
Abstract
GATA3, a member of the GATA zinc finger transcription factor family, has been widely investigated for its role in cancer. Although a recent report has found that GATA3 is downregulated in gastric cancer (GC), the detailed mechanism of GATA3 in GC is still unknown. Here, we investigated whether GATA3 was downregulated in GC patients’ tissue samples and cell lines using quantitative real time polymerase chain reaction and Western blotting. In addition, we conducted several functional experiments to investigate the effect of GATA3 in GC, including cell proliferation, metastasis and epithelial–mesenchymal transition (EMT). The results showed that GATA3 was downregulated in GC tissue samples and cells. Moreover, the expression of GATA3 was associated with tumor size, stage and metastasis. Restoration of GATA3 levels suppressed GC cell proliferation, migration and invasion. Furthermore, chromatin immunoprecipitation and luciferase reporter assay also revealed that GATA3 transcriptionally regulated ZEB1, thereby suppressing EMT. All these findings suggest that GATA3 serves as an oncogene in GC development.
Introduction
Gastric cancer (GC) has become the fourth most common malignant cancer in the world.Citation1 Almost one million new cases are diagnosed each year.Citation1 Although the treatments, including chemotherapy, radiation therapy and surgery, can increase the survival rate, approximately 60% of patients succumb to GC.Citation2 To improve the prognosis of GC, it is necessary to further decipher the detailed molecular mechanisms of GC progression and development.Citation3–Citation5
GATA zinc finger transcription factor family consists of six members, including GATA-binding protein 1–6. GATA family protein has been found to bind to the specific DNA sequence (A/T)GATA(A/G) and plays an essential role in development, proliferation and differentiation of several types of tissues and cells,Citation6 including the kidney,Citation7 thymocytes,Citation8,Citation9 T-lymphocytes,Citation10 the sympathetic nervous system,Citation11 luminal glandular epithelial cells of the mammary gland,Citation12–Citation14 hair follicles of the skinCitation15 and adipose tissue.Citation16 The significance of this transcription factor has been discussed in recent works, and its role as a tumor suppressor or an oncogene has been debated in many papers.Citation17,Citation18 A low expression of GATA3 has also been found to be associated with a poor prognosis of breast cancer.Citation19,Citation20 Moreover, a recent report showed that GATA3 was downregulated in GC and correlated with poor prognosis in primary gastric adenocarcinoma,Citation21 but the detailed mechanism of GATA3 in GC development is still unknown.
In this work, we found that GATA3 was downregulated in GC tissue samples and cells. Interestingly, the expression of GATA3 was negatively associated with tumor size, stage and metastasis, but there were no correlations between the expression of GATA3 and gender, age and alcohol consumption. The restoration of GATA3 suppressed cell proliferation, migration and invasion. Moreover, GATA3, a transcription factor, transcriptionally regulated ZEB1, thereby inhibiting epithelial–mesenchymal transition (EMT). Therefore, in this study, we proved that GATA3 serves as an oncogene in GC development and acts as a novel therapeutic target.
Materials and methods
Cell culture and collection of tissue samples
Human gastric epithelial cells GES-1 (Bogu Biotech, Shanghai, People’s Republic of China) and human GC cell lines CTC-141 and MKN45 (ATCC, Manassas, VA, USA) were grown in Dulbecco’s Modified Eagle’s Medium (DMEM; HyClone, Logan, UT, USA) supplemented with 100 U/ml penicillin, 100 μg/ml streptomycin and 10% fetal bovine serum (FBS) at 37°C with 5% CO2.
The 83 pairs of GC tissue samples and adjacent normal tissue samples were collected from Zhoupu Hospital during 2013–2016. Written informed consent was obtained from each patient, and the experiments involving human tissue samples were approved by the Clinical Research Ethics Committee of Zhoupu Hospital.
Quantitative real time polymerase chain reaction (qRT-PCR)
RNeasy Mini kit (Thermo Fisher Scientific, Waltham, MA, USA) was used to extract total RNA from tissue samples and cells according to our protocol. Subsequently, cDNA was synthesized using EasyScript Reverse Transcriptase kit (TransGen Biotech, Beijing, People’s Republic of China). qRT-PCR was set up using the TransStart Tip Green qPCR SuperMix (TransGen Biotech) and the targeting gene primers (GATA-3: forward: 5′-GCTTCACAATATTAACAGACCC-3′, reverse: 5′-TTAAACGAGCTGTTCTTGGG-3′; E-cadherin: forward: 5′-ATTTCCCAACTCCTCTCCTG-3′, reverse: 5′-GTTGGATTTGATCTGAACCAGG-3′; α-catenin: forward: 5′-AGGTTATTAATGCTGCACTG-3′, reverse: 5-AATGTGATTCTCTGAGACAGC-3′; N-cadherin: forward: 5′-TCATTAATGAGGGCCTTAAAGC-3′, reverse: 5′-GTTCAGGTAATCATAGTCCTGCT-3′; vimentin: forward: 5′-AAATCCAAGTTTGCTGACCTC-3′, reverse: 5′-GGTCTTGGTATTCACGAAGG-3′; ZEB1: forward: 5′-GGTCTTGGTATTCACGAAGG-3′, reverse 5′-CTTGTCTTTCATCCTGATTTCC-3′). The Ct values were normalized to GAPDH (forward: 5′-TCAAGATCATCAGCAATGCC-3′, reverse: 5′-ATGGACTGTGGTCATGAGTC-3′). Each experiment was performed at least three times.
Western blotting
Tissue samples and cells were collected and washed three times with cold PBS and then homogenized in RIPA lysis buffer (1% NP-40, 0.5% deoxycholic acid, 50 mM Tris-HCl [pH 8.0], 0.1% sodium dodecyl sulfate (SDS), 150 mM NaCl) containing a cocktail of protease inhibitors (Sigma-Aldrich, St Louis, MO, USA). Following sonication and centrifugation (13,000× g, 15 min, 4°C), the supernatant was collected. The BCA protein assay kit (Pierce, Waltham, MA, USA) was used to determine the protein concentration according to our protocol. Samples were denatured in loading buffer, separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes. Anti-GATA-3 (Abcam, Cambridge, UK; diluted 1:200), anti-ZEB1 (Abcam; diluted 1:200), EMT kit (CST, Danvers, MA, USA; diluted 1:1000) and β-actin (Sigma-Aldrich; diluted 1:5000) were incubated at 4°C overnight followed by incubation with HRP-conjugated secondary antibody (Abcam; diluted 1:5000). Specific blots were visualized using the ECL kit (Thermo Fisher Scientific).
Transfection
Cells were transfected with GATA3 siRNA using Lipofectamine RNAi reagent (Thermo Fisher Scientific). The sequences of GATA3 siRNA were: siRNA-1: 5′-GGCUCUACUACAAGCUUCACAAUAU-3′ and siRNA-2: 5′-CCACACUCUGGAGGAGGAAUGCCAA-3′.Citation22
Cell Counting Kit-8 (CCK-8) analysis
CCK-8 analysis was used to detect the effect of GATA3 on cell proliferation. GATA3 was overexpressed or knocked down in CTC-141 and MKN45 cells. After transcription for 48 h, a total of 3,000 CTC-141 and MKN45 cells were placed in 96-well plates with 200 μl DMEM. Every 24 h, 20 μl CCK-8 regent (Beyotime, Haimen, People’s Republic of China) was added, and cells were incubated at 37°C for 90 min. The absorbance at OD 450 nm revealed the number of surviving cells. Each experiment was performed at least three times.
Colony formation assay
After transfection for 48 h, approximately 5,000 cells were placed in six-well plates and incubated with serum-free DMEM for 12 days, after which cells were washed with PBS three times and fixed with methanol. After staining with 0.1% crystal violet, the number of colonies was counted under a light microscope. Each experiment was performed at least three times.
Cell cycle analysis
CTC-141 and MKN45 cells were transfected with vector, GATA3, scramble siRNA (SCR) or siGATA3,, respectively, for 48 h. Subsequently, cells were collected and fixed with ice-cold 70% ethanol at −20°C overnight. After washing the cells with ice-cold PBS three times, 1 mg/ml RNase A (Sigma-Aldrich) was added, and cells were incubated at 37°C for 60 min. Finally, cells were stained with propidium iodide (Sigma-Aldrich) for 15 min in the dark, and DNA contents were determined using FACScan flow cytometry. Each experiment was performed at least three times.
Transwell migration and invasion assay
Transwell migration and invasion analysis was used to detect the effect of GATA3 on the migratory and invasive behavior of GC cells. Briefly, for Transwell migration and invasion assay, some Transwell chambers (8 μm pore size; EMD Millipore, Billerica, MA, USA) were coated with 100 μl Matrigel (BD, Franklin Lakes, NJ, USA), and some were left uncoated. A total of 4×104 CTC-141 cells were placed on top of Transwell chambers with serum-free DMEM; the lower chambers were filled with 500 μl DMEM containing 10% FBS. Subsequently, cells were incubated at 37°C with 5% CO2 for 18 h, fixed with methanol and stained with 0.1% violet crystal. Cells that remained on the top of the filter were removed using a cotton swab. Finally, the number of migrated and invaded cells was counted under a light microscope. Each experiment was performed at least three times.
Chromatin immunoprecipitation (ChIP) and quantitative chromatin immunoprecipitation (qChIP) assay
Chromatin Immunoprecipitation Assay kit (Beyotime) was used to perform ChIP analysis according to manufacturer’s protocol. Briefly, CTC-141 cells were grown to 80–95% confluence, washed with cold PBS three times and chemically cross-linked with 1% formaldehyde for 40 min at 37°C. Subsequently, cells were lysed with a lysis buffer at 4°C for 60 min and sonicated (four cycles, each cycle 10 times) at 4°C. Following sonication, 2.5 μg GATA3 antibody and anti-rabbit IgG were added to the lysis solution and incubated at 4°C overnight. Protein A beads were used to isolate GATA3- or IgG-interacted DNA fragments. PCR Purification kit (Qiagen, Hilden, Germany) was used to purify the binding chromatin. Each experiment was performed at least three times.
Luciferase reporter assay
To determine whether GATA3 could bind to the promoter region of ZEB1, we cloned a fragment of human ZEB1 promoter harboring the putative GATA3-binding site into the pGL3 Basic vector (Promega, Madison, WI, USA). CTC-141 cells were co-transfected with GATA3 plasmids, and the ZEB1-pGL3 Basic vector was constructed. The dual luciferase assay system (Promega) was used to detect the firefly and Renilla luciferase activities. Each experiment was performed at least three times.
Statistical analysis
All data were analyzed with Statistical Package for Social Sciences software 18.0 (SPSS Inc., Chicago, IL, USA) and expressed as mean±SD. Correlations between groups were determined using two-sided t-tests. One-way analysis of variance together with Dunnett’s multiple comparison tests was used for multiple group comparisons. P<0.05 was considered statistically significant.
Results
GATA3 is downregulated in GC tissues
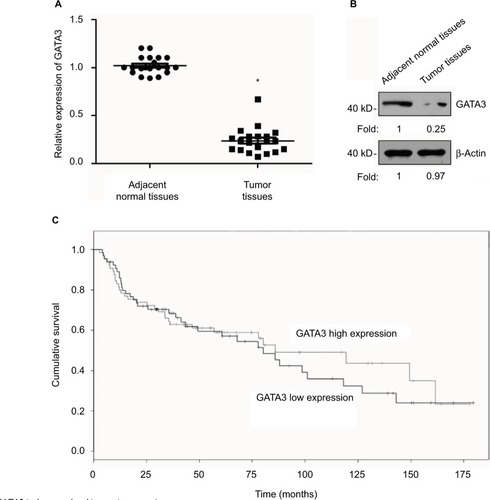
A previous study has reported that GATA3 was downregulated in GC.Citation21 To verify this claim, we collected 83 pairs of tumor tissue samples and adjacent normal tissue samples from GC patients. The expression of GATA3 was detected by qRT-PCR and Western blotting. As shown in , both mRNA level and protein level of GATA3 were significantly downregulated in tumor tissue samples. To further decipher the role of GATA3 in GC, we analyzed the correlation between the expression of GATA3 and clinical data of GC patients. Interestingly, the expression of GATA3 was found to be associated with tumor size, stage and metastasis, but there were no correlations between the expression of GATA3 and gender, age, pathological grade of cancer and alcohol consumption () of patients. We also found that a low expression of GATA2 also predicted a poor prognosis of GC (). Our data confirm that an abnormal expression of GATA3 may play a key role in GC.
Table 1 Clinicopathologic variables in 83 gastric cancer patients
Figure 1 GATA3 is downregulated in gastric cancer tissues.
Abbreviation: qRT-PCR, quantitative real time polymerase chain reaction.
GATA3 is downregulated in GC cells
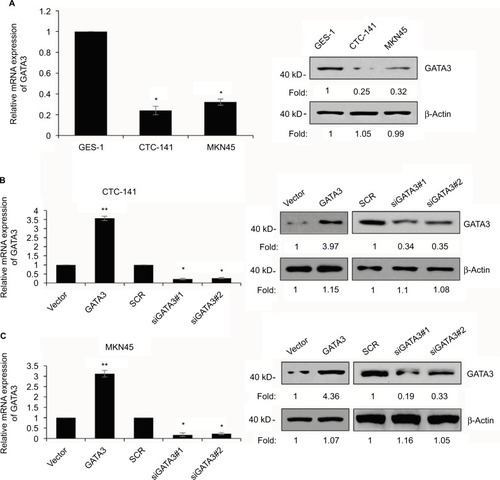
We also found that GATA3 was dramatically downregulated in several gastric cell lines, such as CTC-141 and MKN45, in comparison to normal human gastric mucosal cell line GES-1 (). We first accomplished an overexpression or knocking down of GATA3 in CTC-141 and MKN45. qRT-PCR and Western blotting were performed to detect the expression of GATA3. As shown in , GATA3 was markedly upregulated or knocked down when cells were transfected with FLAG-GATA3 plasmid or GATA3 siRNA, respectively. GATA3 siRNA#1 (siGATA3#1) was found to be more efficient, so it was used for the subsequent experiments.
Figure 2 GATA3 is downregulated in GC cells.
Abbreviations: GC, gastric cancer; SCR, scramble siRNA; qRT-PCR, quantitative real time polymerase chain reaction.
Restoration of GATA3 levels inhibits the proliferation of GC cells
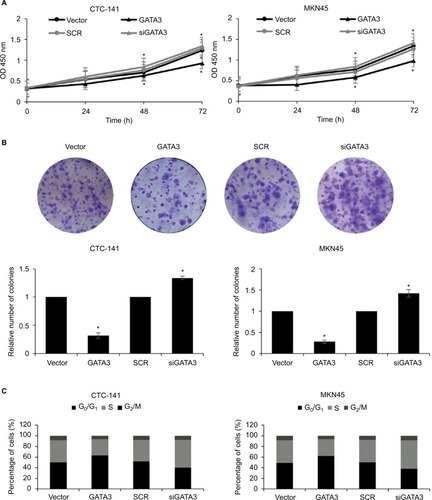
As shown in , a low expression of GATA3 was associated with tumor size, leading us to assume that a downregulation of GATA3 might facilitate the proliferation of GC cells. To determine the role of GATA3 in cell proliferation, CCK-8 assay and colony formation assay were performed. The results suggested that ectopic expression of GATA3 significantly suppressed the viability of CTC-141 cells compared to control groups; however, an inhibition of GATA3 slightly promoted the viability of CTC-141 cells (). Since the expression level of GATA3 in GC cells is low, the effect of knocking down GATA3 is not obvious. Similarly, an overexpression of GATA3 obviously suppressed the viability of MKN45 cells compared to the control group (). From the colony formation assay, it was confirmed that the restoration of GATA3 levels led to a fall in the number of colonies compared to the control group, but GATA3 inhibition also slightly increased the number of colonies (). We further investigated whether GATA3 suppressed cell proliferation through cell cycle regulation. As shown in , ectopic expression of GATA3 largely increased the percentage of cells in G0/G1 phase and decreased the percentage of cells in S+G2/M phase, compared to the vector group. However, the inhibition of GATA3 decreased the percentage of cells in G0/G1 phase and increased the percentage of cells in S+G2/M phase (). Put together, our work shows evidence that the restoration of GATA3 levels inhibited the proliferation of GC cells.
Figure 3 Restoration of GATA3 levels inhibits the proliferation of gastric carcinoma cells.
Abbreviations: CCK-8, Cell Counting Kit-8; SCR, scramble siRNA.
Restoration of GATA3 levels inhibits the migratory and invasive behavior of GC cells
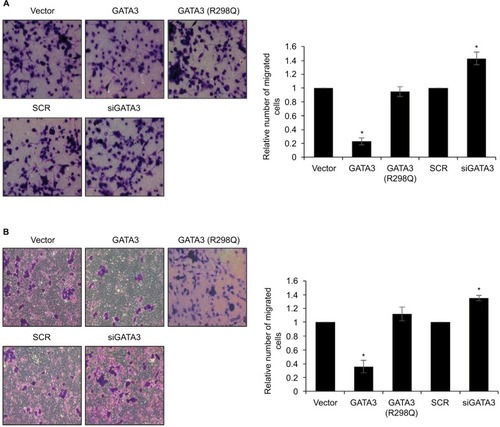
Since a low expression of GATA3 seems to have an association with metastasis, we next detected the effect of GATA3 on cell migration and invasion. Transwell migration and invasion analysis was performed to determine the effect of GATA3 on the migratory and invasive behavior of CTC-141 cells. The results of Transwell migration analysis showed that the migratory behavior of CTC-141 cells that overexpressed GATA3 was obviously reduced compared to the control group, mutant GATA3 (R298Q) had little effect (). However, the migratory behavior of CTC-141 cells that knocked down GATA3 was slightly increased (). Furthermore, Transwell invasion analysis revealed that ectopic expression of GATA3 significantly suppressed the invasive behavior of CTC-141 cells compared to the control groups, mutant GATA3 (R298Q) had little effect; however, inhibition of GATA3 slightly promoted the invasive behavior of CTC-141 cells (). These data suggest that GATA3 suppresses the migration and invasive behavior of GC cells.
Figure 4 Restoration of GATA3 levels inhibits the migratory and invasive behavior of gastric cancer cells.
Abbreviation: SCR, scramble siRNA.
GATA3 inhibits EMT in GC cells
EMT is a complex process which plays a key role in cancer metastasis. Several reports have shown that the GATA family has a significant role to play in EMT.Citation23,Citation24 To further explore the mechanism of GATA3 in GC metastasis, we determined whether GATA3 regulated EMT in GC cells. As shown in , the expression of epithelial markers, including E-cadherin and α-catenin, was obviously upregulated in CTC-141 cells that overexpressed GATA3, but the expression of mesenchymal markers, such as N-cadherin and vimentin, was significantly downregulated. Moreover, mutant GATA3 (R298Q) had little effect on EMT (). However, GATA3 inhibition suppressed the expression of epithelial markers, including E-cadherin and α-catenin, and promoted the expression of mesenchymal markers, including N-cadherin and vimentin ().
Figure 5 GATA3 inhibits epithelial–mesenchymal transition (EMT) in gastric cancer cells.
Abbreviations: SCR, scramble siRNA; qRT-PCR, quantitative real time polymerase chain reaction.
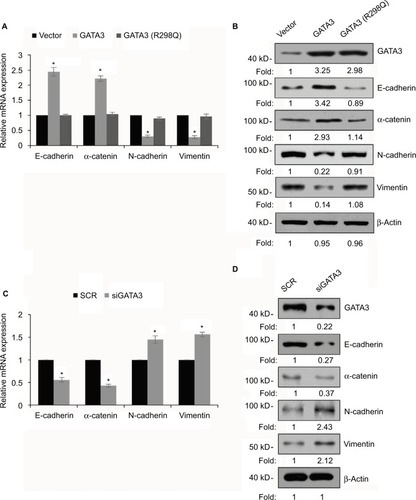
GATA3 transcriptionally regulates ZEB1 in GC cells
EMT was found to be regulated by several transcription factors, including slug, snail and ZEB1.Citation25–Citation27 Thus, we next explored whether GATA3 regulated the expression of these transcription factors. As shown in , we found that only ZEB1 was regulated by GATA3. We next evaluated whether GATA3 transcriptionally regulated ZEB1. ChIP and qChIP assay revealed that GATA3 bounded with the promoter region of ZEB1 (). Moreover, luciferase reporter assay suggested that wild-type GATA3 transcriptionally suppressed ZEB1, but mutant GATA3 (R298Q) had no effect on ZEB1 ().
Figure 6 GATA3 transcriptionally regulates ZEB1 in G cells.
Abbreviations: qRT-PCR, quantitative real time polymerase chain reaction; ChIP, chromatin immunoprecipitation; qChIP, quantitative chromatin immunoprecipitation.
Overexpression of ZEB1 suppresses the migratory and invasive behavior of GC cells
We aimed to detect whether GATA3 regulated the migratory behavior of GC cells through the regulation of ZEB1 after transfecting CTC-141 cells with ZEB1 siRNA. As shown in , the ZEB1 level was obviously lower in CTC-141 cells. ZEB1 siRNA#2 (siZEB1#2) was found to more efficient, so it was used for the subsequent experiments. Subsequently, Transwell migration and invasion assay was carried out to detect the migratory and invasive behavior of each cell group. Consistent with the effect of GATA3 overexpression, knockdown of ZEB1 also dramatically suppressed the migratory and invasive behavior of CTC-141 cells (). These data indicate that GATA3 inhibits the migratory and invasive behavior of GC cells through transcriptionally inhibiting ZEB1.
Figure 7 Overexpression of ZEB1 suppresses the migratory and invasive behavior of gastric cancer cells.
Abbreviations: SCR, scramble siRNA; qRT-PCR, quantitative real time polymerase chain reaction.
Discussion
GATA3, a member of the GATA family, has been found to play a key role in facilitating proliferation and development of several tissues and organs.Citation6,Citation17,Citation28,Citation29 In interfollicular epidermis, GATA3 is required during mouse embryonic development.Citation28 A downregulation of GATA3 in breast carcinoma is correlated with poor clinical outcome.Citation12 Therefore, GATA3 may serve as a novel prognostic biomarker.Citation30
In this study, we detected the expression of GATA3 in GC tissues and cells for the first time. Our results suggest that GATA3 was downregulated in GC tissues and cells. Moreover, the expression of GATA3 was negatively correlated with tumor size and metastasis. Furthermore, a low expression of GATA3 predicted poor prognosis. Similar results have been reported in breast cancer studies.Citation19,Citation20 Our work strongly demonstrated that GATA3 plays a significant role in GC development.
Because cell proliferation, migration and invasion are crucial biological features in cancer development, we performed several functional experiments to detect the effect of GATA3 on these biological processes. CCK-8 and colony formation assay indicated that ectopic expression of GATA3 suppressed cell proliferation in GC. In addition, Transwell migration and invasion assay revealed that ectopic expression of GATA3 suppressed cell migration and invasion in GC. Increasing evidence has suggested the important role of EMT in cancer progression, such as metastasis.Citation31,Citation32 So, we explored the effect of GATA3 on EMT in GC cells by measuring the protein level and mRNA level of EMT-associated markers. As we expected, an overexpression of GATA3 obviously suppressed EMT in GC cells through increasing the expression of epithelial markers, including E-cadherin and α-catenin, and decreasing the expression of mesenchymal markers, including N-cadherin and vimentin. Multiple reports have shown that EMT is regulated by several transcription factors, such as snail, slug and ZEB1.Citation23,Citation24 Luciferase reporter assay and ChIP assay revealed that GATA3 transcriptionally regulated ZEB1, thereby inhibiting EMT in GC. It is better to verify our hypothesis in tumor tissue-derived cells, but all tissue samples in our study have been used for performing qRT-PCR and Western blotting analysis, so the effect of GATA3 on tumor tissue-derived cells will be further decipher in our future research.
In conclusion, our work showed that GATA3 was downregulated in GC tissues and cells. The restoration of GATA3 markedly inhibited cell proliferation, migration, invasion and EMT in GC cells. In addition, we found that GATA3 markedly reduced the expression of ZEB1. These findings confirm that GATA3 serves as an oncogene in GC development.
Acknowledgments
This study was supported by Pudong New District Science and Technology Development fund of Shanghai, item number: PKJ2015-Y31.
Disclosure
The authors report no conflicts of interest in this work.
References
- SiegelRNaishadhamDJemalACancer statistics, 2013CA Cancer J Clin2013631113023335087
- MacdonaldJSSmalleySRBenedettiJChemoradiotherapy after surgery compared with surgery alone for adenocarcinoma of the stomach or gastroesophageal junctionN Engl J Med20013451072573011547741
- ChenCNLinJJChenJJGene expression profile predicts patient survival of gastric cancer after surgical resectionJ Clin Oncol200523297286729516145069
- HippoYTaniguchiHTsutsumiSGlobal gene expression analysis of gastric cancer by oligonucleotide microarraysCancer Res200262123324011782383
- OueNHamaiYMitaniYGene expression profile of gastric carcinoma: identification of genes and tags potentially involved in invasion, metastasis, and carcinogenesis by serial analysis of gene expressionCancer Res20046472397240515059891
- BurchJBRegulation of GATA gene expression during vertebrate developmentSemin Cell Dev Biol2005161718115659342
- GroteDSouabniABusslingerMBouchardMPax 2/8-regulated Gata 3 expression is necessary for morphogenesis and guidance of the nephric duct in the developing kidneyDevelopment20061331536116319112
- PaiSYTruittMLTingCNLeidenJMGlimcherLHHoICCritical roles for transcription factor GATA-3 in thymocyte developmentImmunity200319686387514670303
- HendriksRWNawijnMCEngelJDvan DoorninckHGrosveldFKarisAExpression of the transcription factor GATA-3 is required for the development of the earliest T cell progenitors and correlates with stages of cellular proliferation in the thymusEur J Immunol19992961912191810382753
- TingCNOlsonMCBartonKPLeidenJMTranscription factor GATA-3 is required for development of the T-cell lineageNature199638466084744788945476
- TsarovinaKPattynAStubbuschJEssential role of Gata transcription factors in sympathetic neuron developmentDevelopment2004131194775478615329349
- YoonNKMareshELShenDHigher levels of GATA3 predict better survival in women with breast cancerHum Pathol201041121794180121078439
- Kouros-MehrHSlorachEMSternlichtMDWerbZGATA-3 maintains the differentiation of the luminal cell fate in the mammary glandCell200612751041105517129787
- Cimino-MathewsASubhawongAPIlleiPBGATA3 expression in breast carcinoma: utility in triple-negative, sarcomatoid, and metastatic carcinomasHum Pathol20134471341134923375642
- KaufmanCKZhouPPasolliHAGATA-3: an unexpected regulator of cell lineage determination in skinGenes Dev200317172108212212923059
- TongQDalginGXuHTingCNLeidenJMHotamisligilGSFunction of GATA transcription factors in preadipocyte-adipocyte transitionScience2000290548913413811021798
- ZhengRBlobelGAGATA transcription factors and cancerGenes Cancer20101121178118821779441
- TakakuMGrimmSAWadePAGATA3 in breast cancer: tumor suppressor or oncogene?Gene Expr201516416316826637396
- CioccaVDaskalakisCCioccaRMRuiz-OrricoAPalazzoJPThe significance of GATA3 expression in breast cancer: a 10-year follow-up studyHum Pathol200940448949519084267
- JacquemierJCharafe-JauffretEMonvilleFAssociation of GATA3, P53, Ki67 status and vascular peritumoral invasion are strongly prognostic in luminal breast cancerBreast Cancer Res2009112R2319405945
- KeshariRPWangWZhangYDecreased expression of the GATA3 gene is associated with poor prognosis in primary gastric adenocarcinomaPLoS One201492e8719524504018
- LiuXCaoKXuCGATA-3 augmentation down-regulates Connexin43 in Helicobacter pylori associated gastric carcinogenesisCancer Biol Ther201516698799625901741
- MartinelliPCarrillo-de Santa PauECoxTGATA6 regulates EMT and tumour dissemination, and is a marker of response to adjuvant chemotherapy in pancreatic cancerGut20176691665167627325420
- CampbellKWhissellGFranch-MarroXBatlleECasanovaJSpecific GATA factors act as conserved inducers of an endodermal-EMTDev Cell20112161051106122172671
- DengYLuanFZengLZhangYMaKMiR-429 suppresses the progression and metastasis of osteosarcoma by targeting ZEB1EXCLI J20171661862728694763
- FanFSamuelSEvansKWOverexpression of snail induces epithelial-mesenchymal transition and a cancer stem cell-like phenotype in human colorectal cancer cellsCancer Med20121151623342249
- ToiyamaYYasudaHSaigusaSIncreased expression of Slug and Vimentin as novel predictive biomarkers for lymph node metastasis and poor prognosis in colorectal cancerCarcinogenesis201334112548255724001454
- de Guzman StrongCWertzPWWangCLipid defect underlies selective skin barrier impairment of an epidermal-specific deletion of Gata-3J Cell Biol2006175466167017116754
- PandolfiPPRothMEKarisATargeted disruption of the GATA3 gene causes severe abnormalities in the nervous system and in fetal liver haematopoiesisNat Genet199511140447550312
- ChouJProvotSWerbZGATA3 in development and cancer differentiation: cells GATA have it!J Cell Physiol20102221424919798694
- TakaiMTeraiYKawaguchiHThe EMT (epithelial-mesenchymal-transition)-related protein expression indicates the metastatic status and prognosis in patients with ovarian cancerJ Ovarian Res201477625296567
- HongHZhouTFangSPigment epithelium-derived factor (PEDF) inhibits breast cancer metastasis by down-regulating fibronectinBreast Cancer Res Treat20141481617225284724