Abstract
Shwachman-Bodian-Diamond syndrome (SBDS) is linked to a mutation in a single gene. The SBDS proinvolved in RNA metabolism and ribosome-associated functions, but SBDS mutation is primarily linked to a defect in polymorphonuclear leukocytes unable to orient correctly in a spatial gradient of chemoattractants. Results of data mining and comparative genomic approaches undertaken in this study suggest that SBDS protein is also linked to tRNA metabolism and translation initiation. Analysis of crosstalk between translation machinery and cytoskeletal dynamics provides new insights into the cellular chemotactic defects caused by SBDS protein malfunction. The proposed functional interactions provide a new approach to exploit potential targets in the treatment and monitoring of this disease.
Introduction
Shwachman-Bodian-Diamond syndrome (SBDS) is an autosomal recessive disorder characterized by pancreatic exocrine insufficiency, bone marrow dysfunction, and skeletal abnormalities.Citation1
The SBDS gene (sbds) encodes a member of a highly conserved protein family of unknown function, with orthologs in diverse species, including Archaea, plants, and eukaryotes, but not Eubacteria.Citation2 Structural and functional aspects of SBDS link this protein to a group of proteins involved in RNA metabolism with ribosome-associated functions.Citation3,Citation4 For example, the yeast SBDS ortholog clusters with RNA-processing enzymes and ribosomal RNA-processing factors in gene expression analyses,Citation5,Citation6 and in the SBDS sequence homolog in Saccharomyces cerevisiae, YLR022C, was shown to be physically associated with proteins involved in ribosome biosynthesis.Citation7
The N-terminal domain of the SBDS protein, which is most prone to mutations leading to disease, contains a novel mixed α/β fold that was also suggested for a single domain yeast protein, Yhr087wp, implicated in RNA metabolism.Citation1 The βαββαβ folding topology of the SBDS C-terminal domain III (Glu162–Gly234) is structurally close to domain V of S. cerevisiae elongation factor 2 (Eft2p/Ydr385wp).Citation8 It is also typical for a ferredoxin-like fold and characteristic of a number of heterogeneous nuclear ribonucleoproteins, eg, factors involved in regulation of alternative splicing.Citation9 Although SBDS links to RNA metabolism are apparent, an association has also been observed between the SBDS protein and cell locomotion, in that polymorphonuclear leukocytes isolated from SBDS patients are unable to orient correctly in a spatial gradient of chemoattractants.Citation10 SBDS protein was also shown to localize to the pseudopod of Dictyostelium amoebae during chemotaxis.Citation11 The interaction of SBDS and a structural or regulatory cytoskeletal component is more than likely responsible for the observed defect in polymorphonuclear leukocyte chemotaxis. Nonetheless, no specific candidate for such an interaction has been suggested.
The complexities of the myriad defects associated with SBDS have made it difficult to relate the diverse biochemical and phenotypic properties of the SBDS syndrome on an experimental basis. A way forward on how mutations of the SBDS gene (sbds) may relate to its altered functions is via a functional genomics approach and phylogenetic analysis of gene proximity in sbds loci. Functionally related genes are commonly found clustered in prokaryotic and eukaryotic genomes,Citation12–Citation17 and predicting gene function based on physical proximity to other genes has been used successfully in a number of studies. Therefore, we treated consistency of gene proximity in sbds loci in evolutionary distant genomes as an indication of functional relatedness, which led to a prediction of SBDS protein involvement in initiation of translational wybutosine metabolism. The crosstalk between the translation machinery and elements of the cytoskeleton provides an explanation as to how cell chemotactic defects may be caused by SBDS malfunction.
Materials and methods
We used the Seed database (http://theseed.uchicago.edu/FIG/index.cgi) for chromosome alignment and phylogenetic analysis of gene positional clusters.Citation18 The “Compare Regions” resource provided by Seed allows alignment of chromosome loci that contain open reading frames for homologous proteins, or, in other words, to pin these loci through genes that are homologous to a query sequence. It can be used in a text or graphic format. We used the latter to illustrate phylogenetic conservation of gene proximity. The typical graphic window presents a selected number of chromosome loci from different genomes. The first line of Compare Regions is a graphical display of the chromosomal neighborhood of the features in its genome. All proteins are shown as colored arrows, where the direction depicts the strand of the feature. RNAs and other features are small boxes on the line. Feature overlaps are resolved by drawing the overlapping feature in a new line. The graph is centered on the selected feature, always numbered 1 and colored red. Below, there is the same region for orthologs in other organisms, also colored in red. The colors of the other features (as well as the numbers) also represent ortholog (or sometimes also paralog) features. Whenever there are at least two ortholog or paralog features of a kind, a color (and a number) is assigned to them. The selection of genomes to show in the graphics can be made by similarity or the pair of close homologs pin. We used similarity, which means that the genomes are chosen using the similarity of the selected genes to its orthologs in other genomes. The E value cutoff for selection of pinned coding sequence depicts the minimum similarity in order for its region to be displayed. We used the e-20 E value threshold to obtain all the presented data sets.
There are numerous queering and display options that allow customization of the size of displayed regions, selection of organisms, similarity thresholds for pinning of regions, and coloring of features that we implemented to deliver the illustrations accompanying this paper.
Results
Conservation of gene proximity in SBDS gene loci
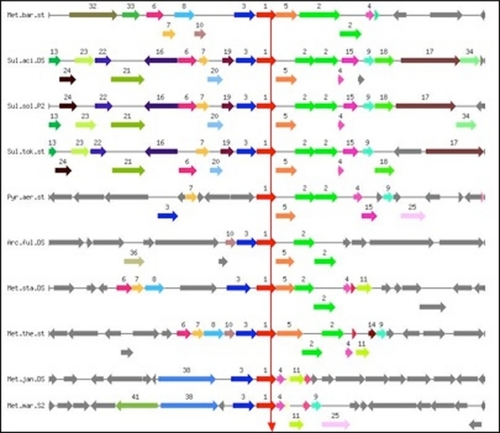
Phylogenetic analysis of archeal sbds loci () shows conservation of sbds gene proximity. Sbds orthologs are shown as red arrows (N1) in the centers of all the selected regions, where one can also see repetitive occurrence of colors/numbers depicting other orthologous genes in different genomes. Almost all of these co-occurring genes are related to RNA modification and degradation, ie, probable exosome complex exonuclease 2 (EC 3.1.13.-)/tRNA nucleotidyltransferase (N2), proteasome subunit α (EC 3.4.25.1) (N3), probable exosome complex RNA-binding protein 1 (N4), large ribosomal subunit protein L37 Ae (N5) large ribosomal subunit protein L15e (N7), ribonuclease P (tRNA processing) protein component 3 (EC 3.1.26.5) (N8), ribonuclease P protein component 2 (EC 3.1.26.5) (N9), prefoldin, chaperonin cofactor (N10), and a predicted exosome subunit containing the IMP4 domain present in small nuclear ribonucleoprotein (N11). An archeal sbds locus that includes all or part of the genes encoding the listed functions is surrounded by a variable region (gray arrows), suggesting that clustered genes related to the archeal exosome complex indeed represent a functionally coupled group or even an operon, and that sbds can be a part of this group.
Figure 1 Graphical display of the chromosomal neighborhood of sbds genes in archeal genomes. Arrows correspond to open reading frames, same color and number depicts gene orthology. Vertical arrow connects pinned genes (red arrows), ie, orthologs of sbds gene from Methanosarcina barkeri (upper line). Genes encode 1-SBDS protein (AAB90746.1 and its orthologs), 2-probable exosome complex exonuclease 2 (EC 3.1.13.-)/tRNA nucleotidyltransferase, 3-proteasome subunit α (EC 3.4.25.1), 4-probable exosome complex RNA-binding protein 1, 5-large ribosomal subunit protein L37 Ae, 7-large ribosomal subunit protein L15e, 8-ribonuclease P (tRNA processing) protein component 3 (EC 3.1.26.5), 9-ribonuclease P protein component 2 (EC 3.1.26.5), 10-prefoldin, chaperonin cofactor, and 11-predicted exosome subunit containing the IMP4 domain present in small nuclear ribonucleoprotein. 16 kbp regions are shown.
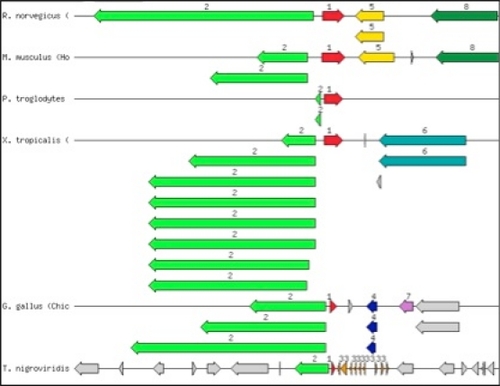
Chromosome loci with vertebrate orthologs to human sbds (ENSG00000126524) have also been aligned. In the human genome, sbds spans a start for tyw1 (ENSG00000198874), which encodes tRNA-wybutosine synthesizing protein (TYW1) from the opposite string. In all vertebrate genomes analyzed (from the SEED database collection), sbds (red, N1) is colocalized with a gene encoding the TYW1 homolog, annotated in SEED as wybutosine biosynthesis reductase (green, N2, ). A small gap between these two genes in the different vertebrate genomes suggests that they may share a regulatory region. The gene region downstream of sbds is not conserved, even in narrower phylogenetic groups, because one can see it for the displayed regions from several mammalian genomes.
Figure 2 Graphical display of the chromosomal neighborhood of sbds genes in genomes of vertebrates. Arrows correspond to open reading frames encoding: red, Shwachman-Bodian-Diamond syndrome (NM_001008289.1 and pinned orthologs, green), tRNA-YW synthesizing protein (NP_001100607.1 (in Rattus norvegicus) and ortholog, multiple arrows in Xenopus tropicalis correspond to known transcripts), 3-claudin, 4-septin, 5-0610007 L01Rik protein, 6-hypothetical protein, 7-putative dual specificity testis-specific protein kinase 2 (EC 2.7.11.1), and 8-RABX5 (NP_001185988.1). 200 kbp regions are shown. Organisms are R. norvegicus, Mus musculus, Pan troglodytes, X. tropicalis, Gallus gallus, Tetraodon nigroviridis.
In the Plasmodium falciparum and Ciona intestinalis genomes, sbds genes are also colocalized with genes encoding functions related to translational initiation and mainly with tRNA-regulated components of translational initiation complexes, ie, a gene for predicted GCN1 (general control of amino-acid synthesis 1) (XP_002120729.1) is immediately downstream and a gene for tRNA/RNA cytosine-C5-methylase (EC 2.1.1.-) is immediately upstream from the human sbds ortholog (XM_002124782.1) in the C. intestinalis genome (). The aIF2β gene is the second gene upstream from the sbds ortholog (XP_001348280.1) in a Plasmodium genome ().
Table 1 Tabular display of the chromosomal neighborhood of sbds gene in Ciona intestinalis genome (SEED db IDs)
Table 2 Tabular display of the chromosomal neighborhood of sbds gene in Plasmodium falciparum genome (SEED db IDs)
In Schizosaccharomyces pombe, the sbds ortholog (O14179, SDO1) is proximal to a gene encoding the L26e ribosomal protein, followed by a gene for the mRNA capping enzyme subunit. Gene encoding RNA processing factor 1 is immediately downstream, followed by one for S23e ().
Table 3 Tabular display of the chromosomal neighborhood of SBDS-encoding genes in Schizosaccharomyces pombe genome (SEED db IDs)
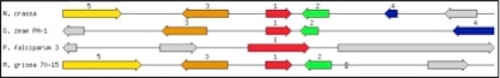
No specific functional gene overrepresentation at the sbds locus in S. cereviciae was observed, apart from a noticeable but comparatively distant proximity to genes encoding RNA-related functions. However, sbds orthologs (XP_387070.1, XP_956795.1, XP_360288.1) are proximal to genes encoding L18e in fungal (Gibberella zeae, Neurospora crassa, and Magnaporthe oryzae) genomes ().
Figure 3 Graphical display of the chromosomal neighborhood of sbds genes in fungal genomes. Arrows correspond to open reading frames encoding: 2- SSU ribosomal protein S18e (S13p), 3-carboxypeptidase precursor, and 4- and 5-hypothetical proteins. 100 kbp regions are shown. Organisms are Neurospora crassa, Gibberella zeae, Plasmodium falciparum, and Magnaporthe grisea.
Discussion
SBDS is a complex disease linked to mutation in a single gene. Recent advances in genomics and functional bioinformatics are providing a new avenue for studying this and similar phenomena, complementing traditional hypothesis-driven laboratory research. Our bioinformatics approach undertaken in this study provides a new insight as to how a single gene mutation may affect diverse molecular functions, such as RNA metabolism, translation initiation, and cytoskeletal dynamics leading to complex disease.
Phylogenetic conservation of gene proximity may manifest itself in the presence of sets of orthologous genes in gene loci from different genomes. However, even in prokaryotes, it often comes up as conservation of positional connections between and inside particular pathways, not exactly the same representatives of the pathways.Citation12,Citation18 Therefore, integration of genomic data may uncover trends in gene positional clustering and the underlying functional links between genes and pathways. Analysis of our data points to initiation of translation as the main functional connection for SBDS (), with particular TYW1 and SBDS functional coupling as characteristic of all the studied vertebrate genomes.
Wybutosine is a hypermodified guanosine with a tricyclic base found at the 3′-position adjacent to the anticodon of eukaryotic phenylalanine tRNA. The UUU phenylalanine codon is highly prone to frameshift in the 3′ (rightward) direction at pyrimidine 3′ contexts,Citation19 and wybutosine supports reading frame maintenance by stabilizing codon–anticodon interactions during decoding on the ribosome.Citation20,Citation21 Wybutosine synthesis might proceed through sequential reactions in a multiple protein complex assembled with the precursor tRNA and may be linked to exosomal/proteosomal structure that includes the SBDS protein. Comparative analysis of archeal sbds loci stresses the potential involvement of SBDS in the exosomal complex where functions of translation, RNA processing, and degradation are tightly coupled.Citation22
The SBDS protein has a thioredoxin fold, which might be directly coupled to the function of wybutosine reductase. Not all stages of wybutosine biosynthesis are known. There is some ambiguity surrounding the source of the C2 atom in wybutosine, suggested to originate from an intermediate in lysine metabolism. Interestingly, SBDS was shown to interact physically (Reactome db) with methylmalonate semialdehyde dehydrogenase, the enzyme in the branched-chain amino acid degradation pathway. It would be too speculative to go further, but the possible involvement of the SBDS protein in biosynthesis of wybutosine merits experimental validation.
Considering an alternative link between wybutosine and SBDS, we may suggest involvement of SBDS in degradation of noncharged or not properly modified tRNA. The tRNA surveillance pathway has been shown to exist in yeast and requires the exosome for polyadenylation and degradation of hypomodified pre-tRNA(i)(Met).Citation23 Analogously, tRNA(Phe) without a modified wybutosine residue may be subject to degradation, and therefore SBDS may play some role in this process. The N-terminus of the Yhr087wp yeast protein has a fold similar to that in SBDS. It also has 2.4e-06 and 100.00% homology with bacterial tRNA pseudouridine 13 synthase, that may support an involvement of Yhr087wp itself and SBDS protein in tRNA modification. Genes do not cluster in S. cerevisiae orthologs of SBDS (Q07953, SDO1) and TYW1 (Q08960, Tyw1p). However, the tyw1 locus in S. cerevisiae also contains a gene for tRNA pseudouridine synthase 1 (YPL212C) and a number of genes encoding proteins of ribosomal biogenesis.
Establishing a link between SBDS and tRNA modification provides an interesting aspect of crosstalk between cytoskeleton function and translation machinery that may explain a faulty chemotactic phenotype of SBDS malfunction.
In the P. falciparum and C. intestinalis genomes, SBDS genes are colocalized with functions associated with tRNA-regulated components of translational initiation complexes.Citation24 GCN1 is a translational activator and regulator of Gcn2p kinase activity. It forms a complex with Gcn20p and is proposed to stimulate Gcn2p activation by an uncharged tRNA.Citation24,Citation25 The second gene colocalized with sbds in the sea squid genome is a gene for tRNA/RNA cytosine C5 methylase, which is required for initiating tRNA(i) (Met) modification. Eukaryotic and archeal initiation factors 2 are heterotrimeric proteins where only the γ subunit of the aIF2αβ heterodimer contacts tRNA.Citation26 Intriguingly, the aIF2β gene is colocalized with sbds in a genome of Plasmodium ().
Indirect evidence implicates actin as a cofactor in eukaryotic protein synthesis. The principle function of EF-1α is to bind aminoacyl-tRNA to the ribosome. EF-1α also interacts with the cytoskeleton by binding and bundling actin filaments and microtubules, and can alter the assembly of F-actin, a filamentous scaffold on which nonmembrane-associated protein translation take placeCitation27 (). F-actin and aa-tRNA compete for EF-1α, and their binding is pH-dependent and mutually exclusive. Release of EF-1α from actin binding was suggested to cause a transient increase in local concentration of the factor to facilitate polypeptide elongation. This interrelationship may ensure that cell proliferation and steady-state protein synthesis is separated from cell migration caused in primitive eukaryotic ancestors by starvation or by an avoidance response to other stressors.
Figure 4 Hypothetical functions of SBDS protein, and their effect on chemotaxis. Clouds represent gene proximity in different groups of organisms, ie, vertebrates (yellow), Ciona intestinalis (blue), and Plasmodium falciparum (purple).
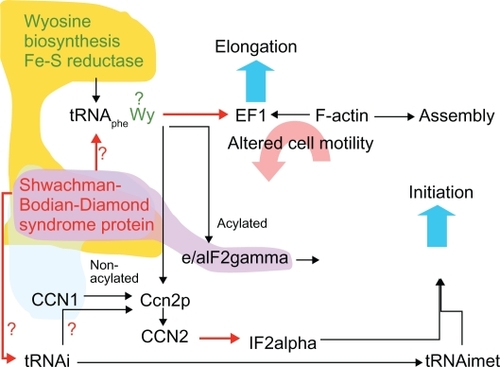
There is also a crosstalk between the two systems in establishing cell polarity during chemotaxis. It has been proposed that the EF-1α-F-actin complex is an important scaffold for anchoring of β-actin mRNA to sites of active actin polymerization.Citation28 Translation only occurs when the RNA-protein complex reaches its destination at the periphery of the cell.Citation29 Nothing is known about localization of exosome complexes to cellular lamella, but the processes of translation and degradation of RNA species are likely to be colocalized and to be coordinated by cell locomotion.
It can be suggested that binding of tRNAphe with or without wybutosine modification differentially affects EF-1α. For example, tRNAphe without wybutosine modification can bind to EF-1α with a higher affinity, and eventually lead to cell polarization/arrest of movement. In this case, the function of the SBDS protein would be in downregulation or destruction of EF-1α, thereby inhibiting tRNA(s). It was shown recently that SBDS protein is also required for release and recycling of the nucleolar shuttling factor, Tif6, from pre-60S ribosomes,Citation30 and also for pre-rRNA modifications and final maturation of the ribosome.Citation31 Both processes can be regulated and coordinated with ribosomal transport to the cell periphery where actin translation and assembly takes place.Citation32,Citation33
We suggest a multiple involvement of SBDS protein in initiation and stability of translation in vertebrates. The suggested roles for this protein in wybutosine/tRNA metabolism would complement its potential involvement in the ribosome assembly recently reported for yeast and would crosstalk tightly, with establishment of cell polarity and cell locomotion. Experimental validation of the relationship between SBDS, wybutosine synthesis, and/or degradation of tRNAphe seems plausible, and we hope that our suggestions will attract the attention of biologists in related fields.
It is still not clear what functional features of a gene pair (structural or functional specificity of the encoded proteins, topology of their interaction, presence of a direct protein–protein contact) correlate significantly with their colocalization. Validation of genomic clustering of genes encoding metabolic functions demonstrates 90% correlation for yeast and just slightly less for a human genome. A high correlation was also shown between gene colocalization and their temporal and spatial expression profiles.Citation14,Citation16 All the existing studies point to phenotypic associations between genes clustered in genomes, but more information is required for a proper large-scale statistical analysis of the correlation. We hope that case-by-case analysis will support the general validity of this method and lead to a routine automatic approach to functional classification of eukaryotic proteins via systematic comparative genomics.
Acknowledgements
The author thanks Dr Bahram Ibrahimi for his useful comments and also colleagues at the Fellowship of Interpretation of Genomes for their constant support and inspiration.
Disclosure
The author reports no conflict of interest in this work.
References
- ShammasCMenneTFHilcenkoCStructural and mutational analysis of the SBDS protein family. Insight into the leukemia-associated Shwachman-Diamond syndromeJ Biol Chem2005280192211922915701631
- BatemanABirneyECerrutiThe Pfam Protein Families DatabaseNucleic Acids Res20023027628011752314
- BoocockGRMorrisonJAPopovicMMutations in SBDS are associated with Shwachman-Diamond syndromeNat Genet2003339710112496757
- de OliveiraJFSforçaMLBlumenscheinTMStructure, dynamics, and RNA interaction analysis of the human SBDS proteinJ Mol Biol20103961053106920053358
- WuLFHughesTRDavierwalaAPRobinsonMDStoughtonRAltschulerSJLarge-scale prediction of Saccharomyces cerevisiae gene function using overlapping transcriptional clustersNat Genet20023125526512089522
- PengWTRobinsonMDMnaimnehSA panoramic view of yeast noncoding RNA processingCell200311391993312837249
- SavchenkoAKroganNCortJRThe Shwachman-Bodian-Diamond syndrome protein family is involved in RNA metabolismJ Biol Chem2005280192131922015701634
- JorgensenROrtizPACarr-SchmidANissenPKinzyTGAndersenGRTwo crystal structures demonstrate large conformational changes in the eukaryotic ribosomal translocaseNat Struct Biol20031037938512692531
- OubridgeCItoNEvansPRTeoCHNagaiKCrystal structure at 1.92 A resolution of the RNA-binding domain of the U1ANature19943724324387984237
- StepanovicVWesselsDGoldmanFDGeigerJSollDRThe chemotaxis defect of Shwachman-Diamond syndrome leukocytesCell Motil Cytoskeleton20045715817414743349
- WesselsDSrikanthaTYiSKuhlSAravindLSollDRThe Shwachman-Bodian-Diamond syndrome gene encodes an RNA-binding protein that localizes to the pseudopod of Dictyostelium amoebae during chemotaxisJ Cell Sci200611937037916410554
- R OverbeekRFonsteinMD’SouzaMPuschGDMaltsevNThe use of gene clusters to infer functional couplingProc Natl Acad Sci U S A1999962896290110077608
- VasievaOWolfRUnraveling functional networks: does gene clustering have a meaning?BMC Sys Biol.20071Suppl v184
- LeeJMSonnhammerELGenomic gene clustering analysis of pathways in eukaryotesGenome Res2003587588212695325
- VolpiEVChevretEJonesTLarge-scale chromatin organization of the major histocompatibility complex and other regions of human chromosome 6 and its response to interferon in interphase nucleiJ Cell Sci20001131565157610751148
- HurstLDPalCLercherMThe evolutionary dynamics of eukaryotic gene orderNat Rev Genet2004529931015131653
- VasievaOThe many faces of glutathione transferase piCurr Mol Med20111112913921342130
- OverbeekRBegleyTButlerRMThe subsystems approach to genome annotation and its use in the project to annotate 1000 genomesNucleic Acids Res2005335691570216214803
- SchwartzRCurranJFAnalyses of frameshifting at UUU-pyrimidine sitesNucleic Acids Res199725200520119115369
- NomaAKirinoYIkeuchiYSuzukiTBiosynthesis of wybutosine, a hyper-modified nucleoside in eukaryotic phenylalanine tRNAEMBO J2006252142215416642040
- SalomonRGiveonDKimhiYLittauerUZAbundance of tRNAPhe lacking the peroxy Y-base in mouse neuroblastomaBiochemistry19761552585262999805
- KooninEVWolfYIAravindLPrediction of the archaeal exosome and its connections with the proteasome and the translation and transcription machineries by a comparative-genomic approachGenome Res20011124025211157787
- KadabaSKruegerATriceTKrecicAMHinnebuschAGAndersonJNuclear surveillance and degradation of hypomodified initiator tRNAmet in S cerevisiaeGenes Dev2004181227124015145828
- OlsenDSSavnerEMMathewADomains of eIF1 A that mediate binding to eIF2, eIF3 and eIF5B and promote ternary complex recruitment in vivoEMBO J20032219320412514125
- KubotaHOtaKSakakiYItoTBudding yeast GCN1 binds the GI domain to activate the eIF2alpha kinase GCN2J Biol Chem2001276175911759611350982
- YatimeLMechulamYBlanquetSSchmittEStructural switch of the γ subunit in an archaeal aIF2αγ heterodimerStructure20061411912816407071
- LiuGTangJEdmondsBTMurrayJLevinSCondeelisJF-actin sequesters elongation factor from interaction with aminoacyl-tRNA in a pH-dependent reactionJ Cell Biol19961359539638922379
- LiuGGrantWMPerskyDLathamVMJrSingerRHCondeelisJInteractions of elongation factor 1a with F-actin and b-actin mRNA: Implications for anchoring mRNA in cell protrusionsMol Biol Cell20021357959211854414
- HüttelmaierSZenklusenDLedererMZBP1 enhances cell polarity and reduces chemotaxisJ Cell Sci200512031733178
- MenneTFGoyenecheaBSánchez-PuigNThe Shwachman-Bodian-Diamond syndrome protein mediates translational activation of ribosomes in yeastNat Genet20073948649517353896
- SavchenkoAKroganNCortJRThe Shwachman-Bodian-Diamond syndrome protein family is involved in RNA metabolismJ Biol Chem2005280192131922015701634
- MirraSSMilesMLJacobsJThe coexistence of ribosome-lamella complex and annulate lamellae in chronic lymphocytic leukemiaUltrastruct Pathol198122492567292629
- SiglerEShvidelLShtalridMBerribiAShaftDResnitzkyPRibosome-lamella complexes in a patient with aggressive chronic lymphocytic leukemiaLeuk Lymphoma20074843443617325913