
The Immortomouse™ (Citation1) harbors a transgene that expresses the thermolabile simian virus 40 (SV40) large T antigen tsA58 under the control of the interferon-inducible murine H-2Kb promoter. This allows the isolation of conditionally immortalized cell lines directly from the transgenic mouse. These cells undergo reversible immortalization when grown under permissive conditions (33°C, with interferon-γ) and stop dividing and differentiate at higher temperatures in the absence of interferon. To date, cell lines derived from the Immortomouse include inner ear hair cells (Citation2), kidney collecting tubuli cells (Citation3), skeletal myoblasts (Citation4), hepatocytes (Citation5), and colonocytes (Citation6). However, the exploitation of this mouse model—particularly for any purposes requiring crossbreeding with other transgenic mice—has been hampered by the fact that no molecular test existed for the distinction of hemizygous and homozygous Immortomice.
A single allele of the transgene is sufficient to immortalize cells (Citation4), and the mice can be obtained from Charles River Laboratories (Wilmington, MA, USA) either as homozygotes or as F1-hemizygotes. Testing for the H-2Kb-tsA58 transgene can easily be accomplished using PCR. However, because the localization of the transgene was not known, until now it was not possible to test for the corresponding wild-type allele and thus identify hemizygotes. The alternative approach, to determine the genotype by backcrossing, is cumbersome and inappropriate for this mouse model because Immortomice have a reduced fertility and lifespan due to thymic hyperplasia (Citation1). Thus, homozygosity of the transgene could hardly ever be restored in a colony after crossbreeding with any other mouse strain.
To localize the transgene, we determined the 3′-flanking region using the Universal GenomeWalker™ Kit (BD Biosciences Clontech, Palo Alto, CA, USA). High-weight genomic DNA was isolated from mousetail clips and digested in aliquots with three blunt-end-cutting restriction enzymes (EcoRV, PvuII, StuI). The restriction fragments were ligated to blunt adaptors provided with the kit and used as templates for PCR. The forward primer was located near the 3′ end of the tsA58 sequence, the reverse primer (AP1) in the adaptor sequence, and a long-distance polymerase with proofreading capability (Advantage™ Polymerase; BD Biosciences Clontech) was used. The PCR products were reamplified in a second PCR with nested primers. The StuI-digested template DNA produced a single band of approximately 1.3 kb in length, which was sequenced from the 5′ end (MWG-Biotech, Munich, Germany). A Basic Local Alignment Search Tool (BLAST) search (http://www.ensembl.org; mouse genome v22.32b1) localized the sequence corresponding to the PCR product in band B5 of mouse chromosome 16 (). The 3′ end of the tsA58 gene is located 1.4 kb upstream of the chromosomal marker D16Mit30 (). Currently, no gene is predicted at this site that could have been disrupted by the insertion of the transgene.
Initial attempts to amplify the 5′-flanking region by PCR failed because part of the wild-type genome apparently had been deleted during the insertion of the transgene. We estimated the size of the deleted sequence by testing several primer pairs located upstream of the insertion site, using wild-type mouse DNA as a positive control. Next, the 5′-transition was identified with a primer pair spanning the sequence between the verified wild-type genomic region and the H-2Kb promoter. The PCR product was sequenced from the 3′ end as described above.
Analysis of the transition sequences showed that both ends of the transgene had been inserted without truncation (). About 7.3 kb of the wild-type mouse genome are deleted at the insertion site, which corresponds roughly to the size of the 6.9-kb transgene. Aside from a short region with tandem repeats upstream of the transgene promoter and from a few minor deletions and base exchanges, the flanking sequences are in line with the sequence published at http://www.ensembl.org for at least 500 bp upstream and 270 bp downstream. For complete alignments of the wild-type and H-2Kb-tsA58 insertion site, see the supplementary material.
(A) Alignment of the wild-type (upper) and the transgene (lower) allele and sequence alignments of wild-type (WT; data from http://www.ensembl.org), Immortomouse (Imm), and the H-2Kb-tsA58 transgene (EcoRI-BamHI fragment) at the transition sites. The insertion site is located 1.4 kb upstream of the chromosomal marker D16Mit30. At the insertion site, 7.3 kb of the mouse genome are deleted (indicated by dashed line in the wild-type allele). The locations of the three primers used in the mixed genotyping test (wt-f, tsA-f, and wt-r) are indicated by arrows. (B) Banding pattern of mouse chromosome 16 with the location of the H-2Kb-tsA58 transgene indicated in B5 (arrow). (C) PCR gel banding patterns of wild-type (lanes 1, 3, and 6), hemizygous (lanes 4, 5, and 7), and homozygous (lane 2) Immortomice tested with the newly developed genotype assay.
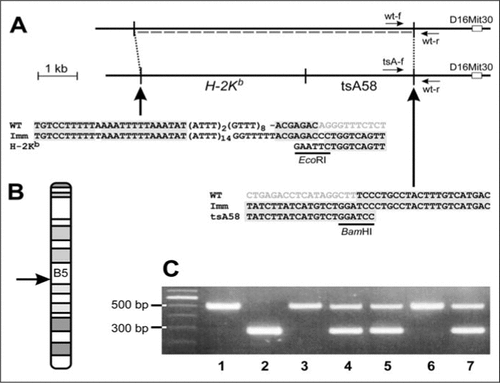
Knowing the sequences of the flanking regions enabled us to develop a PCR-based genotype test for the Immortomouse. We designed three primers with identical melting temperatures (Tms; approximately 60°C) to simultaneously determine the wild-type allele (primers wt-f and wt-r; product approximately 500 bp) and the H-2Kb-tsA58 allele (tsA-f and wt-r; approximately 300 bp) in a single PCR. Primer sequences were 5′-GATCTGCCTGAGGTGTTACTTG-3′ for wt-f, 5′-GGATGGCATCACTAGTCATGAC-3′ for wt-r, and 5′-AGTCCTCACAGTCTGTTCATGATC-3′ for tsA-f (for primer positions, see ).
The PCR assay showed high fidelity with DNA extracted from tail ends using the HotSHOT method (; Reference (Citation7). The results were validated by reexamination of approximately 80 H-2Kb-tsA58-positive mice. In all cases, genotypes deduced from breeding schemes and previous testing were confirmed.
The only reported health impairment in the Immortomouse is thymic hyperplasia, which impedes respiratory movements. Homozygotes are more affected by these symptoms and thus have a significantly reduced lifespan (Citation1). In addition, we found a decreased fertility in homozygous females. The genotype test allowed us to assess the affectedness in hemizygous and homozygous mice. While about two thirds (47 of 72) of the hemizygous females gave birth up to three times, only one fifth (3 in 15) of the homozygous females reproduced, and all of them only once. Homozygous male mice were generally less affected than females. Therefore, the most effective breeding routine in our hands was mating homozygous males with hemizygous females. Finally, the inheritance of the transgene showed a clear gender-dependence. Hemizygous breeding pairs delivered female offspring at a ratio of 16% homozygotes, 58% hemizygotes, and 26% wild-type, and male offspring at a ratio of 32%, 47%, and 21%, respectively (166 mice, genotyped at the age of 5 weeks). Apparently, homozygous females have a reduced viability during embryonic and/or early postnatal development.
In fibroblasts isolated from the Immortomouse, the ability to form colonies and the conditionality of proliferation are dependent on tumor antigen levels (Citation1). In our experiments, immortalization was reversible in both hemizygous and homozygous myogenic cultures, and myoblasts could be induced to differentiate into a skeletal muscle phenotype. However, initial observations suggest that hemizygous cell lines more closely resembled the phenotype of primary mouse myoblasts (spherical myoblasts, earlier fusion, and differentiation), while homozygous cell lines were characterized by higher proliferation rates and flatter myotubes. This indicates that specific characteristics of the resulting immortalized cell lines can be achieved by the controlled production of both hemizygous and homozygous Immortomouse cell lines.
Thus, because the sequencing of the mouse genome is almost complete, PCR-based genomic walking is an efficient method for the precise localization of transgenes. Determining the insertion site of the H-2Kb-tsA58 transgene provided the basis for the development of a simple and reliable PCR-based genotype assay to distinguish homozygous and hemizygous Immortomice. Crossing hemizygous females with homozygous males makes breeding easier and more predictable, while still supplying homozygous offspring. Most importantly, the novel genotype test makes it possible to produce proven H-2Kb-tsA58 homozygous mice after the crossbreeding with other transgenic or knock-out mice.
Competing Interests Statement
The authors declare no competing interests.
Alignment of the Immortomouse sequence with wildtype (from www.ensembl.org) and transgene promoter (H-2Kb) sequences at the 5’-transition site:
Download PDF (71.8 KB)Acknowledgments
We thank Dr. F. Moebius for providing breeding pairs, Ms. M. Angebrand for excellent technical help, and Dr. M. Grabner for helpful discussion. This work was funded in part by grants from the Austrian Science Fund (FWF) and the Austrian National Bank (OeNB) P16532-B05, and by a grant from the European Commission HPRN-CT-2002-00331 to B.E.F.
Supplementary data
To view the supplementary data that accompany this paper please visit the journal website at: www.tandfonline.com/doi/suppl/10.2144/05381BM03
Additional information
Funding
References
- Jat, P.S., M.D.Noble, P.Ataliotis, Y.Tanaka, N.Yannoutsos, L.Larsen, and D.Kioussis. 1991. Direct derivation of conditionally immortal cell lines from an H-2Kb-tsA58 transgenic mouse. Proc. Natl. Acad. Sci. USA88:5096–5100.
- Holley, M.C., Y.Nishida, and N.Grix. 1997. Conditional immortalization of hair cells from the inner ear. Int. J. Dev. Neurosci.15:541–552.
- Takacs-Jarrett, M., W.E.Sweeney, E.D.Avner, and C.U.Cotton. 1998. Morphological and functional characterization of a conditionally immortalized collecting tubule cell line. Am. J. Physiol.275:F802–F811.
- Morgan, J.E., J.R.Beauchamp, C.N.Pagel, M.Peckham, P.Ataliotis, P.S.Jat, M.D.Noble, K.Farmer, et al.. 1994. Myogenic cell lines derived from transgenic mice carrying a thermolabile T antigen: a model system for the derivation of tissue-specific and mutation-specific cell lines. Dev. Biol.162:486–498.
- Allen, K.J., R.Reyes, K.Demmler, J.F.Mercer, R.Williamson, and R.H.Whitehead. 2000. Conditionally immortalized mouse hepatocytes for use in liver gene therapy. J. Gastroenterol. Hepatol.15:1325–1332.
- Whitehead, R.H., P.E.VanEeden, M.D.Noble, P.Ataliotis, and P.S.Jat. 1993. Establishment of conditionally immortalized epithelial cell lines from both colon and small intestine of adult H-2Kb-tsA58 transgenic mice. Proc. Natl. Acad. Sci. USA90:587–591.
- Truett, G.E., P.Heeger, R.L.Mynatt, A.A.Truett, J.A.Walker, and M.L.Warman. 2000. Preparation of PCR-quality mouse genomic DNA with hot sodium hydroxide and tris (HotSHOT). BioTechniques29:52–54.

