Electronic Appendix), F. rubra/Poa (24.5% ± 9.1%) was the most frequent species group while other graminoids played a minor role. However, the total contribution of graminoids in early winter was less in 2000 (60.2% ± 11.5%) than in 2001 (63.5% ± 9.3%) and 2002 (69.9% ± 4.9%). On the other hand, the diet from this period featured a relatively higher contribution from bryophytes (13.1% ± 4.2%) of different species than early winter in 2001 (9.9% ± 4.5%) and 2002 (7.4% ± 2.4%). In early winter in 2001, A. borealis (12.2% ± 3.1%) and Luzula (11.0% ± 6.4%) together with F. rubra/Poa (15.3% ± 11.2%) contributed to the large percentage of graminoids in the diet. S. polaris (15.8% ± 5.3%) was also found frequently and contributed relatively more than in early winter in 2000 (8.7% ± 4.4%) and 2002 (7.7% ± 3.5%). The samples from early winter in 2002 had the same dominant species/species groups as the samples from early winter in 2000, but in general, graminoids contributed relatively more while bryophytes contributed relatively less.
Figure 2 Percentages of species/species groups ± SD across individuals in the early winter diet in 2000 (n = 10), 2001 (n = 10), and 2002 (n = 6) based on rumen samples from female Svalbard reindeer (Rangifer tarandus platyrhynchus). The different growth forms are displayed using different shadings; graminoids (from left), forbs and evergreen cushion plants, deciduous and evergreen shrubs, sphenophytes, bryophytes, lichens, and unidentified plant fragments.
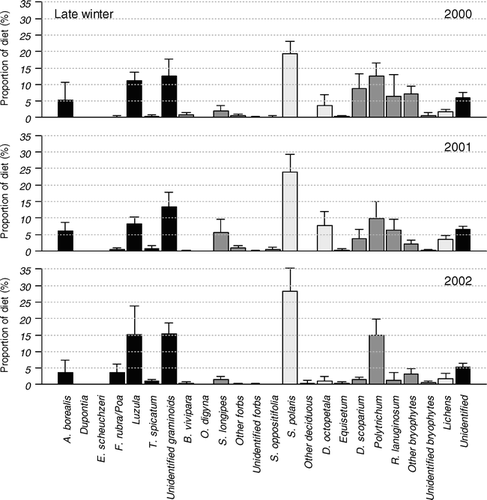
Considering the samples from late winter in 2000 (; in Electronic Appendix), bryophytes (35.7% ± 5.9%) were found most frequently and contributed relatively more than in late winter in 2001 (22.1% ± 8.0%) and 2002 (21.5% ± 3.4%). Graminoids (29.3% ± 9.9%) also represented a large percentage, with Luzula (11.0% ± 2.6%) and A. borealis (5.2% ± 5.4%) as the most frequent species. However, on the species level, S. polaris (19.4% ± 3.7%) contributed the most. The samples from late winter in 2001 and 2002 showed the same overall patterns as the samples from late winter in 2000. However, in 2001 dwarf shrubs were more frequent with more S. polaris (23.8% ± 5.4%) and D. octopetala (7.7% ± 4.2%). In addition, there was a larger percentage from S. longipes (5.5% ± 4.0%) than the other late winters (1.9% ± 1.6% in 2000 and 1.5% ± 0.9% in 2002). In 2002, on the other hand, the percentage from S. polaris (28.3% ± 7.0%), Luzula (15.1% ± 8.7%), and Polytrichum (15.0% ± 4.8%) peaked.
Figure 3 Percentages of species/species groups ± SD across individuals in the late winter diet in 2000 (n = 8), 2001 (n = 8), and 2002 (n = 8) based on rumen samples from female Svalbard reindeer (Rangifer tarandus platyrhynchus). The different growth forms are displayed using different shadings; graminoids (from left), forbs and evergreen cushion plants, deciduous and evergreen shrubs, sphenophytes, bryophytes, lichens, and unidentified plant fragments.
Discussion
Seasonal Variation in The Dietary Composition
This study demonstrates seasonal variations in the dietary composition of Svalbard reindeer during winter. Many forage species that are important during summer, such as various grasses growing in mesic sites and deciduous shrubs growing in a wide range of habitats except for wet marshes ( in Electronic Appendix, but also see CitationPunsvik et al., 1980; CitationVan der Wal et al., 2000a), were also used during early winter. In addition, the early winter diet contained a substantial proportion of bryophytes (10.6% ± 4.5%). Later in the winter, the diet was dominated by species growing in mesic and exposed sites, such as rushes, various dwarf shrubs, evergreens, and bryophytes.
During summer, forage species of high quality and species with large biomass, such as various grasses, sedges, and forbs often growing in wet to mesic sites, were prevalent in the diet of Svalbard reindeer (cf. CitationPunsvik et al., 1980; CitationVan der Wal et al., 2000a). S. polaris was also used during summer (cf. CitationEkern and Kildemo, 1978; CitationPunsvik et al., 1980; CitationVan der Wal et al., 2000a), possibly due to high biomass per area (CitationHalle, 2000) where other preferred forage species occur (Citationvan der Wal et al., 2000a). The presence of Dupontia (7.4% ± 8.7%) and E. scheuchzeri (8.2% ± 10.0%) in the summer diet showed that wetland habitats were used. Drier habitats were also used, indicated by a large contribution of various forbs (19.6% ± 9.4%), such as B. vivipara and O. digyna, which are of high quality during summer (CitationEkern and Kildemo, 1978; CitationStaaland et al., 1983; CitationStaaland, 1984), and various grasses, such as A. borealis, F. rubra subsp. arctica (from now called F. rubra), and Poa species. Based on fecal samples from the same valley system, CitationVan der Wal et al. (2000a) found that the summer diet consisted of more forbs and bryophytes and less graminoids than our results indicated. Although our results are based on observations from only one late summer, differences in methods may have created this bias. For example, rumen samples will give a different picture of the diet than fecal material due to variation in digestibility of plant species (CitationGaare et al., 1977).
In early winter, when plant growth had ceased, it was evident that grasses growing in mesic sites, such as F. rubra and Poa species, were still prevalent in the diet, while wetland species dropped out. In addition, A. borealis, also growing in mesic sites, increased in frequency. Wetland habitats freeze and are inaccessible during winter, most likely leading to a shift in habitat use. Further, CitationStaaland (1984) demonstrated that F. rubra and Poa have lower concentrations of crude fiber in autumn than several other graminoid species. There is also a widespread formation of viviparous plantlets in several Poa species, which are found to be of high quality and digestibility (CitationStaaland, 1984). Therefore, the continuous use of F. rubra and Poa in early winter as shown in our study may be a function of both availability and relatively high quality. S. polaris was prevalent in the diet in all seasons, which is in line with other studies of Svalbard reindeer (cf. CitationEkern and Kildemo, 1978; CitationPunsvik et al., 1980; CitationSørmo et al., 1999; CitationBjune, 2000; CitationVan der Wal et al., 2000a), but this species made an increasing contribution to the diet from summer until late winter (). The use of S. polaris in early winter may either be due to its high quality (i.e. it is actively selected), or due to the occurrence of the species together with other preferred forage species in mesic habitats, as was suggested by CitationVan der Wal et al. (2000a) for summer conditions.
While the contribution of shrubs increased during winter, the frequency of forbs decreased from late summer to winter (). During summer, many forbs are favorable forage plants, but they wither at the end of the growing season and are usually confined to sheltered habitats. Thus they are already less accessible by early winter, when a thin snow cover often has developed. However, the forb Stellaria longipes, which is still green in early winter, and the evergreen cushion plant Saxifraga oppositifolia, both made small contributions to the early winter diet and provided green tissue when other species withered.
In the late winter, the dietary composition indicated another shift in habitat use, as the prevalent species, such as S. polaris, Luzula, and various moss species, are found in exposed habitats, although not exclusively (CitationVan der Wal et al., 2000a). With an increasing contribution of S. polaris, in addition to a relatively high presence of the evergreen shrub Dryas octopetala, it was evident that exposed ridges with S. polaris and D. octopetala became preferred feeding grounds (cf. CitationVan der Wal et al., 2000a; CitationLindner, 2002). As a consequence of such late winter foraging on Salix, CitationBjune (2000) found high amounts of Salix pollen in winter feces from Svalbard reindeer. This may be surprising at first, but because S. polaris plants flower early in spring, their flower buds already contain developed pollen below the thin snow layer that reindeer dig through. The evergreen shrub D. octopetala, on the other hand, may provide green plant tissue during late winter, which in general has higher digestibility than withered, non-evergreen tissue that time of the year (CitationKlein, 1990).
The rush Luzula is an unfavorable forage species during summer (cf. CitationPunsvik et al., 1980; CitationHalle, 2000) due to its low concentration of nitrogen and minerals (CitationEkern and Kildemo, 1978) and low digestibility (CitationStaaland et al., 1983) compared to other forage species. In late winter we found that the frequency of Luzula increased in the reindeer diet, which may be a consequence of decreased accessibility and withering of more palatable species. Aside from the Luzula heaths (CitationVan der Wal et al., 2000a), Luzula spp. are also found on dry exposed ridges where snow melts early (CitationRønning, 1996) while the vegetation in other habitats is covered by hard packed snow (CitationLindner, 2002).
Rangifer usually selects against bryophytes (e.g. CitationFerguson et al., 2001), although heavy exploitation of bryophytes during winter has been reported in several habitats across the northern hemisphere (see review by CitationVan der Wal, 2006). The results of this study indicated that bryophytes contributed substantially to the winter diet, especially to the late winter diet, of Svalbard reindeer. Based on abomasum samples, CitationVan der Wal (2006) reported even larger contribution of bryophytes to the winter diet of the same population of Svalbard reindeer. Bryophytes are in general of poor nutritional quality. Although they may have acceptable levels of some minerals compared to vascular plants during summer, the content of nitrogen is low while the content of lignin (CitationProp and Vulnik, 1992) and fiber is very high (CitationEkern and Kildemo, 1978; CitationStaaland et al., 1983). In an experimental field study on Svalbard reindeer eating goose feces, reindeer showed clear avoidance of moss material in goose droppings (CitationVan der Wal and Loonen, 1998). However, especially during the late arctic winter, the quantity of forage is heavily limited by snow and ice covering the most favorable habitats, and reindeer may be left with no other choice than to eat bryophytes. Actually, Svalbard reindeer may have developed into a specialist moss-consumer (CitationVan der Wal, 2006). They have been shown to have a large digestive system relative to body weight in comparison with other subspecies of R. tarandus (CitationReimers et al., 1982; CitationSørmo et al., 1999). By increasing the volume of the fermentation chambers, rumen turnover time may decrease, resulting in increased digestive capacity (see review by CitationHanley, 1982). Additionally, Svalbard reindeer have very high numbers of fiber-digesting rumen bacteria (CitationOrpin et al., 1985), easing the processing of poorly digested forage such as bryophytes.
In the High Arctic areas where preferred lichens are scarce, it has been suggested that bryophytes have replaced lichens as important winter forage in reindeer (CitationStaaland et al., 1983; CitationStaaland, 1986; CitationLongton, 1997). The only lichen genus identified in our study was Stereocaulon. In a study of reindeer's effects on high arctic lichens, Stereocaulon spp. seemed to be least affected by grazing (CitationVan der Wal et al., 2001), possibly resulting in relatively high availability of this genus compared to other lichen genera.
Interannual Variation in The Winter Diet
Our study demonstrates that there is an interannual variation in the dietary composition of Svalbard reindeer. Data from three years are insufficient to draw any conclusions about the causality of this matter, but it provides a basis for suggesting several hypotheses that will need further testing. Two hypotheses can be used to account for the variation among years in early winter, one associated with the amount of snow present at the time, and the other associated with the amount of rain the previous summer. Meteorological data are from the airport in Longyearbyen, situated 15–30 km from the study area (Norwegian Meteorological Institute). (1) In the early winter, the amount of precipitation as snow can potentially influence habitat use, although forage is normally available across all vegetation types this time of the year. In October 2001 there were higher amounts of precipitation (14.7 mm) on days with temperatures <0°C than in October 2000 (4.5 mm) and 2002 (4.2 mm). Based on our results we therefore hypothesize that the amount of snow affects the dietary composition in early winter through variation in plant availability. Thus, when a relatively high amount of snow limits plant availability, the diet of Svalbard reindeer may consist primarily of species with an erect growth form in mesic sites, such as A. borealis, F. rubra, and Poa species, or rushes and dwarf shrubs growing in exposed habitats, such as Luzula and S. polaris. In contrast, low amounts of snow may allow relatively more consumption of forbs and bryophytes, which have much of their biomass located close to the ground. (2) Conditions for plant growth during summer also influence food availability during winter. For instance, cold and wet summers limit plant growth, which in turn negatively influence population growth rates for Svalbard reindeer, suggesting that there may be a bottom-up effect in the ecosystem (CitationAanes et al., 2002). Within years, variation in biomass availability may influence the selection of forage (CitationAlbon and Langvatn, 1992; CitationLangvatn and Hanley, 1993; CitationVan der Wal et al., 2000b). Based on our results, we hypothesize that among-year variation in biomass availability, caused by variations in summer precipitation, initiates among-year variation in dietary composition during early winter. During the summer of 2000, there was slightly more rain (43.5 mm during June, July, and August) than in 2001 (40.6 mm) and 2002 (40.6 mm). The relatively large consumption of bryophytes by reindeer in the early winter of 2000 might be explained by lower availability of high quality vascular plant forage at that time compared with the other two years.
During late winter, snow and ice are thought to restrict the number of available forage sites (CitationReimers, 1982), hence the options for selection at the species level may be limited. In agreement with this, we found that the interannual variation in dietary composition was smaller during late winter than during early winter. The amount of precipitation during days with temperatures >0°C during winter, known as the “icing index” (CitationSolberg et al., 2001), has previously been used to describe the severity of the icing condition at Svalbard. The index has been found to be positively correlated with total winter precipitation (CitationAanes et al., 2000). During the three late winters in our study, bryophytes contributed to the Svalbard reindeer diet to a larger extent when the icing index was small, as in 2000 (icing index = 7.2). One may hypothesize that when the winter conditions are relatively favorable, reindeer may be able to reach biomass located close to the ground, e.g. by cratering. In 2001 and 2002, when the icing indices were larger (41.9 and 29.4, respectively), indicating less favorable winter conditions, the diet featured a higher percentage of dwarf shrubs, such as S. polaris and D. octopetala, growing on exposed ridges where snow melts early.
Conclusions
It is evident that plant availability is crucial for the diet of Svalbard reindeer during winter. However, the early winter diet seemed to be an intermediate between the summer diet, when species of high quality and species with large biomass are used, and the late winter diet, which is characterized by the presence of species growing in exposed habitats. We found that plant species growing in both mesic heaths and on dry, exposed ridges, such as rushes, dwarf shrubs, evergreen plants, and various bryophyte species, became increasingly important during the winter, suggesting the importance of plant biomass availability. Previously, the growth rate of Svalbard reindeer populations has been related to summer and winter climate (CitationReimers, 1982; CitationAanes et al., 2000; CitationSolberg et al., 2001). We hypothesize that such climatic effects may be mediated through interannual variation in the diet of the Svalbard reindeer where increased summer precipitation and winter rain may affect dietary composition through decreased forage biomass availability. However, in order to make robust conclusions on the link between climate and the Svalbard reindeer diet, long-term studies of dietary composition are needed, incorporating the effects of climate on local plant growth and plant availability.
uaar_a_11957378_sm0001.pdf
Download PDF (60.5 KB)uaar_a_11957378_sm0004.pdf
Download PDF (60.3 KB)uaar_a_11957378_sm0003.pdf
Download PDF (56.8 KB)uaar_a_11957378_sm0002.pdf
Download PDF (60.3 KB)uaar_a_11957378_sm0006.pdf
Download PDF (60.3 KB)uaar_a_11957378_sm0005.pdf
Download PDF (56.8 KB)Acknowledgments
We would like to thank the Norwegian University of Science and Technology (NTNU) and The University Centre in Svalbard (UNIS) for financial support. We are also grateful to Justin Irvine and collaborators for providing rumen samples, and to Christiane Hübner, Vebjørn Veiberg, and Leif Egil Loe for help and advice on Svalbard issues. Thanks to Claudia Melis for help with R, to Snorre Henriksen, Brage Bremset Hansen, Marianne Havnes, and Geir H. Bolstad for fruitful discussions on topics related to the present paper, to Nancy Bazilchuk for linguistic revision, and to Rene Van der Wal and an anonymous referee for valuable comments on the manuscript.
Related Research Data
References Cited
- Aanes, R. , B-E. Sæther , and N. A. Øritsland . 2000. Fluctuations of an introduced population of Svalbard reindeer: the effects of density dependence and climatic variation. Ecography 23:437–443.
- Aanes, R. , B-E. Sæther , F. M. Smith , E. J. Cooper , P. A. Wookey , and N. A. Øritsland . 2002. The Arctic Oscillation predicts effects of climate change in two trophic levels in a High Arctic ecosystem. Ecology Letters 5:445–453.
- Aanes, R. , B-E. Sæther , E. J. Solberg , S. Aanes , O. Strand , and N. A. Øritsland . 2003. Synchrony in Svalbard reindeer population dynamics. Canadian Journal of Zoology 81:103–110.
- Albon, S. D. and R. Langvatn . 1992. Plant phenology and the benefits of migration in a temperate ungulate. Oikos 65:502–513.
- Albon, S. D. , A. Stien , J. R. Irvine , R. Langvatn , E. Ropstad , and O. Halvorsen . 2002. The role of parasites in the dynamics of a reindeer population. Proceedings of the Royal Society B269:1625–1632.
- Anderson, M. J. 2001. A new method for non-parametric multivariate analysis of variance. Austral Ecology 26:32–46.
- Anderson, M. J. 2003. XMATRIX: calculation of design matrices for ANOVA in linear models, a computer program. Department of Statistics, University of Auckland, New Zealand.
- Anderson, M. J. 2004. DISTLM v.5: distance-based multivariate analysis for a linear model, a computer program. Department of Statistics, University of Auckland, New Zealand.
- Anderson, M. J. and C. J F. ter Braak . 2003. Permutation tests for multi-factorial analysis of variance. Journal of Statistical Computation and Simulation 73:85–113.
- Bjune, A. E. 2000. Pollen analysis of faeces as a method of demonstrating seasonal variations in the diet of Svalbard reindeer ( Rangifer tarandus platyrhynchus ). Polar Research 19:183–192.
- Brattbakk, I. Øritsland, N. A. 1986. Flora og vegetasjon. In . Svalbardreinen og dens livsgrunnlag. Oslo Universitetsforlaget AS. 15–34.
- Chapin III, F. S. , K. Van Cleve , and L. L. Thiezen . 1975. Seasonal nutrient dynamics of tundra vegetation at Barrow, Alaska. Arctic and Alpine Research 7:209–226.
- Crawford, R. M M. 1989. Studies in Plant Survival: Ecological Case Histories of Plant Adaptation to Adversity. Oxford Blackwell Scientific Publications.
- Ekern, K. and K. Kildemo . 1978. Svalbardreinens ernæring. Hovedoppgave, Institutt for Naturforvaltning. Norges Landbrukshøyskole, Norway.
- Elvebakk, A. Wielgolaski, F. E. 1997. Tundra diversity and ecological characteristics of Svalbard. In . Ecosystems of the World, Polar and Alpine Tundra, 3 . Amsterdam Elsevier Scientific. 347–359.
- Elvebakk, A. and P. Prestrud . 1996. A Catalogue of Svalbard Plants, Fungi, Algae and Cyanobacteria. Skrifter 198. Oslo Norwegian Polar Institute.
- Ferguson, M. A D. , L. Gauthier , and F. Meisser . 2001. Range shift and winter foraging ecology of a population of Arctic tundra caribou. Canadian Journal of Zoology 79:746–758.
- Gaare, E. , A. Sørensen , and R. G. White . 1977. Are rumen samples representative of the diet. Oikos 29:390–395.
- Golley, F. B. and H. C. Buechner . 1968. A Practical Guide to the Study of the Productivity of Large Herbivores. IBP Handbook, 7 . Oxford-Edinburgh Blackwell.
- Halle, Y. 2000. Forage selection of vascular plants in different spatial and temporal scales by Svalbard reindeer ( Rangifer tarandus platyrhynchus Vrolik). Candidata scientiarum thesis. University of Oslo, Norway.
- Hanley, T. A. 1982. The nutritional basis for food selection by ungulates. Journal of Range Management 35:146–151.
- Henriksen, S. , R. Aanes , B-E. Sæther , T. H. Ringsby , and J. Tufto . 2003. Does availability of resources influence grazing strategies in female Svalbard reindeer. Rangifer 23:25–37.
- Jefferies, R. L. , D. R. Klein , and G. R. Shaver . 1994. Vertebrate herbivores and northern plant communities: reciprocal influences and responses. Oikos 71:193–206.
- Jónsdóttir, I. S. 2005. Terrestrial ecosystems on Svalbard: heterogeneity, complexity and fragility from an arctic island perspective. Biology and Environment: Proceedings of the Royal Irish Academy 105B:155–165.
- Klein, D. R. 1990. Variation in quality of caribou and reindeer forage plants associated with season, plant part, and phenology. Rangifer Special Issue 3:123–130.
- Kudo, G. 1991. Effects of snow-free period on the phenology of alpine plants inhabiting snow patches. Arctic and Alpine Research 23:436–443.
- Langvatn, R. and T. Hanley . 1993. Feeding-patch choice by red deer in relation to foraging efficiency. An experiment. Oecologia 95:164–170.
- Legendre, P. and M. J. Anderson . 1999. Distance-based redundancy analysis: testing multispecies responses in multifactorial ecological experiments. Ecological Monographs 69:1–24.
- Lindner, E. 2002. Use of vegetation types by Svalbard reindeer from arctic winter to spring. Polar Record 39:245–247.
- Longton, R. E. Woodin, S. 1997. The role of bryophytes and lichens in polar ecosystems. In . Ecology of Arctic Environments 69–96. British Ecological Society Symposium Volume. Oxford: Blackwell.
- Mysterud, A. , R. Langvatn , N. G. Yoccoz , and N. C. Stenseth . 2001. Plant phenology, migration and geographical variation in body weight of a large herbivore: the effect of a variable topography. Journal of Animal Ecology 70:915–923.
- Øritsland, N. A. and E. Alendal . Øritsland, N. A. 1986. Bestandens størrelse og livshistorie. In . Svalbardreinen og dens livsgrunnlag. Oslo Universitetsforlaget AS. 52–60.
- Orpin, C. G. , S. D. Mathiesen , Y. Greenwood , and A. S. Blix . 1985. Seasonal changes in the ruminal microflora of the high-arctic Svalbard reindeer ( Rangifer tarandus platyrhynchus ). Applied and Environmental Microbiology 50:144–151.
- Prop, J. and T. Vulnik . 1992. Digestion by barnacle geese in the annual cycle: the interplay between retention time and food quality. Functional Ecology 6:180–189.
- Punsvik, T. , A. Syvertsen , and H. Staaland . Reimers, E. , E. Gaare , and S. Skjenneberg . 1980. Reindeer grazing in Adventdalen, Svalbard. In . Reindeer/Caribou Symposium II. Norway 1979 . Trondheim Direktoratet for Vilt og Ferskvannsfisk. 115–123.
- Reimers, E. 1982. Winter mortality and population trends of reindeer on Svalbard, Norway. Arctic and Alpine Research 14:295–300.
- Reimers, E. , T. Ringberg , and R. Sørumsgård . 1982. Body composition of Svalbard reindeer. Canadian Journal of Zoology 60:1812–1821.
- Rønning, O. I. 1996. Svalbards flora. Polarhåndbok 9 . Oslo Norwegian Polar Institute.
- Solberg, E. J. , P. Jordhøy , O. Strand , R. Aanes , A. Loison , B-E. Sæther , and J. D C. Linnell . 2001. Effects of density-dependence and climate on the dynamics of a Svalbard reindeer population. Ecography 24:441–451.
- Sørmo, W. , ØE. Haga , E. Gaare , R. Langvatn , and S. D. Mathiesen . 1999. Forage chemistry and fermentation chambers in Svalbard reindeer ( Rangifer tarandus platyrhynchus ). Journal of Zoology 247:247–256.
- Staaland, H. 1984. On the quality of Svalbard reindeer pasture in the summer and autumn. Rangifer 4:16–23.
- Staaland, H. Øritsland, N. A. 1986. Svalbardreinens ernæring. In . Svalbardreinen og dens Livsgrunnlag. Oslo Universitetsforlaget AS. 72–91.
- Staaland, H. and T. Punsvik . Reimers, E. , E. Gaare , and S. Skjenneberg . 1980. Reindeer grazing on Nordaustlandet, Svalbard. In . Reindeer/Caribou Symposium II. Norway 1979 . Trondheim Direktoratet for Vilt og Ferskvannsfisk. 142–150.
- Staaland, H. and K. Røed . Øritsland, N. A. 1986. Om Svalbardreinens slektsskapsforhold og opprinnelse. In . Svalbardreinen og dens Livsgrunnlag. Oslo Universitetsforlaget AS. 61–71.
- Staaland, H. , I. Brattbakk , K. Ekern , and K. K. Kildemo . 1983. Chemical composition of reindeer forage in Svalbard and Norway. Holarctic Ecology 6:109–122.
- Staaland, H. , J. O. Scheie , F. A. Grøndahl , E. Persen , A. B. Leifseth , and O. Holand . 1993. The introduction of reindeer to Brøggerhalvøya, Svalbard: grazing preference and effect on vegetation. Rangifer 13:15–19.
- Storeheier, P. V. , S. D. Mathiesen , N. J C. Tyler , and M. A. Olsen . 2002. Nutritive value of terricolous lichens for reindeer in winter. Lichenologist 34:247–257.
- Tyler, N. J C. 1986. The relationship between the fat content of Svalbard reindeer in autumn and their death from starvation in winter. Rangifer Special Issue 1:311–314.
- Tyler, N. J C. 1987. Body composition and energy balance of pregnant and non-pregnant Svalbard reindeer during winter. Symposia of the Zoological Society of London 57:203–229.
- Tyler, N. J C. and N. A. Øritsland . 1989. Why don't Svalbard reindeer migrate. Holarctic Ecology 12:369–376.
- Underwood, A. J. 1997. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance. Cambridge Cambridge University Press.
- Van der Wal, R. 2006. Do herbivores cause habitat degradation or vegetation state transition? Evidence from the tundra. Oikos 114:177–186.
- Van der Wal, R. and M. J J. E. Loonen . 1998. Goose droppings as food for reindeer. Canadian Journal of Zoology 76:1117–1122.
- Van der Wal, R. , J. Irvine , A. Stien , N. Shepherd , and S. D. Albon . 2000a. Faecal avoidance and the risk of infection by nematodes in a natural population of reindeer. Oecologia 124:19–25.
- Van der Wal, R. , N. Madan , S. M J. van Lieshout , C. Dormann , R. Langvatn , and S. D. Albon . 2000b. Trading forage quality for quantity? Plant phenology and patch choice by Svalbard reindeer. Oecologia 123:108–115.
- Van der Wal, R. , R. Brooker , E. Cooper , and R. Langvatn . 2001. Differential effects of reindeer on High Arctic lichens. Journal of Vegetation Science 12:705–710.
- Walker, D. A. , W. A. Gould , H. A. Maier , and M. K. Raynolds . 2002. The circumpolar Arctic vegetation map: AVHRR-derived base maps, environmental controls, and integrated mapping procedures. International Journal of Remote Sensing 23:4551–4570.
- White, R. G. 1983. Foraging patterns and their multiplier effects on productivity of northern ungulates. Oikos 40:377–384.