
Abstract
Endothelial cell dysfunction plays a critical role in the pathogenesis of cardiovascular diseases. Gene targeted mutant, knockout, or transgenic mice are widely used in the laboratory investigation of these disorders. We describe a simple and reproducible “lumen digestion” technique to isolate aortic endothelial cells from mice that would be useful for researchers in endothelial cell biology. We used wild-type, homozygote, or heterozygote heme oxygenase-1 null mice from which the aorta is isolated and removed under anesthesia. After cauterizing all the branches, both ends of the aorta are cannulated using an Intramedic® PE-20 tube. After flushing the aorta with phosphate-buffered saline (PBS), the lumen is repeatedly instilled (five times) with 50 µL 0.25% trypsin in PBS, incubated for 2 min, and flushed with PBS. The outflow is collected in endothelial cell media with 20% fetal bovine serum. After centrifugation, the endothelial cells in the pellet are resuspended in media and plated in a 24-well tissue culture dish. Following culture for 2 to 3 weeks, the cells demonstrate typical cobblestone appearance, stain positive for the endothelial marker CD31, and are capable of low-density lipoprotein uptake. Following challenge with oxidized lipids, heme oxygenase-1 deficient endothelial cells demonstrate increased susceptibility to cell injury.
Introduction
Vascular endothelium is no longer considered a mere barrier between blood and parenchymal cells. It is also an active, regulatory organ that participates in many physiological and patho-physiological processes. The functions of the endothelium include endothelial barrier function, regulation of micro-vascular permeability and vascular tone, angiogenesis, and metabolism. Endothelial dysfunction occurs in pathological conditions involving inflammation, tumor onset and progression, transplant rejection, and wound healing and contributes to the pathogenesis of diseases such as atherosclerosis, hypertension, ischemic heart disease, diabetic complications, sepsis, vasculitis, and thromboembolic diseases.
The availability of transgenic mice with either a deficient or an overexpressed gene has provided investigators the ability to study the role of genetic modifiers in the pathogenesis of these diseases. While intact animal studies and isolated vessel preparations have provided significant advances in vascular biology, the use of in vitro endothelial cell cultures allows for more precise and specific information. Several endothelial isolation procedures have been previously reported (Citation1–8). Many of these result in the generation of mixed population of cells, require digestion with collagenase, and use of matrigels, fluorescent-activated cell or magnetic bead sorting. We report a new “lumen digestion” technique, for isolation of pure aortic endothelial cells from heme oxygenase-1 (HO-1) deficient mice that can serve as a model system for developing in vitro endothelial cell cultures from transgenic mice.
Materials and methods
Animals
Homozygous (−/−), heterozygous (+/−), and wild-type (+/+) HO-1 mice (C57Bl/6 × FVB background) from 8 to 10 weeks of age (n = 6–8 from each genotype) were selected from offspring of heterozygous breeding pairs by PCR using tail DNA as previously described (Citation9). The study protocol was approved by the Institutional Animal Care and Use Committee at the University of Florida.
Aortic Endothelial Cell Isolation
Following anesthesia with 2.5% isoflurane, the abdominal fur was trimmed by electronic clips, and the abdomen was sterilized with iodixanol. A midline laporotomy incision was made, and the aorta was isolated and removed carefully, after all branches were cauterized (Bovie® Aaron Medical, St. Petersburg, FL, USA) under an operating microscope. Both ends of the aorta were cannulated with Intramedic® PE-20 tubes (inner diameter, 0.015 inches, outer diameter, 0.053 inches; BD Diagnostic System, Sparks, MD, USA) and tied. The aorta was flushed with 0.5 mL phosphate-buffered saline (PBS), pH 7.4. Then, 50 µL of 0.25% trypsin in PBS was instilled into the aorta and allowed to dwell for 2 min, with this process being repeated five to six times. The digestion fluid of each aorta was drained into a 15-mL tube containing 10 mL of endothelial basal medium (EBM) supplemented with 20% fetal bovine serum (FBS), 50 µg/mL gentamicin, 50 µg/mL amphotericin B, 1 µg/mL hydrocortisone, 10 ng/mL human epidermal growth factor, and 6 µg/mL bovine brain extract (Clonetics, Walkersville, MD, USA). The collected outflow was centrifuged at 300× g for 5 min. The supernatant was discarded, the cell pellet was resuspended in 1 mL complete EBM with 10% FBS and transferred to a 24-well plate, one well per aorta. The cells were grown at 37°C in 95% air and 5% CO2. After 10–14 days, the cells were trypsinized (0.25%) and plated in the same well with complete EBM containing 10% FBS. Cells from two individual mice of the same genotype were pooled in subsequent cultures. The cells were split (1:2) when they reached about 85%–90% confluence.
Identification of the Cells
The cells were identified as endothelial cells by labeling the cells with rat anti-mouse CD31 (platelet endothelial cell adhesion molecule-1; Research Diagnostic, Flanders, NJ, USA) and uptake of 1,1′-dioctadecyl-1,3,3,3′,3′-tetramethyl-indocarbocyanine-acetylated low-density lipoprotein (DiI-Ac-LDL; Biomedical Technologies, Stoughton, MA, USA) (Citation6,Citation10). Staining was also performed using mouse anti-α-smooth muscle actin antibodies (Sigma, St. Louis, MO, USA). Briefly, cells cultured on glass coverslips were fixed in 3.7% formaldehyde and 1% methanol for 10 min. The cells were then permeabilized with fluorescence-activated cell sorting (FACS™) permeabilizing solution (BD Biosciences, San Jose, CA, USA) for 20 min. After incubation with 5% donkey serum for 30 min, the cells were incubated for 2 h with 1:100 dilution of rat anti-mouse CD31 antibody, followed by incubation with 1:400 Cy™3-labeled donkey anti-rat antibody. Between each treatment, the cells were washed with PBS three times for 5 min. The cells were observed under an Olympus Provis fluorescent microscope (Olympus Optical Ltd, Tokyo, Japan) with red filter. For DiI-Ac-LDL-labeling, the cells on a 6-well dish were washed twice with PBS and incubated with 1:20 DiI-Ac-LDL solution in serum-free EBM for 2 h. After washing with PBS three times for 5 min each, the cells were incubated with 10% FBS complete EBM and observed under an immunofluorescence microscope (Provis; Olympus America, Melville, NY, USA).
Immunoblot Analysis
Confluent mouse aortic endothelial cells (MAECs) were treated with PBS (control) or 5 µM hemin (Sigma) for 16 h. Cells were washed twice with ice-cold PBS and lysed in a buffer containing a broad spectrum mixture of protease inhibitors consisting of 10 µg/mL aprotinin, 5 mM EDTA, 1 µg/mL leupeptin, 0.7 µg/mL pepstatin A, 1 mM phenylmethanesulfonyl fluoride, and Triton® X-100. Protein concentration of lysates was assessed by the bicinchoninic acid (BCA) assay (Pierce, Rockford, IL, USA). Samples were separated in a 12% sodium dodecyl sulfate (SDS) polyacrylamide gel and then transferred onto a polyvinylidene difluoride membrane. The membranes were incubated for 1.5 h with the anti-HO-1 and/or anti-HO-2 antibody (1:1000 dilution) (Stressgen, Victoria, BC, Canada) followed by incubation with peroxidase-conjugated goat anti-rabbit immunoglobulin G (IgG) antibody (1:20,000 dilution) for 1 h. Labeled protein bands were examined by using an enhanced chemiluminescence method (ECL™ Amersham Biosciences, Piscataway, NJ, USA) according to the manufacturer's recommendation.
DNA Isolation and PCR Analysis
MAECs were collected by treatment with 500 µL TENS lysis buffer (0.1 M NaCl, 10 mM Tris, 1 mM EDTA, and 1% SDS) and 250 µg proteinase K per 10-cm dish. Following an overnight incubation in a 1.5-mL centrifuge tube at 55°C and a 5-min denaturation of proteinase K at 95°C, genomic DNA was extracted using phenol:chloroform (1:1) in Eppendorf® Gel Light 2-mL tubes. DNA was then precipitated, and pellets were resuspended in 50 µL water. Wild-type and mutant DNA were amplified by PCR using primers listed in . The conditions for the PCR included an initial denaturation for 10 min at 94°C, followed by 35 cycles of 15 s at 94°C, 14 s at 54°C, and 30 s at 72°C. These cycles were followed by one 10-min hold at 72°C. Samples were then visualized by 2% agarose gel electrophoresis. One band of DNA, visible at 280 bp, is indicative of a HO-1 mutant-derived (−/−) cell line. A single band at 160 bp is indicative of a wild-type-derived (+/+) cell line, while two bands, one at 280 bp and one at 160 bp, are indicative of a heterozygous (+/−) genotype.
Table 1. Primers Used for PCR
Cell Proliferation and Caspase-3 Activity
MAECs (1000 cells/well) were plated in 96-well plates in complete EBM media, and cell proliferation was evaluated at 1, 3, and 5 days using the tetrazolium-based assay, XTT (Roche Diagnostics, Indianapolis, IN, USA) according to the manufacturer's instructions.
For the caspase-3 assay, confluent MAECs grown in 12-well dishes were exposed to oxidized linoleic acid (13-HPODE; L), prepared by oxidizing linoleic acid (Sigma) using soybean lipoxygenase (Sigma). Oxidation was monitored continuously on a spectrophotometer at 234 nm for conjugated diene formation as previously described (Citation11). MAECs were exposed to a concentration of 50 µM 13-HPODE for 24 h, and caspase activity was determined with the EnzChek® Caspase-3 Assay Kit (Molecular Probes, Eugene, OR, USA). Briefly, cells were washed in 500 µL PBS, centrifuged at 300× g for 5 min, the supernate was aspirated, and cell pellets were placed overnight at −20°C. The pellets were then resupended in 25 µL of 1× cell lysis buffer. Protein concentration of a 1/50 dilution of lysate was measured using the BCA procedure. Five microliters of cell lysis was combined with Z-DEVD-rhodamine 110 substrate on ice and brought up to a final volume of 12 µL in a buffer consisting of 10 mM PIPES, 10 mM dithiothreitol, and 0.1% CHAPS, pH 7.1. Kinetic measurements of caspase activity were recorded using a Model FL600 microplate fluorescent reader (Bio-Tek Instruments, Winooski, VT, USA). The cleavage of the rhodamine substrate was determined using a 485-nm excitation filter and a 530-nm emission filter at a temperature of 25°C. Rate of caspase-3 cleavage was determined by calculating the slope at the last readings that substrate was in excess, and the conversion of relative fluorescent units (RFU)/min to nanomoles DEVD substrate cleaved/min/mg of protein of the lysate.
Statistical Analysis
Statistical significance was calculated using analysis of variance (ANOVA) and the Students Neuman Keuls test. Experiments were performed in triplicate and repeated at least three times from independent mice of each genotype. Data are expressed as significant at P values of <0.05.
Results and discussion
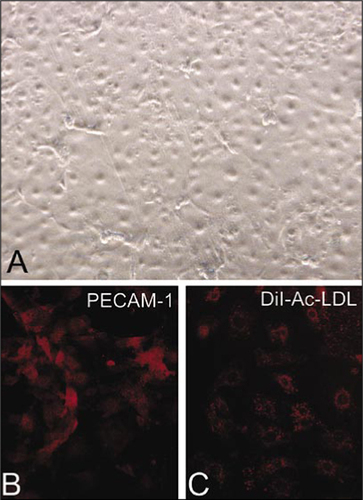
Due to the relatively small size of the mouse compared to larger animals, it is more difficult to isolate and culture mouse endothelial cells. Most investigators use methods that involve digestion of vascular segments followed by selection with magnetic beads or flow cytometric sorting with endothelial cell-specific markers (Citation1–6,Citation8,Citation12). Antibody-antigen binding or fluorescent labeling, mechanical sorting, and other factors used in this technique may alter endothelial cell physiology. We describe a simple and reproducible technique that involves gentle lumen digestion of the mouse aorta with trypsin for the isolation of pure endothelial cells. Given the biological importance of HO-1 gene expression in several cardiovascular diseases, including atherosclerosis, vascular restenosis, and hypertension (Citation13–20), we selected HO-1 deficient mice in our studies. shows the characteristic cobblestone appearance on differential interference contrast microscopy of a confluent monolayer of endothelial cells derived from a HO-1 +/− mouse. Subconfluent MAECs were stained for the endothelial cell surface marker, CD-31 (PECAM-1), and demonstrated significant positive staining in more than 95% of the cells (). MAECs also showed significant uptake of DiI-Ac-LDL, as shown in . Both PECAM-1 staining and DiI-Ac-LDL uptake were observed in more than 95% of the cells. The cells were negative for staining with anti-α-smooth muscle actin antibodies (data not shown). These results provide evidence that the isolated cells are endothelial cells.
(A) Differential interference contrast microscopy of a confluent monolayer of MAECs while in plastic tissue culture dishes. (B) Subconfluent MAECs grown on glass cover slips were stained with an endothelial surface marker, platelet endothelial cell adhesion molecule-1 (PECAM-1), using anti-CD31 antibody (primary) and a Cy3-labeled secondary antibody. (C) Uptake of DiI-Ac-LDL in subconfluent MAECs grown on glass cover slips. DiI-Ac-LDL, 1,1′-dioctadecyl-1,3,3,3′,3′-tetramethyl-indocarbocyanine-acetylated lowdensity lipoprotein.
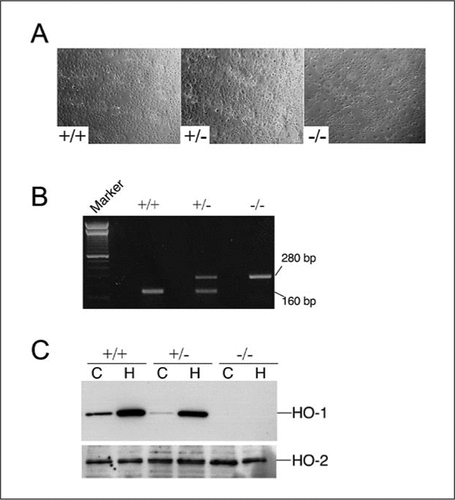
In addition to PECAM-1 and fluorescent LDL uptake, we have further characterized the MAEC cultures from HO-1 +/+, +/−, and −/− mice by PCR on genomic DNA, Western blot analysis, and cell growth (). is a representative phase-contrast image of MAECs in culture from HO-1 +/+, +/−, or −/− mice following three to four passages. We performed PCR on genomic DNA isolated from these cells to verify their genotype (). To confirm the presence or absence of HO-1, MAECs were exposed to PBS (control; C) or 5 µM hemin (H), an inducer of HO-1 for 16 h. Western blot analysis revealed significant induction of HO-1 protein in MAECs from +/+ and +/− mice, but no basal or induced HO-1 was observed in the MAECs derived from the HO-1 −/− mice (). The blot was also probed for HO-2, the constitutive isoform with an anti-HO-2 antibody, and shows no significant difference between both basal and induced levels in all three genotypes (, lower panel).
(A) Phase-contrast microscopy of confluent MAECs from HO-1 +/+, +/−, and −/− mice. (B) PCR on genomic DNA from MAECs to confirm the genotype of the cells. Marker, 100-bp DNA ladder (Invitrogen, Carlsbad, CA, USA). (C) Cells were exposed to PBS (control) or 5 µlM hemin for 16 h, and Western blot analysis was performed using anti-HO-1 or anti-HO-2 antibodies. C, control; H, hemin.
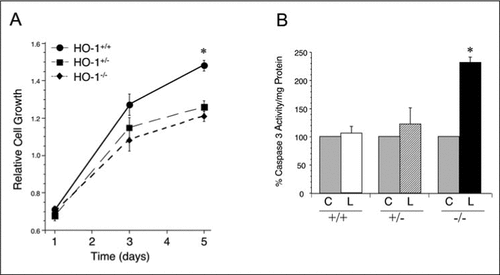
To evaluate the growth characteristics of the MAECs from the HO-1 +/+, +/−, and −/− mice, we performed cell proliferation studies using the XTT assay. As shown in , significant differences were observed in cell growth in the HO-1 +/+ versus the HO-1 +/− and HO-1 −/− cell lines. At days 3 and 5, MAECs from the HO-1 +/− and HO-1 −/− mice demonstrated slower cell growth compared to the HO-1 +/+ cells (n = 3 in each group, P < 0.05). To evaluate the susceptibility to oxidized lipid-mediated cell injury, MAECs from HO-1 +/+, +/−, or −/− mice were exposed to PBS (control; C) or 50 µM oxidized linoleic acid (13-HPODE; L) for 24 h, and caspase-3 activity was determined as a measure of cell injury. Significant increase in caspase 3 activity was observed in the HO-1 −/− MAECs exposed to 13-HPODE () (n = 3 in each group, P < 0.05), while minimal increases were seen in the HO-1 +/+ or HO-1 +/− MAECs, suggesting that the absence of HO-1 leads to increased susceptibility to cell injury.
(A) Cell growth was assessed at day 1, 3, and 5 in MAECs derived from HO-1 +/+, +/−, and −/− mice using the tetrazolium-based cell proliferation assay, XTT. (B) Confluent MAECs were exposed to PBS (control; C) or 50 µM oxidized linoleic acid (L) for 24 h, and caspase-3 activity/mg protein was determined. Gray bars, control MAECs from HO-1 +/+, +/−, or −/− mice; open bars, HO-1 +/+ MAECs; striped bars, HO-1 +/− MAECs; and dark bars, HO-1 −/− MAECs. The results in panels A and B are an average from three independent experiments, with each experiment having MAECs pooled from two individual mice from each genotype. Values are the mean (±sem). PBS, phosphate-buffered saline; *, P < 0.05.
The results of the present study demonstrate the feasibility of a simple lumen digestion technique for the isolation and culture of MAECs from transgenic mice and would be of interest to the study of cardiovascular diseases. The described procedure does not result in contamination with smooth muscle cells nor does it require manipulations for cell sorting with magnetic beads or flow cytometry. Previous studies by Suh et al. (Citation7) cultured endothelial cells from small pieces of mouse aorta directly placed on matrigel enriched with endothelial growth factors. Cells were studied on the matrigel after removing the pieces of aorta or after serial passage by using dispase and reseeding the cells. The difficulty of this technique is to control the specific culture conditions to prevent other cells, such as smooth muscle cells, from growing out. However, this technique can be useful for experiments such as patch clamping, where single endothelial cells can be selected from a mixed population of cells.
During the isolation process of this technique, the enzyme (trypsin) comes in contact only with the cells lining the inner surface of the aorta, which provides a pure population of endothelial cells. Overdigestion with trypsin could result in release of smooth muscle cells beneath the endothelium, but this can be minimized by shortening the time of enzyme digestion or by decreasing the concentration of trypsin. The isolation and subsequent culture of these cells allows for an adequate number of cells for in vitro experiments as shown in our report.
In summary, a simple yet reproducible method for isolating primary endothelial cells from mouse aorta of wild-type and gene knockout mice is reported. The method will provide pure endothelial cells with their specific genetic characteristics in vitro for further evaluation and study of endothelial cells in cardiovascular diseases.
Acknowledgments
We are grateful to Elaine Beem and Jairo Garcia for technical assistance. This work was supported by National Institutes of Health grant nos. R01 DK59600 and R01 HL68157 (to A.A.).
References
- Dong, Q.G., S.Bernasconi, S.Lostaglio, R.W.De Calmanovici, I.Martin-Padura, F.Breviario, C.Garlanda, S.Ramponi, et al.. 1997. A general strategy for isolation of endothelial cells from murine tissues. Characterization of two endothelial cell lines from the murine lung and subcutaneous sponge implants. Arterioscler. Thromb. Vasc. Biol.17:1599–1604.
- Gerritsen, M.E., C.P.Shen, W.J.Atkinson, R.C.Padgett, M.A.Gimbrone, Jr., and D.S.Milstone. 1996. Microvascular endothelial cells from E-selectin-deficient mice form tubes in vitro. Lab. Invest.75:175–184.
- Gerritsen, M.E., C.P.Shen, M.C.McHugh, W.J.Atkinson, J.M.Kiely, D.S.Milstone, F.W.Luscinskas, and M.A.Gimbrone, Jr. 1995. Activation-dependent isolation and culture of murine pulmonary microvascular endothelium. Microcirculation2:151–163.
- Kevil, C.G. and D.C.Bullard. 2001. In vitro culture and characterization of gene targeted mouse endothelium. Acta Physiol. Scand.173:151–157.
- Magid, R., D.Martinson, J.Hwang, H.Jo, and Z.S.Galis. 2003. Optimization of isolation and functional characterization of primary murine aortic endothelial cells. Endothelium10:103–109.
- Shi, W., M.E.Haberland, M.L.Jien, D.M.Shih, and A.J.Lusis. 2000. Endothelial responses to oxidized lipoproteins determine genetic susceptibility to atherosclerosis in mice. Circulation102:75–81.
- Suh, S.H., R.Vennekens, V.G.Manolopoulos, M.Freichel, U.Schweig, J.Prenen, V.Flockerzi, G.Droogmans, and B.Nilius. 1999. Characterisation of explanted endothelial cells from mouse aorta: electrophysiology and Ca2+ signalling. Pflugers Arch.438:612–620.
- Wu, Z., F.M.Hofman, and B.V.Zlokovic. 2003. A simple method for isolation and characterization of mouse brain microvascular endothelial cells. J. Neurosci. Methods130:53–63.
- Poss, K.D. and S.Tonegawa. 1997. Heme oxygenase 1 is required for mammalian iron reutilization. Proc. Natl. Acad. Sci. USA94:10919–10924.
- Feng, D., J.A.Nagy, K.Pyne, H.F.Dvorak, and A.M.Dvorak. 2004. Ultrastructural localization of platelet endothelial cell adhesion molecule (PECAM-1, CD31) in vascular endothelium. J. Histochem. Cytochem.52:87–101.
- Agarwal, A., F.Shiraishi, G.A.Visner, and H.S.Nick. 1998. Linoleyl hydroperoxide transcriptionally upregulates heme oxygenase-1 gene expression in human renal epithelial and aortic endothelial cells. J. Am. Soc. Nephrol.9:1990–1997.
- Barkalow, F.J., M.J.Goodman, and T.N.Mayadas. 1996. Cultured murine cerebral microvascular endothelial cells contain von Willebrand factor-positive Weibel-Palade bodies and support rapid cytokine-induced neutrophil adhesion. Microcirculation3:19–28.
- Durante, W. and A.I.Schafer. 1998. Carbon monoxide and vascular cell function (Review). Int. J. Mol. Med.2:255–262.
- Exner, M., M.Schillinger, E.Minar, W.Mlekusch, G.Schlerka, M.Haumer, C.Mannhalter, and O.Wagner. 2001. Heme oxygenase-1 gene promoter microsatellite polymorphism is associated with restenosis after percutaneous transluminal angioplasty. J. Endovasc. Ther.8:433–440.
- Ishikawa, K., D.Sugawara, X.Wang, K.Suzuki, H.Itabe, Y.Maruyama, and A.J.Lusis. 2001. Heme oxygenase-1 inhibits atherosclerotic lesion formation in ldl-receptor knockout mice. Circ. Res.88:506–512.
- Juan, S.H., T.S.Lee, K.W.Tseng, J.Y.Liou, S.K.Shyue, K.K.Wu, and L.Y.Chau. 2001. Adenovirus-mediated heme oxygenase-1 gene transfer inhibits the development of atherosclerosis in apolipoprotein E-deficient mice. Circulation104:1519–1525.
- Kaneda, H., M.Ohno, J.Taguchi, M.Togo, H.Hashimoto, K.Ogasawara, T.Aizawa, N.Ishizaka, and R.Nagai. 2002. Heme oxygenase-1 gene promoter polymorphism is associated with coronary artery disease in Japanese patients with coronary risk factors. Arterioscler. Thromb. Vasc. Biol.22:1680–1685.
- Otterbein, L.E., B.S.Zuckerbraun, M.Haga, F.Liu, R.Song, A.Usheva, C.Stachulak, N.Bodyak, et al.. 2003. Carbon monoxide suppresses arteriosclerotic lesions associated with chronic graft rejection and with balloon injury. Nat. Med.9:183–190.
- Tulis, D.A., W.Durante, K.J.Peyton, A.J.Evans, and A.I.Schafer. 2001. Heme oxygenase-1 attenuates vascular remodeling following balloon injury in rat carotid arteries. Atherosclerosis155:113–122.
- Yet, S.F., M.D.Layne, X.Liu, Y.H.Chen, B.Ith, N.E.Sibinga, and M.A.Perrella. 2003. Absence of heme oxygenase-1 exacerbates atherosclerotic lesion formation and vascular remodeling. FASEB J.17:1759–1761.

