
Single strand conformational polymorphism (SSCP) is a mobility shift analysis of single-stranded DNA (ssDNA) by nondenaturing polyacrylamide gel electrophoresis to detect DNA sequence polymorphisms (Citation1). Typical SSCP protocols detect single strands by autoradiography (Citation2–4) or by treatment with a DNA stain after electrophoresis (Citation5–12). We report here that treatment with a nucleic acid stain before electrophoresis reveals polymorphisms undetected by staining after electrophoresis.
Our SSCP protocol includes four phases. In phase I, double-stranded DNA (dsDNA) is denatured into ssDNA by heating for 5 min at 95°C in loading buffer (95% formamide, 20 mM NaOH, 0.05% xylene cyanol, 0.05% bromphenol blue, 10 mM EDTA). Phase II stabilizes ssDNA by rapid cooling on ice. In phase III, ssDNA fragments are separated by electrophoresis in a nondenaturing gel. Phase IV is staining (if necessary) and imaging.
SYBR® Gold is a proprietary, cyanine-based nucleic acid stain from Molecular Probes (Eugene, OR, USA) and is characterized by Tuma et al. (Citation13). We recently adopted it for detecting DNA in gels. It is more sensitive than ethidium bromide and more time efficient than silver staining. To achieve additional efficiency, we include SYBR Gold in the phase I buffer at the manufacturer's suggested stain bath concentration of 1× = 1/10,000 dilution of stock stain. This pre-electrophoretic (pre-staining) eliminates a phase IV staining step (post-staining), decreasing by 1–4 orders of magnitude the amount of SYBR Gold required to detect DNA by submerging an entire gel in staining buffer.
Testing of identical samples with pre- and post-staining protocols led to the serendipitous discovery that pre-staining with SYBR Gold reveals SSCPs (, compare lanes 2 and 3 to lanes 4 and 5) undetected by post-staining with SYBR Gold (1× stain bath; , lanes 12–15) or ethidium bromide (0.5 µg/mL stain bath; , lanes 17–20). Pre-staining with ethidium bromide (0.5 µg/mL phase I buffer) failed to detect these same polymorphisms (, lanes 7–10).
SSCP of H09g marker for two Tribolium castaneum strains (M2, mas p au; T2, Tiw-1) show duplicate applications in each of four treatments (EB, ethidium bromide; SYB, SYBR Gold). λ DNA (50 ng) loaded into lanes marked “L” served as fluorescent controls. This 10% polyacrylamide gel [99:1 methylene-bis-acrylamide in 1× Tris-borate EDTA (TBE)] was run in a vertical position at 10°C (5°C water jacket) with 0.5× TBE running buffer for 60 min at 180 constant volts. Pre- and post-stained samples were run on the same gel and physically separated for postelectrophoretic treatment.
![Figure 1. Four single strand conformational polymorphism (SSCP) staining methods in a single polyacrylamide gel. SSCP of H09g marker for two Tribolium castaneum strains (M2, mas p au; T2, Tiw-1) show duplicate applications in each of four treatments (EB, ethidium bromide; SYB, SYBR Gold). λ DNA (50 ng) loaded into lanes marked “L” served as fluorescent controls. This 10% polyacrylamide gel [99:1 methylene-bis-acrylamide in 1× Tris-borate EDTA (TBE)] was run in a vertical position at 10°C (5°C water jacket) with 0.5× TBE running buffer for 60 min at 180 constant volts. Pre- and post-stained samples were run on the same gel and physically separated for postelectrophoretic treatment.](/cms/asset/de47f89d-2974-438d-b69b-09a4a3a85c21/ibtn_a_12357935_f0001.gif)
We screened PCR products of nine sequence-tagged sites (STS) amplified from two distantly related strains (mas p au, Tiw-1) of the red flour beetle, Tribolium castaneum. The post-staining protocol using ethidium bromide detected three polymorphisms (, Post). The pre-staining protocol using SYBR Gold detected the same three polymorphisms (, Pre) plus three additional polymorphisms (, Pre). In no case did post-staining reveal a polymorphism that was not revealed by pre-staining. In some cases, neither protocol detected a polymorphism (). Sequence analysis corroborated the absence of sequence polymorphism in these latter cases and the presence of sequence polymorphism in all other cases (). One case found resolution of the polymorphism to be sensitive to MgCl2 concentration in the phase I buffer. Pre-stain loading buffers effectively stained DNA up to 14 days if held at 5°–6°C. In addition we have successfully reproduced SSCP patterns from PCR-amplified DNA stored for 18 months at 5°–6°C in the phase I buffer containing SYBR Gold (data not shown).
Negative fluorescent images are shown for two strains of Tribolium castaneum (M, mas p au; T, Tiw-1), two staining methods (SYBR Gold pre-stain, ethidium bromide post-stain), and nine molecular markers. Markers are grouped as: (A) stain-dependent polymorphisms (H09g, H115, M2s); (B) stain-independent polymorphisms (A011, B16s, F2s); and (C) nonpolymorphic (Tsh, J1s, and CHSA). Polyacrylamide gel electrophorsis conditions were same as and run for 90 min. Pre- and post-stained samples were run on the same gel and physically separated for postelectrophoretic treatment. STS, sequence-tagged site.
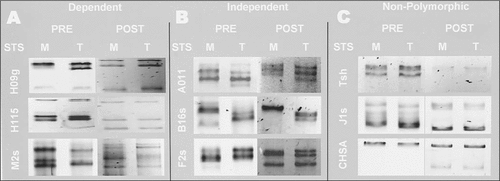
Table 1. STS Polymorphisms
We surmise that SYBR Gold interacts with ssDNA in a sequence-specific way to reveal SSCPs. Cyanine-based nucleic acid stains have been shown to alter the molecular character of DNA both electrostatically and structurally and as a result modify electrophoretic properties (Citation14). Our results further support this premise with the influence upon mobility shifts and show sensitivity to a single nucleotide polymorphism within an ssDNA fragment of 138 nucleotides, and to larger fragments (401 nucleotides) with multiple polymorphisms. Pre-stained PCR products show SSCP pattern reproducibility within gels and across gels over time. Investigation is under way to determine sequence characteristics of the pre-stain dependency.
Acknowledgments
Richard W. Beeman (U.S. Department of Agriculture, Grain Marketing Research and Production Laboratory, Manhattan, KS) provided the Tribolium strains and some STS sequences. Ben Krenke (University of Wisconsin-Parkside) also provided STS sequence. Technical assistance was provided by Maribel Cabral, Ahmed Cheema, and Crystal Smith. Sequence analysis was provided by Charles Nicolet (University of Wisconsin, Madison). Assistance with images was provided by Donald Lintner (University of Wisconsin-Parkside).
References
- Orita, M., H.Iwahana, H.Kanazawa, K.Hayashi, and T.Sekiya. 1989. Detection of polymorphisms of human DNA by gel electrophoresis as single-strand conformation polymorphisms. Proc. Natl. Acad. Sci. USA86:2766–2770.
- Saeki, Y., S.Ueno, N.Takahashi, F.Soga, and T.Yanagihara. 1992. A novel mutant (transthyretin Ile-50) related to amyloid polyneuropathy. Single-strand conformation polymorphism as a new genetic marker. FEBS Lett.308:35–37.
- Erster, S., N.Slade, and U.M.Moll. 2003. Mutational analysis of p53 in human tumors: direct DNA sequencing and SSCP. Methods Mol. Biol.234:219–230.
- Gupta, A. and S.Agarwal. 2003. Efficiency and cost effectiveness: PAGE-SSCP versus MDE and Phast gels for the identification of unknown beta thalassaemia mutations. Mol. Pathol.56:237–239.
- Reiss, J., U.Lenz, F.Rininsland, P.Ballhausen, D.Drews, and H.G.Posselt. 1992. A novel CFTR mutation, 4035delA, detected by non-radioactive SSCP analysis. Hum. Genet.90:303–304.
- Ballhausen, W.G. and C.Kraus. 1993. Non-isotopic detection of single-stranded conformation polymorphisms using ethidium bromide/UV light. Appl. Theor. Electrophor.3:129–131.
- Hongyo, T., G.S.Buzard, R.J.Calvert, and C.M.Weghorst. 1993. ‘Cold SSCP’: a simple, rapid and non-radioactive method for optimized single-strand conformation polymorphism analyses. Nucleic Acids Res.21:3637–3642.
- Thunberg, U., M.Alemi, C.Sundstrom, and J.Sallstrom. 1995. Nonradioactive detection of monoclonal immunoglobulin heavy chain gene rearrangement with PCR-SSCP. Hematol. Pathol.9:141–153.
- Kumeda, Y. and T.Asao. 1996. Single-strand conformation polymorphism analysis of PCR-amplified ribosomal DNA internal transcribed spacers to differentiate species of Aspergillus section Flavi. Appl. Environ. Microbiol.62:2947–2952.
- Strippoli, P., S.Sarchielli, R.Santucci, G.P.Bagnara, G.Brandi, and G.Biasco. 2001. Cold single-strand conformation polymorphism analysis: optimization for detection of APC gene mutations in patients with familial adenomatous polyposis. Int. J. Mol. Med.8:567–572.
- Kong, P., C.Hong, P.A.Richardson, and M.E.Gallegly. 2003. Single-strand-conformation polymorphism of ribosomal DNA for rapid species differentiation in genus Phytophthora. Fungal Genet. Biol.39:238–249.
- Schmalenberger, A. and C.C.Tebbe. 2003. Genetic profiling of noncultivated bacteria from the rhizospheres of sugar beet (Beta vulgaris) reveal field and annual variability but no effect of a transgenic herbicide resistance. Can. J. Microbiol.49:1–8.
- Tuma, R.S., M.P.Beaudet, X.Jin, L.J.Jones, C.Y.Cheung, S.Yue, and V.L.Singer. 1999. Characterization of SYBR Gold nucleic acid gel stain: a dye optimized for use with 300-nm ultraviolet transilluminators. Anal. Biochem.268:278–288.
- Carlsson, C., A.Larsson, and M.Jonsson. 1996. Influence of optical probing with YOYO on the electrophoretic behavior of the DNA molecule. Electrophoresis17:642–651.

