
Abstract
Curcumin (CUR) is a compound that has antibacterial, antiviral, anti-inflammatory, and anticancer properties. In this study, we have analyzed the effects of CUR on the expression of ERα and p53 in the presence of hormones and anti-hormones in breast cancer cells. Cells were cultured in a medium containing charcoal-stripped fetal bovine serum to deplete any endogenous steroids and treated with CUR at varying concentrations or in combination with hormones and anti-hormones. Protein analysis revealed a relative decrease in the levels of p53 and ERα upon treatment with 5–60 µM CUR. In cell proliferation studies, CUR alone caused a 10-fold decrease compared with the treatment with estrogen, which suggests its antiproliferative effects. Delineating the role of CUR in the regulation of p53, ERα, and their mechanisms of action may be important in understanding the influence of CUR on tumor suppressors and hormone receptors in breast cancer.
Introduction
In the USA alone, breast cancer is the second leading cause of cancer death in women.Citation1 Invasive breast cancer (IBC) is known to develop after the progression of multiple stages, one of the later stages being referred to as ductal carcinoma in situ (DCIS). DCIS is an intraductal neoplastic proliferation of epithelial cells that do not invade through the basement membrane layer to the breast stroma.Citation1–Citation3 In several cases, it is known to be preliminary to IBC, and it has been suggested that the proliferation of DCIS is dependent on the presence of estrogen. Once the DCIS becomes cancerous, it is known as ductal carcinoma, which is a form of hormone-dependent breast cancer.Citation1–Citation3 The T-47D cell line used in our experiments was isolated from infiltrating ductal carcinoma of the breast and is positive for both estrogen receptor (ER) and progesterone receptor (PR).
Curcumin (CUR), also known as diferuloylmethane, has the chemical formula 1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione. It is an active polyphenolic compound that is isolated from the spice turmeric (Curcuma longa). Its chemical structure includes α,β unsaturated β-diketones, along with hydrophobic phenyl groups that are vital to its binding activity with molecular targets. While CUR is commonly used as a spice, food additive, and dietary pigment, it has captivated attention for having antibacterial, antiviral, anti-inflammatory, and anticancer properties.Citation4–Citation6 It is also considered to have valuable potential with regards to the targeting of various cancers.Citation6 Hallman et al
CUR also exhibits anticancer effects in cancer stem-like cells (CSCs). It has been shown to considerably diminish microtentacles of the plasma membrane of CSCs, thereby preventing reattachment of the cells at distant tissues.Citation7 It has also been demonstrated that CUR restores expression of the tumor suppressor gene E-cadherin in breast CSCs.Citation8
A significant number of other reports have indicated that the anticancer effects of CUR are due to the regulation of multiple molecular targets involved in tumorigenesis, angiogenesis, metastasis, invasion, proliferation, and apoptosis. It is also known to significantly reduce the harmful reactions of reactive oxygen species.Citation9–Citation23 These anticancer effects are also demonstrated when CUR is present in different analogs, or when combined with other compounds.Citation24–Citation32 CUR-loaded nanoparticles may also have potential applications in breast cancer therapy and other various cancers.Citation33,Citation34
Numerous studies have demonstrated that CUR exhibits multiple effects in the suppression of human breast cancer cells in vitro, leading to the incorporation of CUR in ongoing clinical trials.Citation35–Citation39 It has been reported that CUR hinders the growth of T-47D breast cancer cells in a concentration-dependent manner.Citation40
CUR-induced apoptosis has been associated with stimulation of the p53 pathway.Citation41 Consistent with our findings that CUR decreases the regulation of p53 in T-47D breast cancer cells, it has been demonstrated that CUR impairs the expression and function of p53 in colon cancer cells through inhibition of its gene transcription.Citation42 Literature has shown that the p53 gene is mutated in the T-47D breast cancer cell line. However, the actual quantity of mutated versus wild-type p53 remains unknown in the T-47D cells.Citation43–Citation45 Studies have also shown that in estrogen receptor (ER)-positive breast cancer cells, the antiproliferative effects of CUR are dependent on the presence of estrogen.Citation11,Citation46 Reports indicate that breast cancer cells treated with CUR show a significant downregulation in the expression of ERα compared with the control cells.Citation40 Previous studies, including studies from our laboratory, have shown that treatment with estrogen in T-47D breast cancer cells causes increased cell proliferation and upregulation of the tumor suppressor protein, p53.Citation47–Citation49 In this study, we have analyzed the effects of CUR on the expression of ERα and p53 in the presence of estrogen, bisphenol-a (BPA), and anti-estrogens in T-47D breast cancer cells. Our studies validate CUR’s potential as a therapeutic agent for breast cancer.
Methods and materials
Cell culture and treatment with ligands
T-47D human breast cancer cells (ATCC® HTB-133™; American Type Culture Collection, Rockville, MD, USA) were routinely cultured in RPMI-1640 media (Hyclone, Logan, UT, USA) containing 2 mM l-glutamine, 25 mM HEPES, 24 mM sodium bicarbonate, 0.5% 100× non-essential amino acids, 100 U/mL penicillin, 0.1 mg/mL streptomycin, 0.25 mg/mL amphotericin B (Hyclone), and 0.14 IU/mL insulin (Sigma, St Louis, MO, USA) supplemented with 10% (v/v) fetal bovine serum (FBS; Hyclone). In all studies, cells were plated in 10% FBS media and incubated at 37°C in the presence of 5% CO2. After the cells were allowed to attach and grow for 48 hours, they were cultured in 5% FBS stripped with dextran-coated charcoal (DCC-FBS), which ensures steroid-free treatment conditions, for a total of 6 days with fresh medium added every 48 hours. On the sixth day, semi-confluent cells were treated with various ligands for 3 or 24 hours. Stock solutions of the ligand used for cell treatment were prepared in DMSO to a 1000-fold higher concentration than the final concentration to be extracted.
Extraction of cells
Following the removal of media by aspiration, cells were washed with ice-cold Hanks balanced salt solution and scraped into 200 µL of RIPA lysis buffer. The cells were thawed on ice. To prepare a high-speed supernatant (HSS), the cells were centrifuged for 15 minutes at 15,000×g at 4°C. The HSS for each sample was stored at −80°C until further use.
SDS–PAGE and Western blot analysis
Extracted protein supernatants were quantified using the Bradford method to standardize the amount of protein to be loaded into each lane. After the supernatants for each sample were denatured for 3 minutes at 85°C, 30 µg aliquots of total protein were loaded into each lane on a 7.5% polyacrylamide gel. The gels were subjected to electrophoresis to perform protein band separation. Proteins were wet transferred onto an Immobilon-P polyvinylidene fluoride (PVDF) membrane (Millipore, Bedford, MA,USA) in a tris-glycine buffer system (0.025% SDS and 15% methanol) contained in a Bio-Rad (Hercules, CA, USA) trans blot cell set to 110 V for 45 minutes. Membranes were blocked for 1 hour in 5% (w/v) non-fat dry milk (NFDM) in TBS-T (20 mM Tris–HCl, 140 mM NaCl, pH 7.4, 0.1% [v/v] Tween 20) and incubated with primary antibody anti-ERα clone F-10 and anti-p53 (Santa Cruz) diluted 1:500 in 5% (w/v) NFDM in TBS-T for 2 hours. Actin bands were probed by anti-actin (monoclonal antibody clone C4) (Millipore). ERα and p53 levels were normalized to protein levels of the evolutionarily conserved actin protein according to the manufacturer’s protocol. After washing with TBS-T (3×10 minutes) and incubation with horse radish peroxidase (HRP)-conjugated goat antimouse IgG2a secondary antibody (Santa Cruz) diluted 1:1000 in 5% (w/v) NFDM in TBS-T for 1 hour, blots were developed using Amersham ECL Prime (GE Healthcare Biosciences, Piscataway, NJ, USA). ERα and p53-specific bands were visualized using the Bio-Rad Chemi-Doc XRS + System. After immunoblotting, the PVDF membranes were stained with Coomassie blue to ensure the correct normalization against total protein levels and full transfer of protein. The protein band density on each membrane was quantified using the Image Studio Lite program version 3.1 (LI-COR Biosciences, Lincoln, NE, USA).
Cell viability and apoptosis assays
All growth studies were conducted in 12-well culture plates. To ensure active, non-confluent cell populations during treatment duration, 12-well culture plates were initially seeded with 3.0×104 cells per well in 1 mL culture medium containing 10% FBS. Studies were limited to 7-day total duration to correlate with the results of Western blot analyses. On the seventh day, the cells were trypsinized and removed from individual wells of the culture plate. Cells were stained with propidium iodide (PI) for viability assays and fluorescein isothiocyanate-conjugated Annexin V for apoptosis assays per manufacturer’s instruction (Cellometer Inc., Lawrence, MA, USA). The cells underwent imaging cytometry by the Cellometer Vision CBA and FCS Express-6 software. The fluorescent threshold in the software was set to 0% to measure total fluorescence of each counted cell from the captured images. The measured fluorescence intensities were exported to Microsoft Excel and converted to FCS file for analysis in the De Novo Software.Citation55
Immunofluorescence and confocal microscopy
T-47D cells were plated on cover slips in 12-well plates (30,000 cells/well) and cultured for 48 hours in a 10% FBS medium. The medium was then changed to 5% DCC-FBS, and fresh medium was added at 2-day intervals. The cells were cultured in this medium for a total of 4 days. On the fourth day, the ligands were suspended in 5% DCC-FBS media, and semi-confluent cells were treated for 24 hours.
Cellular localization of p53 by immunocytochemistry
The cells were fixed on cover slips for 10 minutes with 1% formalin in phosphate-buffered saline (PBS), permeabilized with ice-cold acetone and methanol (50:50), and washed three times with PBS. Staining procedures were performed in a humidified chamber at 23°C. Cells were incubated in 10% goat serum (Sigma) to suppress nonspecific binding of IgG, followed by 3 hours incubation with 1:150 dilution of anti-p53 (F-10) monoclonal antibody. After washing with PBS, cells were incubated for 3 hours with 1:200 dilution of antimouse IgG conjugated with Cy3 (Jackson Immuno Research Laboratories, West Grove, PA, USA). Cover slips were washed in PBS and incubated for 2 minutes in 1 µg/mL DAPI dissolved in PBS. Cells were washed three times in PBS, mounted with Fluoromount-G (Electron Microscopy Sciences, Hatfield, PA, USA) and stored in the dark at 4°C.
Structural analysis by DIC
DIC images were acquired by using the DIC objectives with Nikon Digital Eclipse C1 Plus confocal microscope. The distribution of three-dimensional fluorescent structures was analyzed using a Nikon Digital Eclipse C1 Plus confocal microscope. NIS elements software (Nikon Instruments, Melville, NY, USA) was used for noise reduction and three-dimensional reconstruction of the images.
Statistical analyses
The results are expressed as mean±SEM. Statistical significance was determined by Kruskal–Wallis test followed by post-hoc analysis using Mann–Whitney U test. The p values were adjusted for multiple testing corrections using the false discovery rate. Differences are considered significant at p<0.05. Statistical analyses were carried out using SPSS for Windows version 11.5 (SPSS Inc., Chicago, IL, USA).
Results
Effects of CUR on cellular viability and the regulation of ERα and p53 levels: concentration dependency
demonstrates the concentration-dependent effects of CUR on cell viability and the expression of ERα and p53 protein levels. To examine the effects on cell viability, the T-47D cells were treated with 5–100 µM CUR for 6 days. Cellular viability was then determined by propidium iodide (PI) staining and image cytometry. As shown in , 20–100 µM CUR significantly decreases T-47D cell viability as compared with the control. CUR at lower concentrations (5–10 µM) did not display a significant effect on the cell viability compared with the control.
Figure 1 Concentration-dependent effects of CUR: curcumin decreases T-47D cell viability and downregulates p53 and ERα protein expression.
Abbreviations: CUR, curcumin; DMSO, dimethyl sulfoxide; ERα, estrogen receptor; PI, propidium iodide.
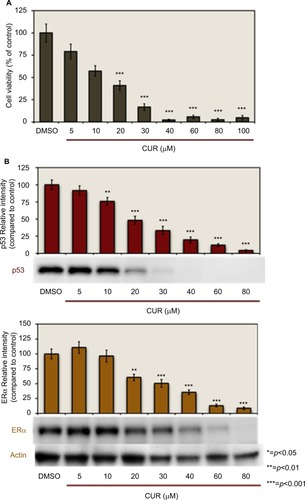
T-47D cells were treated with 5–80 µM CUR for a duration of 24 hours. The cells were then subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) and Western blot analysis to examine the concentration-dependent effects of CUR on the level of ERα and p53 (). The relative densities of each of the protein bands were compared with the control. Upon examination of each of the densities, shows that 20–80 µM CUR significantly downregulates both ERα and p53 protein levels.
Differential interference contrast microscopy of 60 μM CUR cytotoxicity in T-47D cells
Differential interference contrast (DIC) images were taken of the T-47D cells to see the contour of the cells (). T-47D cells were treated with 60 µM CUR for 6 days, and images were acquired as described in Methods and Materials section. The merged images in indicate that 60 µM CUR depleted the T-47D cells.
Figure 2 Effects induced by 60 μM CUR.
Abbreviations: CUR, curcumin; ERα, estrogen receptor; DIC, Direct Image Contrast; DAPI, diamidino-2-phenylindole; DMSO, dimethyl sulfoxide.
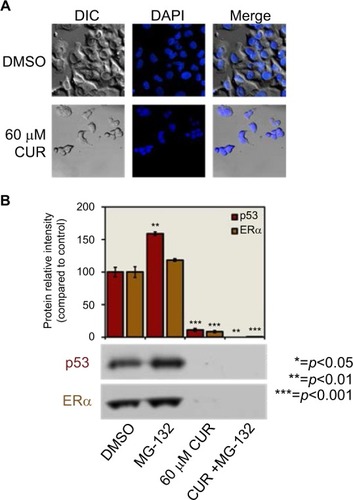
Effects induced by 60 μM CUR in combination with MG-132
MG-132 is a proteasomal inhibitor that is known to prevent the degradation of short-lived proteins. Therefore, it is a valuable research tool for examining the cellular degradation through the proteasome pathway.Citation50,Citation51 To determine whether CUR is an inhibitor of proteasomal degradation, 60 µM CUR was analyzed to verify if MG-132 reversed its effects on the downregulation of protein levels ().
T-47D cells were pretreated with 10 µM MG-132 for 30 minutes, followed by a 3-hour treatment with 60 µM CUR. Protein extracts were subjected to SDS–PAGE and Western blot analysis, and the relative densities of each band were compared with control (dimethyl sulfoxide [DMSO]), as described in Methods and Materials section. As indicated in , MG-132 did not reverse the effects of 60 µM CUR, which suggests that the downregulation of protein levels caused by CUR is not due to mechanistic causes but rather due to its cytotoxicity to the cells.
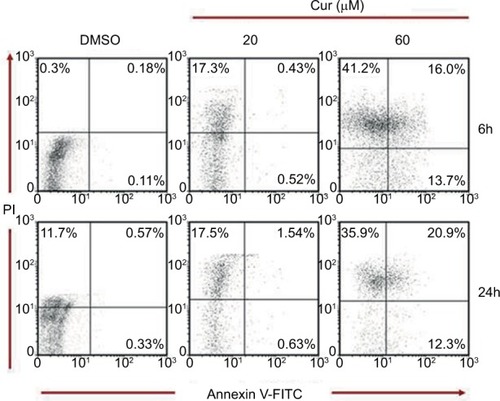
Effects of 20 and 60 μM CUR on the apoptosis of T-47D cells
As illustrated in , scatter plots following Cellometer Vision CBA and FCS Express-6 Analysis show an increased trend in the apoptotic/necrotic cell population following incubation with 60 µM of CUR treatment for 6 hours compared with the negative controls. This trend explains a shift from the apoptotic (annexin V-positive only) to necrotic (annexin V and PI positive) cell population. This trend seems to increase when the cells were treated with 60 µM of CUR for 24 hours. However, treating the cells with 20 µM of CUR has shown a lesser trend of apoptotic/necrotic cell population in 6 and 24 hours.Citation52–Citation55
Figure 3 Apoptosis assay: effects of 20 and 60 μM curcumin on T-47D cells.
Abbreviations: CUR, curcumin; DMSO, dimethyl sulfoxide; PI, propidium iodide.
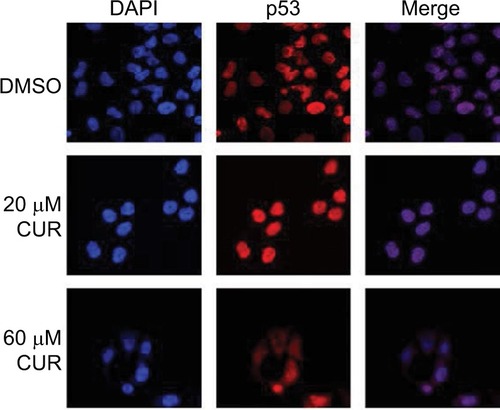
Effects of CUR on the immunolocalization of p53 in T-47D cells
Along with Western blot studies ( and ), image analysis was performed by confocal microscopy involving immunofluorescence to determine the cytolocalization of cellular p53. 4′,6-Diamidino-2-phenylindole (DAPI) is used to stain the nuclei of the cells so that the nuclei will be displayed in the image analysis (). As shown in , p53 is localized exclusively in the nuclei of the T-47D cells. The control group (DMSO) represents cells grown in a charcoal-treated serum that displays immunofluorescence localized in the nuclei of the T-47D cells. Treatments of 20 µM and 60 µM CUR did not affect the cytolocalization of cellular p53, as the image analysis shows p53 to still be localized in the nucleus (). The images of the cells stained for the nuclei and stained for p53 were merged to further demonstrate that p53 is localized exclusively in the nuclei.
Figure 4 Effects of CUR on the immunolocalization of p53 in T-47D cells.
Abbreviations: CUR, curcumin; DAPI, diamidino-2-phenylindole; DMSO, dimethyl sulfoxide.
Effects of CUR in combination with ER agonists and antagonists on cellular viability and expression of ERα and p53
displays the effects of ER agonists and antagonists in combination with CUR. T-47D cells were treated with 20 and 60 µM CUR, 10 nM E2, 600 nM BPA, 1 µM ICI, and 1 µM tamoxifen (TAM) either alone or in combination. As mentioned in Methods and Materials section, the cells studied for cell viability were treated for 6 hours, and the cells being studied for p53 and ERα expression by Western blot analysis were treated for 24 hours. The data show that even with the addition of the anti-estrogens ICI and TAM, the estrogen E2, and the ER agonist BPA, the effect of CUR on T-47D cell viability could not be reversed ().
Figure 5 Curcumin-induced effects are not mediated by ERα.
Abbreviations: DMSO, dimethyl sulfoxide; TAM, tamoxifen; CUR, curcumin; BPA, Bisphenol A.
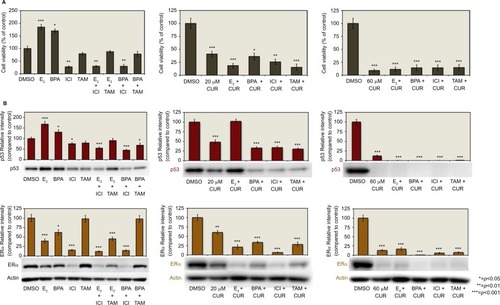
indicates that when the cells were treated with 20 and 60 µM CUR, the level of p53 protein expression was significantly downregulated compared with the control. The addition of E2, ICI, and TAM to the treatment of 20 µM CUR did not alter the significant downregulation of p53, but the addition of BPA with 20 µM CUR had no effect. The additions of E2, BPA, ICI, and TAM to the 60 µM treatment of CUR still resulted in the downregulation of p53.
As shown in , the expression of ERα is also significantly downregulated compared with the control by 20 and 60 µM CUR. It is also demonstrated that the addition of E2, BPA, ICI, and TAM to both 20 and 60 µM CUR did not reverse the downregulation of ERα expression ().
Discussion
Our studies demonstrate that 20–80 µM CUR significantly downregulates both p53 and ERα protein levels with a concomitant decrease in T-47D cell viability. Other studies involving different breast cancer cell lines treated with CUR, such as BT-483 and MDA-MB-23, have shown a significant decrease in cell proliferation. For example, 48-hour treatments of 5 µL/mL CUR inhibited proliferation of BT-483 cells by 60%–70% and MDA-MB-23 cells by 50%–60%.Citation56 CUR has been found to be highly cytotoxic to some breast cancer cell lines. This is consistent with our findings that the proteasomal inhibitor MG-132 did not reverse the effects of 60 µM CUR on the downregulation of ERα and p53 in T-47D cells.Citation57 In our studies, the presence of anti-estrogens and ER agonists, including BPA, did not reverse the effects of CUR on T-47D cell viability, and this reduction could not be explained by an induction of apoptosis under our experimental conditions. In addition, when CUR is used in combination with ERα agonists and antagonists, the alterations in p53 protein expression do not correlate with alterations in cell viability.
It has been shown that CUR inhibits the phosphorylation of p53, its ability to bind to DNA, and the transactivation of genes that correlate with its function as a tumor suppressor.Citation42 However, our experiments suggest that the CUR-induced effects on the T-47D cells are not due to a mechanism involving p53 and appear to involve a mechanism that includes proteins other than p53 and ERα. CUR has also been found to inhibit the expression of the leptin gene in T-47D breast cancer cells, and this inhibition was positively associated with a decrease in the expression of ERα.Citation40 Experiments involving the multiple effects of CUR on human breast cancer cells have shown that the antiproliferative effects of CUR are estrogen-dependent in ER-positive MCF-7 breast cancer cells.Citation46 These findings suggest that CUR acts through estrogen receptors to demonstrate its anticancer effects in hormone-dependent breast cancer cells. However, our results indicate that CUR-induced effects are not mediated via ERα. These results are supported by the findings of CUR having antiproliferative effects on the ER-negative MDA-MB-231 breast cancer cell line, and these effects do not depend on the presence of estrogen.Citation46 When MDA-MB-231 cells are treated with a combination of CUR and epigallocatechin gallate, another polyphenolic compound, antiproliferative effects are still present, further suggesting that the effects of CUR do not involve ERα.Citation26
It remains clear that 20–80 µM CUR affects p53 and ERα protein expression on a molecular level. Whether these two proteins, known to contribute to the formation and progression of hormone-dependent breast cancer, are regulated on a transcriptional or translational level remains unclear. Although several studies have indicated estrogen-dependent effects of CUR, our results demonstrate effects that are not dependent on the presence of estrogen. Therefore, the effectiveness of CUR on hormone-dependent breast cancer cells needs to be re-evaluated. CUR has been studied as a chemopreventive agent in numerous clinical trials, and its effects should be further examined in vivo as well as in vitro.Citation35–Citation39 Further studies are warranted as our results support the potential and suitability of CUR as a treatment modality in hormone-dependent breast cancer.
Disclosure
The authors report no conflicts of interest in this work.
Acknowledgments
Amy E Siebert is currently working in Department of Biological Sciences at Oakland University.
References
- WiechmannLKuererHMThe molecular journey from ductal carcinoma in situ to invasive breast cancerCancer2008112102130214218383519
- VaidyaYVaidyaPVaidyaTDuctal carcinoma in situ of the breastIndian J Surg201577214114626139969
- CowellCFWeigeltBSakrRAProgression from ductal carcinoma in situ to invasive breast cancer: revisitedMol Oncol20137585986923890733
- ChoudhuriTPalSAgwarwalMLDasTSaGCurcumin induces apoptosis in human breast cancer cells through p53-dependent Bax inductionFEBS Lett20025121–333434011852106
- MudduluruGGeorge-WilliamJNMuppalaSCurcumin regulates miR-21 expression and inhibits invasion and metastasis in colorectal cancerBiosci Rep201131318519720815812
- DeguchiACurcumin targets in inflammation and cancerEndocr Metab Immune Disord Drug Targets2015152889625772169
- CharpentierMSWhippleRAVitoloMICurcumin targets breast cancer stem–like cells with microtentacles that persist in mammospheres and promote reattachmentCancer Res20147441250126024371229
- MukherjeeSMazumdarMChakrabortySCurcumin inhibits breast cancer stem cell migration by amplifying the E-cadherin/β-catenin negative feedback loopStem Cell Res Ther20145511625315241
- KunnumakkaraABAnandPAggarwalBBCurcumin inhibits proliferation, invasion, angiogenesis and metastasis of different cancers through interaction with multiple cell signaling proteinsCancer Lett2008269219922518479807
- MukhopadhyayACurcumin-induced suppression of cell proliferation correlates with down-regulation of cyclin D1 expression and CDK4-mediated retinoblastoma protein phosphorylationOncogene200221578852886112483537
- VermaSPGoldinBRLinPSThe inhibition of the estrogenic effects of pesticides and environmental chemicals by curcumin and isoflavonoidsEnviron Health Perspect199810612807812
- SarkarFHLiYWangZPadhyeSLesson learned from nature for the development of novel anti-cancer agents: implication of isoflavone, curcumin, and their synthetic analogsCurr Pharm Des201016161801181220345353
- ZhouHBeeversCSHuangSThe targets of curcuminCurr Drug Targets201112333234720955148
- PrasadCPRathGMathurSBhatnagarDRalhanRPotent growth suppressive activity of curcumin in human breast cancer cells: Modulation of Wnt/β-catenin signalingChem Biol Interact2009181226327119573523
- ZhanYChenYLiuRZhangHZhangYPotentiation of paclitaxel activity by curcumin in human breast cancer cell by modulating apoptosis and inhibiting EGFR signalingArch Pharm Res20143781086109524318305
- PrasadCPRathGMathurSBhatnagarDRalhanRExpression analysis of maspin in invasive ductal carcinoma of breast and modulation of its expression by curcumin in breast cancer cell linesChem Biol Interact2010183345546119944674
- ChenBZhangYWangYRaoJJiangXXuZCurcumin inhibits proliferation of breast cancer cells through Nrf2-mediated down-regulation of Fen1 expressionJ Steroid Biochem Mol Biol2014143111824486718
- KimBKimHS2JungEJCurcumin induces ER stress-mediated apoptosis through selective generation of reactive oxygen species in cervical cancer cellsMol Carcinog201655591892825980682
- GuanFDingYZhangYZhouYLiMWangCCurcumin suppresses proliferation and migration of MDA-MB-231 breast cancer cells through autophagy-dependent akt degradationPLoS One2016111e014655326752181
- KimSRParkHJBaeYHCurcumin down-regulates visfatin expression and inhibits breast cancer cell invasionEndocrinology2012153255456322186408
- KimHIHuangHCheepalaSHuangSChungJCurcumin inhibition of integrin (α6β4)-dependent breast cancer cell motility and invasionCancer Prev Res200815385391
- GuoJLiWShiHSynergistic effects of curcumin with emodin against the proliferation and invasion of breast cancer cells through upregulation of miR-34aMol Cell Biochem2013382110311123771315
- SunKDuanXCaiHCurcumin inhibits LPA-induced invasion by attenuating RhoA/ROCK/MMPs pathway in MCF7 breast cancer cellsClin Exp Med2016161374725596714
- PatelPBThakkarVRPatelJSCellular effect of curcumin and citral combination on breast cancer cells: induction of apoptosis and cell cycle arrestJ Breast Cancer201518322523426472972
- De GasperiMCavazosDdeGraffenriedLCurcumin modulates tamoxifen response in resistant breast cancer cellsCancer Res20096924 Suppl3098
- Somers-EdgarTJScandlynMJStuartECLe NedelecMJValentineSPRosengrenRJThe combination of epigallocatechin gallate and curcumin suppresses ERα-breast cancer cell growth in vitro and in vivoInt J Cancer200812291966197118098290
- SunALuYJHuHShojiMLiottaDCSnyderJPCurcumin analog cytotoxicity against breast cancer cells: exploitation of a redox-dependent mechanismBioorg Med Chem Lett200919236627663119854644
- KakaralaMBrennerDEKorkayaHTargeting breast stem cells with the cancer preventive compounds curcumin and piperineBreast Cancer Res Treat2010122377778519898931
- Al-HujailyEMMohamedAGAl-SharifIPAC, a novel curcumin analogue, has anti-breast cancer properties with higher efficiency on ER-negative cellsBreast Cancer Res Treat201112819710720680677
- MohankumarKPajaniradjeSSridharanSMechanism of apoptotic induction in human breast cancer cell, MCF-7, by an analog of curcumin in comparison with curcumin – An in vitro and in silico approachChemico-Biol Interact20142105163
- WangSChenRZhongZEpigallocatechin-3-gallate potentiates the effect of curcumin in inducing growth inhibition and apoptosis of resistant breast cancer cellsAm J Chin Med20144251279130025242081
- MohankumarKSridharanSPajaniradjeSBDMC-A an analog of curcumin, inhibits markers of invasion, angiogenesis, and metastasis in breast cancer cells via NF-κB pathway-a comparative study with curcuminBiomed Pharmacother20157417818626349982
- YallapuMMOthmanSFCurtisETCurcumin-loaded magnetic nanoparticles for breast cancer therapeutics and imaging applicationsInt J Nanomed2012717611779
- AnithaAMayaSDeepaNEfficient water soluble O-carboxymethyl chitosan nanocarrier for the delivery of curcumin to cancer cellsCarbohydrate Polymers2011832452461
- KanaiMOtsukaYOtsukaKA phase I study investigating the safety and pharmacokinetics of highly bioavailable curcumin (Theracurmin®) in cancer patientsCancer Chemother Pharmacol20137161521153023543271
- ShehzadAWahidFLeeYSCurcumin in cancer chemoprevention: molecular targets, pharmacokinetics, bioavailability, and clinical trialsArch Pharm20103439489499
- DhillonNAggarwalBBNewmanRAPhase II trial of curcumin in patients with advanced pancreatic cancerClin Cancer Res200814144491449918628464
- CarrollREBenyaRVTurgeonDKPhase II a clinical trial of curcumin for the prevention of colorectal neoplasiaCancer Prev Res201143354364
- SharmaRAEudenSAPlattonSLPhase I clinical trial of oral curcumin: biomarkers of systemic activity and complianceClin Cancer Res200410206847685415501961
- Nejati-KoshkiKAkbarzadehAPourhassan-MoghaddamMCurcumin inhibits leptin gene expression and secretion in breast cancer cells by estrogen receptorsCancer Cell Int2014146625866478
- LiontasAYegerHCurcumin and resveratrol induce apoptosis and nuclear translocation and activation of p53 in human neuroblastomaAnticancer Res2004242B98799815161054
- MoosPJEdesKMullallyJEFitzpatrickFACurcumin impairs tumor suppressor p53 function in colon cancer cellsCarcinogenesis20042591611161715090465
- BerglindHPawitanYKatoSIshiokaCSoussiTAnalysis of p53 mutation status in human cancer cell lines: a paradigm for cell line cross-contaminationCancer Biol Ther20087569970818277095
- LimLYVidnovicNEllisenLWLeongCOMutant p53 mediates survival of breast cancer cellsBr J Cancer200910191606161219773755
- GartelALFelicianoCTynerTLA new method for determining the status of p53 in tumor cell lines of different originOncol Res2003136–1040540812725531
- ShaoZMShenZZLiuCHCurcumin exerts multiple suppressive effects on human breast carcinoma cellsInt J Cancer200298223424011857414
- SiebertAESanchezALDindaSMoudgilVKEffects of estrogen metabolite 2-methoxyestradiol on tumor suppressor protein p53 and proliferation of breast cancer cellsSyst Biol Reprod Med201157627928722077725
- HurdCKhattreeNAlbanPHormonal regulation of the p53 tumor suppressor protein in T47D human breast carcinoma cell lineJ Biol Chem19952704828507285107499360
- DindaSSanchezAMoudgilVKEffects of LY117018 (a SERM analog of raloxifene) on tumor suppressor proteins and proliferation of breast cancer cellsHorm Mol Biol Clin Investig201021211217
- LaïosIJournéFNonclercqDRole of the proteasome in the regulation of estrogen receptor α turnover and function in MCF-7 breast carcinoma cellsJ Steroid Biochem Mol Biol200594434735915857754
- YehWLShiodaKCoserKRRivizzignoDMcSweeneyKRShiodaTFulvestrant-induced cell death and proteasomal degradation of estrogen receptor? Protein in MCF-7 cells require the CSK c-Src tyrosine kinasePLoS One201384e6088923593342
- JiZTangQZhangJYangYLiuYPanYOridonin-induced apoptosis in SW620 human colorectal adenocarcinoma cellsOncol Lett2011261303130722848306
- BergerEAMcClellanSAVistisenKSHazlettLDHIF-1α is essential for effective PMN bacterial killing, antimicrobial peptide production and apoptosis in Pseudomonas aeruginosa keratitisPLoS Pathog201397e100345723874197
- YedjouCGSaeedMAHossainMADorseyWYuHTchounwouPBBasic apoptotic and necrotic cell death in human liver carcinoma (HepG2) cells induced by synthetic azamacrocycleEnviron Toxicol201429660561122644747
- ChanLLLaiNWangESmithTYangXLinBA rapid detection method for apoptosis and necrosis measurement using the cellometer imaging cytometryApoptosis201116121295130321910006
- LiuQLooWTSzeSCTongYCurcumin inhibits cell proliferation of MDA-MB-231 and BT-483 breast cancer cells mediated by down-regulation of NFκB, cyclinD and MMP-1 transcriptionPhytomedicine2009161091692219524420
- LiuDChenZThe effect of curcumin on breast cancer cellsJ Breast Cancer201316213313723843843