
Mention the words “Gibson cloning” and many synthetic biologists light up. The Gibson isothermal assembly technique (Citation1), which involves multiple DNA fragments overlapping at their ends being simultaneously joined by an exonuclease, a DNA polymerase, and a DNA ligase, has been a boon to the field.
In the Gibson method, the exonuclease chews back the ends of each fragment, leaving 3′ single-stranded overhangs. Fragments anneal to each other through their complementary overhangs, with the gaps filled in by DNA polymerase and the nicks sealed by DNA ligase.
Although Gibson cloning has made a name for itself with synthetic biologists around the globe for its ability to join together multiple fragments to form large natural genes or even small bacterial genomes, the technique has not been used to assemble numerous transcriptional units (TUs) in a modular fashion for artificial genetic circuits, a key synthetic biology application. But now, two papers (Citation2,Citation3) have been published describing similar methods for the rapid, modular construction of large, complex gene circuits that demonstrate the scalability of the Gibson technique, launching a new wave of Gibson fans.
Not UNSpecific
“It became pretty clear that something like Gibson technology could be one of those disruptive technologies that really changes how we approach synthetic biology,” explained Ron Weiss, associate professor in the Department of Biological Engineering at MIT and senior author of the paper (Citation2) published in July in the journal Nucleic Acids Research. However, people who used the Gibson method “realized that it’s not trivial to take parts and then create circuits out of them” and that the results depended on the actual sequence of the parts, with some working well and others not.
Synthetic biologists need to use standardized, well-characterized elements such as promoters, ribosome binding sites, and terminators for the TUs in a genetic circuit, which presented another critical consideration for Joseph Torella, a graduate student in Pamela Silver’s lab in the Department of Systems Biology at the Harvard Medical School and lead author of the other Gibson assembly article that recently appeared in Nucleic Acids Research (Citation3). “The challenge was that if you tried to Gibson together several parts where they have promoters and terminators on each end and they all kind of look alike, you can’t do it because you need unique homology regions to facilitate the assembly.”
To solve these problems, both groups placed algorithmically generated unique nucleotide sequences (UNSs) at the ends of the component TUs to be Gibson-assembled into a single construct. Systematic use of these UNSs allowed components containing repeated sequences such as standardized promoters or terminators to be joined in an ordered fashion and ensured consistency in the efficiency of end joining during Gibson assembly.

Rapid and modular
Excited by the apparent scalability of Gibson cloning but still cognizant of the difficulties of designing overhangs for each set of sequences involved, Weiss sought “to create a platform that would really simplify the whole construction process…make it as predictable as possible…for even people with limited experience to be able to build very large circuits.”
In the Weiss lab’s approach, which was developed for mammalian gene circuits, 40 bp UNSs were each computationally designed to maximize self-annealing in the Gibson reaction but to minimize misannealing to other sequences and hairpin formation.
The authors first constructed each TU by selecting a promoter and a gene from a library of standard sequence-verified parts and then cloning the pair using Gateway recombination into an appropriate destination vector called a “position” vector. Each position vector contains a chromatin insulator sequence, a Gateway recombination cassette, and a polyadenylation sequence, which are all flanked by two different 40 bp UNSs that are flanked in turn by two I-SceI restriction sites. (I-SceI is an extremely rare base cutter whose sequence is not found in the human or mouse genomes.)
I-SceI digestion of each position vector releases a fragment containing one TU with two different UNwSs at its ends. Ordered assembly of these TU fragments is then achieved in a Gibson reaction where the UNS 1/2 fragment is joined to the UNS 2/3 fragment, which is then joined to the UNS 3/4 fragment, and so on, until the final UNS (N-1)/N fragment is joined using an adapter vector containing UNS N/X to the carrier vector containing UNS 1/X and E. coli propagation sequences.
Using their method, Weiss’s group first demonstrated they could assemble as many as 11 TUs into a single vector. They next cloned a 7-TU gene circuit into mammalian cells and observed strong ligand-inducible expression from the circuit. Finally, they used the technique to hierarchically assemble a 45 kb 7-TU module with 5 additional TUs to create a 63 kb 12-TU circuit. The additional TUs contained components already present in the 7-TU circuit, demonstrating that the assembly method was not affected by repetitive sequences in the assembled parts.
Modular and rapid
For Torella, the impetus for improving Gibson assembly was his interest in optimizing fatty acid biosynthesis in bacteria. To accomplish this, he needed to co-express various proteins as well as build the necessary gene circuits.
As with Weiss’s approach, Torella’s method depends on 40-nucleotide UNSs placed at the ends of the genetic circuit components for their ordered and simultaneous Gibson assembly. In this case, 100,000 random 40-mer sequences were algorithmically screened to eliminate sequences that contained restriction sites or start codons, were GC-rich, formed hairpins, had high BLAST scores for the E. coli genome, and finally those that were likely to misassemble with each other. This screen left ∼50 UNSs, of which 10 are described in the paper.
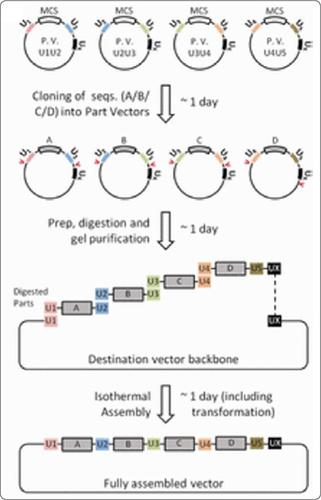
These UNSs were incorporated into “part” and “destination” vectors used for Gibson assembly of TUs into genetic circuits. Each part vector consists of a UNS “UN”, a multiple cloning site (MCS; with BioBrick and BglBrick sites), another UNS “UN+1”, and finally a third UNS “UX” that is common to all the part vectors. A promoter, a gene, and a terminator were cloned into the MCS of a part vector to create each TU of the final genetic circuit to be assembled. Prior to Gibson assembly, each TU construct was digested with restriction enzymes flanking UN and UN+1, except for the final piece, which was instead digested at restriction sites flanking UN+1 and UX. The gel-isolated, UNS-flanked TU fragments were then Gibson-assembled with a digested PCR-purified destination vector containing U1 and UX, with the order of the TUs determined by the UNSs flanking them.
It was clear that the presence of repeated sequences in the pieces had no effect on the success of assembly since they were able to efficiently assemble 5 pieces, each consisting only of a T7 promoter, MCS, and T7 terminator and which were 80% similar to each other, into a destination vector. The authors also demonstrated that strongly insulated parts could be assembled into expression libraries where the expression level of each TU was determined by its own promoter’s strength without affecting expression levels of other TUs.
The authors used the method to construct and optimize a bacterial biosynthetic pathway that produces the chemical deoxychromoviridans using three genes from Chromobacterium violaceum. Each gene was combined with 6 possible promoters and 1 terminator sequence, and then all 216 combinations of the 3 genes were assembled and transformed into E. coli.
Testing of insert sizes and sequencing of randomly selected clones showed very efficient assembly, and these clones produced varying levels of deoxychromoviridans. Interestingly, the high-producing strains all had low expression of one gene and moderate expression of the two other genes, indicating that engineering strong expression of all three genes was not the optimal design strategy for this 3-TU biosynthetic pathway circuit, as might have been expected.
Finally, the authors used the technique to construct mammalian circuits by assembling two- and three-input split TALE AND gates into a single BAC for integration into mouse embryonic stem cells (ESC). Ironically enough, this wasn’t an original goal of Torella’s, and the results (though not the methodology) of the ESC work were actually published before his paper describing the technique. After his preliminary success using the method for assembling bacterial circuits, other groups in the Silver lab began using it as well for their projects.
“The mammalian cell people got so excited about it that they beat me to the punch and published their embryonic stem cell circuit work before I even published the methodology,” noted Torella with much amusement. Due to another paper on his fatty acid biosynthesis work that had some very demanding reviewers, “I got sidetracked for probably three solid months where I wasn’t working on this assembly paper because of that,” explained Torella.
And in a further ironic twist that illustrates the fickle nature of scientific success, Torella’s fatty acid biosynthesis project, for which the new assembly method was developed, didn’t work out. “At the end, we had this approach that now everyone in the lab was using because it was clearly valuable and saving people time, but ironically the original application we had in mind hadn’t come through; this is why my friend managed to publish his paper using our method before I got anything out the door.”
Future prospects
Weiss plans to find better insulator sequences, given that the present ones were not as perfect as they hoped, even after multiple cycles of improvement. He also plans to automate the method, for example using liquid handling robots, which he believes really represents an exciting direction for assembly.
In contrast, Torella and the Silver lab are not pursuing automation for their technique, not being convinced that robotics helps to save that much time for their projects based on their and other labs’ experiences. “I think whether you’re working on a robot or at a lab bench as an individual, this kind of approach is going to speed up whatever pathway you’re trying to design,” noted Torella.
Torella believes the major challenge will be trying to understand how to use the methodology in an intelligent way, instead of brute-force empirical testing of all possible combinations of a genetic circuit. “Now that this method exists, we’re no longer held back by the construction of a 5-gene or a 10-gene circuit. …In my mind, the important point in the future probably won’t be improving the assembly methodology so much as it will be improving our understanding of the systems we’re trying to put together adequately so we don’t have to assemble 55 constructs (for example) every time we want to build a biosynthetic pathway.”
Torella’s method currently is being used in the Silver lab for building optimized alkane biosynthetic pathways in E. coli, building logic gates in stem cells for therapeutic applications, and for Torella’s current project of building a “bionic leaf.” The latter is a collaboration with Dan Nocera’s lab in the Dept. of Chemistry at Harvard, where they “are interfacing autotrophic bacteria with a really efficient solar water-splitting catalyst to try to make a hybrid inorganic microbial photosynthetic system.” With that system mostly working, they’re now using the multi-gene assembly method to engineer a new bacterium, Ralstonia eutropha, to produce biofuel and sugar within the system. Such a fascinating and potentially high-impact synthetic biological application clearly illustrates the potential of the method developed by the Weiss and Silver labs.
References
- Gibson, D.G., L.Young, R.Y.Chuang, J.C.Venter, C.A.Hutchison, and H.O.Smith. 2009. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods6:U343–U341.
- Guye, P., Y.Li, L.Wroblewska, X.Duportet, and R.Weiss. 2013. Rapid, modular and reliable construction of complex mammalian gene circuits. Nucleic Acids Res.41:e156.
- Torella, J.P., C.R.Boehm, F.Lienert, J.H.Chen, J.C.Way, and P.A.Silver. 2013. Rapid construction of insulated genetic circuits via synthetic sequence-guided isothermal assembly. Nucleic Acids Res. 2013 Sep 26. [Epub ahead of print].

