
Abstract
Purpose
Liver cancer stem cells (CSCs) are known to be associated with the development, survival, proliferation, metastasis, and recurrence of liver tumors. The aim of this study was to investigate the association of liver-enriched activator protein 1 (LAP1) with hepatocellular carcinoma (HCC) and liver CSCs (LCSCs) and explore the impact of LAP1 on LCSCs.
Materials and methods
Differences in LAP1 expression in liver cancer tissues versus matched para-tumoral liver tissues and LCSCs versus non-CSCs were analyzed by Western blotting, real-time polymerase chain reaction, immunohistochemistry, and flow cytometry. The effect of LAP1 on liver cancer cells was evaluated by the expression of CSC markers, oncosphere formation, proliferation, migration, and invasion in vitro. Cell cycle distribution and the number of apoptotic cells were analyzed to assess cell cycle and cell apoptosis. Furthermore, a mouse subcutaneous tumor implant model was established to explore the role of LAP1 in the development of HCC in vivo. Finally, the expression of CSC markers in paraffin-embedded sections was evaluated by immunofluorescence.
Results
LAP1 was weakly expressed in HCC tumors and cell lines and even weaker in LCSCs. LAP1 inhibited the expression of stem cell–associated genes and reduced the abilities of oncosphere formation, proliferation, migration, and invasion in vitro. Cell cycle assay revealed that LAP1 induced G1/G0 arrest. Furthermore, LAP1 decreased subcutaneous tumor-formation ability and the expression of CSC markers and Ki67 in vivo.
Conclusion
LAP1 suppressed the stem cell features of HCC, indicating that it possessed an antitumor effect in liver cancer, both in vitro and in vivo; therefore, LAP1 may prove to be a potential target in liver CSC-targeted therapy.
Introduction
Hepatocellular carcinoma (HCC) is one of the most common malignancies in the People’s Republic of China. Histologically, HCC is the most common type of primary liver cancer, accounting for 85%–90% of all primary liver cancers.Citation1 The incidence of liver cancer ranks fifth, and mortality from HCC ranks third among all malignancies.Citation1–Citation3 Although continuous progress has been made in the diagnosis and surgical and biological therapies of liver cancer, the overall prognosis of patients with liver cancer remains unsatisfactory due to recurrence, metastasis, chemoresistance, and radiation resistance.Citation4 Therefore, more efforts are required to explore novel and more effective treatments for liver cancer.
Cancer stem cells (CSCs) play important roles in the development, metastasis, drug resistance, and many other biological processes of tumors.Citation5–Citation7 The concept of CSCs was first proposed by Mackillop et al,Citation8 who proposed that there may be a small number of cells in cancer that had similar functions as stem cells. In 1997, Bonnet and DickCitation9 identified CD34+/CD38− in leukemia as CSCs. Subsequent research further identified CSCs in various solid tumors, including cancers of the breast, lung, prostate, brain, and pancreas.Citation10–Citation13 Meanwhile, many studies confirmed that CSCs also exist in HCC and could be enriched by several defined surface markers such as CD133, CD13, CD44, OV6, epithelial cell adhesion molecule (EpCAM), CD24, and CD90.Citation6,Citation14,Citation15 According to the CSC theory, CSCs contribute to tumorigenesis, metastasis, recurrence, and chemoresistance through cell symmetry and asymmetric division;Citation16 therefore, CSCs could be used as the target in tumor therapy.Citation17,Citation18
CCAAT/enhancer-binding protein beta (CEBPβ) – a liver-enriched transcription factor – contains three isoforms, including two liver-enriched activating protein isoforms, LAP1 and LAP2, and a liver-enriched inhibitory protein isoform called LIP.Citation19 CEBPβ not only regulates the differentiation, proliferation, and survival of hepatocytes but also plays a critical role in inflammatory response and tumorigenesis.Citation20–Citation22 Abnormal expression of CEBPβ is closely associated with the malignancy of a variety of tumors such as glioma, Wilm’s tumor, renal cell tumor, and ovarian cancer.Citation23–Citation25 LAP1 – the longest isoform – is the most highly expressed isoform of CEBPβ. LAP1 is implicated in cancer progression. LAP1 was reported to repress migration and invasion of HCC cells in vitro by binding to the ORM2 promoter and activating its expression.Citation26 However, little is known about the function of LAP1 in liver CSCs (LCSCs).
In the present study, we explored the association of LAP1 with HCC and LCSCs and the effect of LAP1 on LCSCs. CEBPβ and LAP1 were found to be weakly expressed in HCC and even weaker in LCSCs. LAP1 suppressed the expression of marker genes of LCSCs and reduced capacities of oncosphere formation, proliferation, wound healing, migration, and invasion in vitro. More importantly, LAP1 decreased subcutaneous tumorigenicity in vivo. These data suggest that LAP1 could be used as an indicator of the diagnosis and prognosis of HCC and may be developed as a target for liver cancer therapy in the future.
Materials and methods
Cell lines and tumor specimens
Human liver cancer cell lines Huh7, Hep3B, and HepG2 were purchased from the American Type Culture Collection and stored in our laboratory. LM3 was purchased from the Cell Bank of the Type Culture Collection of the Chinese Academy of Sciences. All liver cancer cell lines were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 10% fetal bovine serum (FBS; HyClone), 100 mg/mL penicillin, and 100 U/mL streptomycin. The primarily cultured human liver cancer cell line CLC13 from liver cancer samples was kindly provided by Dr Lijian Hui from the Chinese Academy of Sciences (Shanghai, People’s Republic of China).Citation27 CLC13 cells were cultured in RPMI1640 (HyClone), supplemented with 10% FBS (Gibco), 110 µg/mL sodium pyruvate, 10 µg/mL insulin, 5.5 µg/mL transferrin, 40 ng/mL epidermal growth factor (EGF), and 6.7 ng/mL sodium selenite (Gibco). The use of CLC13 was approved by the ethical committees of Shanghai Xuhui District Central Hospital and Shanghai East China Hospital. Human liver cancer specimens were obtained from the Eastern Hepatobiliary Surgery Hospital/Institute (Shanghai, People’s Republic of China), with written informed consent from the patients and approval from the Institutional Review Board of Shanghai Eastern Hepatobiliary Surgery Hospital.
Western blot
Tumor cells or tumor tissues were lysed in radio immunoprecipitation assay buffer for Western blotting. The proteins were separated on 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and electrophoretically transferred onto polyvinylidene difluoride membranes (Millipore), which were then blocked in tris-buffered saline and Tween 20 (TBST) containing 5% defatted milk for 1 h at room temperature and incubated with primary antibodies overnight at 4°C with mouse anti-tubulin (1:10,000; Proteintech Group, Chicago, IL, USA) and rabbit anti-CEBPβ (1:1,000; Santa Cruz Biotechnology, Santa Cruz, CA, USA). After being washed with TBST and incubated with either horseradish peroxidase (HRP)-conjugated goat anti-rabbit (1:5,000; Proteintech Group) or HRP-conjugated goat anti-mouse (1:5,000; Proteintech Group) secondary antibody, immune complexes were visualized by using Pierce™ enhanced chemiluminescent (ECL) Western Blotting Substrate (Thermo Fisher Scientific).
Quantitative real time-PCR (qRT-PCR)
Total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), and reverse transcription was undertaken using M-MLV Reverse Transcriptase (Promega, M530A) according to the manufacturer’s instructions. Quantitative PCR was conducted using LightCycler 480 (Roche) with SYBR Premix Ex Taq reagent (TaKaRa). All samples were examined in triplicate. Data were normalized to control samples. The primers are shown in .
Hematoxylin and eosin (H&E), immunohistochemistry, and immunofluorescence staining
Paraffin-embedded HCC sections from 14 patients were deparaffinized with xylene and rehydrated. For H&E staining, the rehydrated sections were stained with H&E (Sigma-Aldrich). For immunohistochemical (IHC) staining, the rehydrated sections were immersed in citrate buffer (pH 6.0) for antigen retrieval at 121°C for 2 min and then cooled down to room temperature slowly. The endogenous peroxidase activity was blocked with 3% hydrogen peroxide (H2O2) for 10 min. Nonspecific binding was blocked with 1% bovine serum albumin for 20 min at room temperature. The sections were incubated with rabbit anti-CEBPβ antibodies (1:200; Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 4°C overnight in a humidified chamber, then incubated with HRP-conjugated goat anti-rabbit antibodies (1:500; Proteintech Group) at 37°C for 30 min, and then detected with 3,3′-diaminobenzidine (DAB) (Dako, Glostrup, Denmark) and counterstained with Mayer’s hematoxylin solution (Sigma-Aldrich). Seeing that there existed differences from IHC staining, rabbit anti-Ki67 (1:200; Invitrogen, Carlsbad, CA, USA) and mouse anti-CD133 (Proteintech Group, Chicago) and mouse anti-CD13 (Proteintech Group) antibodies were used for immunofluorescence (IF) staining. Nuclei were labeled with the blue fluorescent Hoechst 33342 dye.
Colony-formation assays
For colony-formation assays, 1,000 Huh7 cells or 2,000 CLC13 cells per well were plated on six-well plates and cultured for 7 days. Then, cells were fixed with 4% formaldehyde for 10 min and stained with crystal violet solution. The number of clones (>50 cells) was counted microscopically.
CCK8 assays
For CCK8 assays, 1,200 Huh7 cells or 3,000 CLC13 cells per well were plated on 96-well plates and incubated for 24 h. Then, 10 µL CCK8 reagent was added to each well and incubated for 2 h at 37°C. The optical density value was measured at a wavelength of 450 nm by using a microplate reader every day for 7 days.
Flow cytometry and cell sorting
Cells were dissociated into single cells by trypsinization and labeled with BV421-CD13 human antibody (301716, Biolegend, San Diego, CA, USA) at 4°C for 15 min. The concentration of the antibody used was according to the manufacturer’s recommendation. The stained cells were analyzed or collected with the FACS Calibur machine (BD Biosciences).
Cell cycle and cell apoptosis
Cell cycle and cell apoptosis were assessed by the Cell Cycle Assay Kit (CCS012, MULTI Sciences) and the Annexin V-APC/7-AAD Apoptosis Detection Kit (AP105, MULTI Sciences), respectively, following the manufacturer’s instructions.
Sphere-formation assay
Hepatoma cells were plated on six-well ultra-low attachment culture plates (Corning) in DMEM/F12 supplemented with B27, N2, 20 ng/mL EGF and 20 ng/mL basic fibroblast growth factor (bFGF), and cultured in a CO2 incubator for 7 days. The number of spheroids was counted under a microscope.
For separation of oncospheres and nonsphere cells, we collected sphere-formation medium containing both nonsphere and sphere cells in a 15-mL centrifuge tube. Pellets formed after 5-min tranquilization were recognized as spheres. The supernatant was then moved into a new centrifuge tube carefully and centrifuged at 1,500g for 5 min to obtain nonspheric pellets.
In vitro migration and invasion assays
Cell migration was assessed by wound-healing and Transwell assays. For the wound-healing assay, a scratch was made in the center of the culture dish using a sterile 20-µL pipette tip before cells were fully confluent. The wound was observed using a bright-field microscope at 24- and 48-h after scratching. The migration rate = (scratch widths before migration – scratch widths after migration)/scratch widths before migration ×100%. Transwell migration assay was conducted using 24-well Transwell Chambers without Matrigel (BD Biosciences) according to the manufacturer’s instructions. Briefly, 1×105 Huh7 cells or 1×105 CLC13 cells in medium with 10% FBS were added to the top chamber, and the medium containing 20% FBS was added to the bottom chamber as an attractant. After 48-h incubation, the migrated cells were fixed and stained with crystal violet. The number of migrated cells was counted in 10 fields under a 10× objective lens.
Transwell invasion assay was conducted using 24-well BioCoat Matrigel Invasion Chambers (BD Biosciences) according to the manufacturer’s instructions. Briefly, 4×105 Huh7 cells in medium with 10% FBS were added to the top chamber, and the medium containing 20% FBS was added to the bottom chamber as an attractant. The remaining steps are the same as those described for the Transwell migration assay.
In vivo xenograft experiments
For tumor growth assays, 5×105 Huh7 cells or 1×106 CLC13 cells were subcutaneously injected into 6-week-old male BALB/c nude mice. Every 3 days, tumor volume was calculated by the formula V=0.5W2L (V, volume; L, length; and W, width). BALB/c nude mice were obtained from the Animal Centre of the Chinese Academy of Medical Sciences, they were fed in standard pathogen-free conditions, and all animal experiments were in accordance with the Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee of the Second Military Medical University.
Statistical analysis
All data are presented as the mean±SD. Statistical methods were indicated in figure legends, and statistical computations were conducted using GraphPad Prism version 6.0. P<0.05 was considered statistically significant; *, **, and *** indicate P<0.05, P<0.01, and P<0.001, respectively.
Results
LAP1 is weakly expressed in HCC
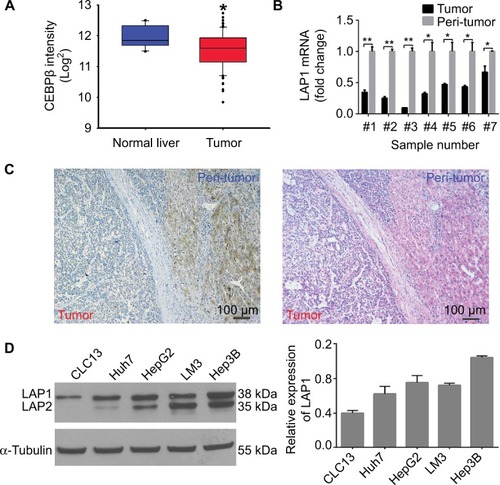
The result of an online-available HCC transcriptome dataset (GSE62232) analysis showed that CEBPβ was weakly expressed in HCC tumors (, Student’s t-test, *P<0.05). In addition, qRT-PCR showed that the expression level of LAP1 mRNA in HCC tissues was significantly lower than that in seven pairs of matched paratumoral liver tissues (, Student’s t-test, *P<0.05). These findings were further validated by IHC staining. At the same time, H&E staining of the same tissue samples was done to confirm the pathological type of HCC (). The result showed that LAP1 was significantly downregulated in HCC tumor tissues as compared with para-tumoral tissues. Then, we measured the expression level of CEBPβ in five human hepatoma cell lines (Huh7, Hep3B, HepG2, LM3, and CLC13) and found that Huh7 and CLC13 cell lines contained the lowest amounts of LAP1 among these cell lines () and, therefore, these two cell lines were used in our further experiments.
Figure 1 LAP1 is weakly expressed in HCC.
Notes: (A) CEBPβ was weakly expressed in patients with HCC. CEBPβ expression in HCC tumor and normal liver tissues provided by Park’s cohort (GSE62232) was analyzed. (B) LAP1 expression levels were verified in samples of patients with HCC by qRT-PCR. GAPDH served as a loading control. (C) HCC samples were assayed by immunohistochemical and H&E staining. Scale bar: 100 µm. (D) The endogenous levels of liver-enriched activating protein and liver-enriched inhibitory protein were determined by Western blot using rabbit IgG anti-CEBPβ (sc-150) and horseradish peroxidase-labeled anti-rabbit IgGs. The α-tubulin protein level was used as a loading control (left panel). For each cell, three independent protein samples were analyzed, and each band was quantified and LAP1 protein expression was normalized with the expression level of the α-tubulin gene. Mean ± SD are represented (right panel). Student’s t-test was used for statistical analysis, *P<0.05 and **P<0.01; data are shown as mean ± SD. Data are representative of at least three independent experiments.
Abbreviations: CEBPβ, CCAAT/enhancer binding protein beta; HCC, hepatocellular carcinoma; IgG, immunoglobulin G; LAP1, liver-enriched activator protein 1; qRT-PCR, quantitative real time polymerase chain reaction.
LAP1 is weakly expressed in LCSCs
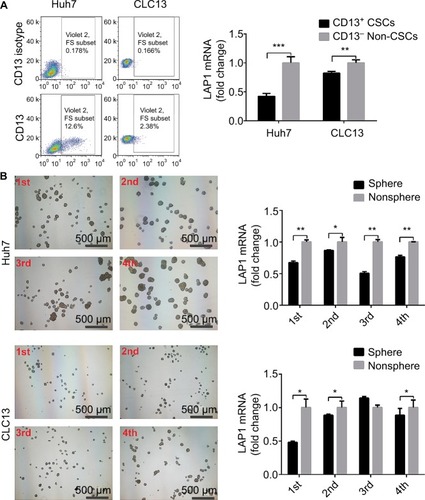
Knowing that CSCs play complex roles in the development and progression of cancer, we further investigated the expression of LAP1 in LCSCs. CD13 is known to be a target for identifying CSCs in a proportion of HCC; therefore, we sorted CD13+ cells from Huh7 and CLC13 HCC cell lines as LCSCs. Then, we tested the LAP1 expression in CD13+ CSCs and CD13− non-CSCs of the two cell lines and found that the LAP1 expression in CD13+ CSCs was significantly lower than that in CD13− non-CSCs (). Furthermore, we conducted an oncosphere experiment, and the results showed that LAP1 expression in CSC-enriched spheres was significantly lower than that in nonspheres. Importantly, through serial passage of Huh7 and CLC13 sphere cells, similar observations were obtained in the following three generations of oncosphere assay (). Altogether, LAP1 was weakly expressed in HCC tumor tissues and LCSCs.
Figure 2 LAP1 is weakly expressed in liver CSC.
Notes: (A) LAP1 was weakly expressed in CD13+ cells sorted from Huh7 and CLC13 cells. LAP1 messenger RNA (mRNA) was measured by qRT-PCR. (B) LAP1 was even more weakly expressed in oncospheres than in nonsphere tumor cells; through serial passage of Huh7 and CLC13 sphere cells, similar observations were obtained in the first, second, third, and fourth generations of oncosphere assay. Nonsphere: Huh7 or CLC13 cells that failed to form spheres. Scale bar: 500 µm. Student’s t-test was used for statistical analysis, *P<0.05, **P<0.01, ***P<0.001; data are shown as mean ± SD. Data are representative of at least three independent experiments.
Abbreviations: CSC, cancer stem cells; LAP1, liver-enriched activator protein 1; qRT-PCR, quantitative real time polymerase chain reaction.
LAP1 inhibits the stemness of LCSCs
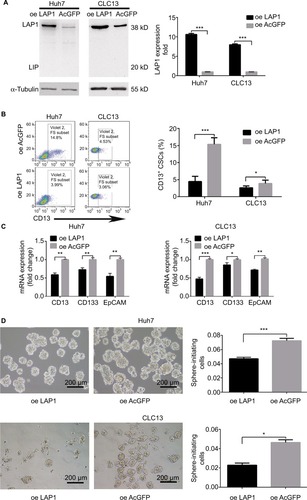
To explore the function of LAP1 in HCC, we constructed Huh7 and CLC13 cells stably expressing LAP1 by using the lentivirus infection system, and an pLVX-AcGFP1-N1 (AcGFP)-overexpressing empty vector was used as the control (). Notably, flow cytometric analysis revealed that LAP1 overexpression decreased the number of CD13+ liver CSCs markedly (). Compared with the control, the mRNA level of CSC markers CD133, CD13, and EpCAM was significant downregulated in both Huh7 and CLC13 cells infected with LAP1 (). Furthermore, sphere-formation experiments showed that LAP1 overexpression dramatically decreased oncosphere formation as compared with the AcGFP-overexpressing group (). These data indicate that LAP1 overexpression suppressed the stemness features of LCSCs.
Figure 3 LAP1 inhibits the stemness of liver CSCs.
Notes: (A) LAP1-overexpressing Huh7 cells and control Huh7 cells were established. LAP1-overexpressing CLC13 cells and control CLC13 cells were established (left panel). For each cell, three independent RNA samples were analyzed and CEBPβ mRNA expression was normalized with the expression level of the GAPDH gene. Mean ± SD are represented (right panel). (B) CD13+ (CSC) subpopulations were detected in LAP1-overexpressing Huh7 and LAP1-overexpressing CLC13 cells and their control cells by fluorescence-activated cell sorting (FACS) analysis. Results are shown as mean ± SD. (C) Expression of CD13, CD133, and EpCAM (stemness-associated transcription factors) in LAP1-overexpressing Huh7 and LAP1-overexpressing CLC13 cells, and their control cells were compared by real-time PCR. Significant downregulation in CD13, CD133, and EpCAM mRNA levels was detected in both cell lines after overexpressing LAP1. Results are shown as mean ± SD. (D) LAP1 overexpression causes a diminished oncosphere-forming capacity in Huh7 and CLC13 cells. The right panel represents statistical results as mean ± SD. Scale bar, 200 µm. *P<0.05, **P<0.01, ***P<0.001.
Abbreviations: CEBPβ, CCAAT/enhancer binding protein beta; CSC, cancer stem cell; EpCAM, epithelial cell adhesion molecule; LAP1, liver-enriched activator protein 1; oe, overexpression; PCR, polymerase chain reaction.
LAP1 suppresses the proliferation of HCC cell lines in vitro
Seeing that LAP1 inhibited the stemness features of LCSCs, we explored whether LAP1 played a critical role in the expansion of HCC cell lines. We conducted CCK8 cell-proliferation and colony-formation assays to explore the effect of LAP1 expression on the proliferation of HCC cells. Compared with the control, LAP1 overexpression suppressed the proliferation of Huh7 and CLC13 cells markedly (). The colony-formation assay showed that LAP1 overexpression could also markedly reduce the frequency of colony formation in Huh7 and CLC13 cells ().
Figure 4 LAP1 suppress the proliferation of HCC cell line in vitro.
Notes: (A) The suppression rate was up to more than 50% on the sixth day after LAP1 infection, compared with AcGFP and noninfected control groups (one-way analysis of variance). *P<0.05, **P<0.01, and ***P<0.001. (B and C) Both HCC cell lines infected with LAP1 formed fewer and smaller colonies than those infected with AcGFP. Each value represents the mean ± SD for triplicate samples (Student’s t-test). *P<0.05, **P<0.01, and ***P<0.001. (D) The cell cycle transition of Huh7 and CLC13 cells was examined, and the increase of the percentage of cells in G1/G0 phase was caused in both cell lines by LAP1 overexpression. All experiments were repeated at least three times, and representative data are shown. Each value represents the mean ± SD for triplicate samples (Student’s t-test). *P<0.05, **P<0.01, and ***P<0.001.
Abbreviations: HCC, hepatocellular carcinoma; LAP1, liver-enriched activator protein 1; oe, overexpression.
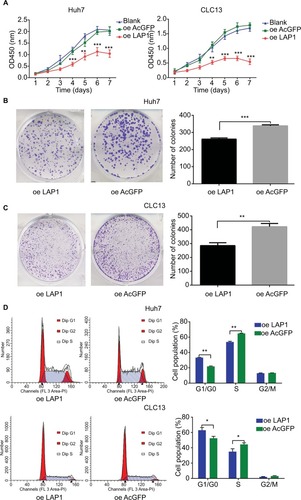
To distinguish between the possibilities that LAP1 overexpression inhibited growth or caused cell death in hepatoma cells, we undertook flow cytometry to quantitate the population of apoptotic cells and cell cycle distribution. Compared with AcGFP overexpression, LAP1 overexpression promoted G1/G0 phase arrest in Huh7 and CLC13 cells (), whereas no cell apoptosis was observed in these two cell lines (Figure S1).
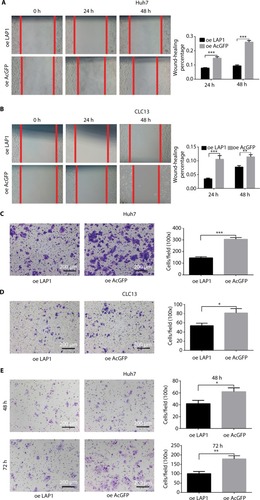
LAP1 inhibits the migration and invasion of HCC
To explore the function of LAP1 on the migration and invasion of HCC cells, wound-healing and Transwell migration assays were conducted. It was found that LAP1 suppressed the migratory ability of Huh7 and CLC13 cells efficiently (). The result of a Matrigel invasion assay showed that LAP1 suppressed the invasive ability of Huh7 cells (). These findings demonstrate that LAP1 possesses antitumor activity in HCC cells.
Figure 5 LAP1 inhibits the migration and invasion of HCC.
Notes: (A and B) A wound-healing assay showed that cell motility was suppressed by LAP1 overexpression in Huh7 and CLC13 cells. Microscopic images were acquired at 0, 24, and 48 h after scratching the surface of a confluent layer of cells. (C and D) The in vitro migration ability of Huh7 and CLC13 cells infected with LAP1 was assessed using Transwell assays. Cells infected with AcGFP were used as a control. Scale bar, 200 µm. (E) Matrigel invasion assay was conducted to compare the cell invasion ability between LAP1-overexpressing and control Huh7 cells after 48- and 72-h infection. Invaded cells were fixed and stained with crystal violet, Scale bar, 200 µm. The number of invaded cells was calculated and is shown in the bar chart. All data are mean ± SD of three independent experiments (*P<0.05, **P<0.01, ***P<0.001, Student’s t-test).
Abbreviations: LAP1, liver-enriched activator protein; oe, overexpression.
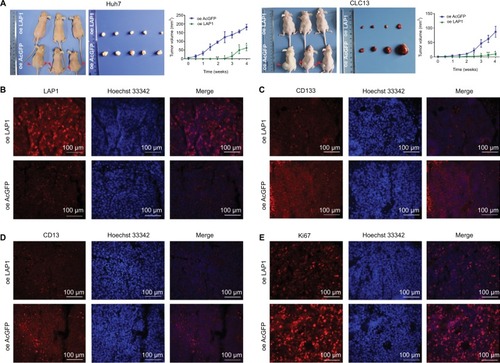
LAP1 suppressed the tumor proliferation and stemness of HCC in vivo
For the in vivo tumor-formation assay, tumor formation was observed in nude mice injected with LAP1-transfected and AcGFP empty vector-transfected Huh7 and CLC13 cells. In addition, the tumor size and tumor volume in the LAP1 overexpression group were significantly smaller than those in the AcGFP empty vector-transfected cell group. The tumor size was measured every 3 days and, after 1 month, the mice were sacrificed, and the tumors were removed and weighed (). Then, the tumors were paraffin embedded and IF stained. Using a rabbit CEBPβ antibody, the overexpression of LAP1 was confirmed (). Furthermore, the result showed that the intensity of CD13, CD133, and Ki67 was obviously lower in LAP1-overexpressing CLC13 cells than in the control, indicating that LAP1 overexpression was negatively correlated with the LCSC markers CD13 and CD133 as well as Ki67 (). Taken together, LAP1 overexpression inhibited xenograft tumor growth and stemness markedly, suggesting that LAP1 plays a tumor suppressor role in HCC.
Figure 6 LAP1 suppresses the tumor proliferation and stemness of HCC in vivo.
Notes: (A) LAP1-overexpressing and AcGFP-overexpressing Huh7 cells (5×105) and LAP1-overexpressing and AcGFP-overexpressing CLC13 cells (1×106) were subcutaneously injected into BALB/c nude mice for observation of tumor growth. Tumor sizes were observed every 3 days and photographed weekly, and representative pictures are shown. After a 4-week tumor observation, the animals were sacrificed and tumors were removed and weighed. Growth curves of tumors over 4 weeks are summarized and analyzed. Results are shown as mean ± SD, n=6 for each group. (B) The xenograft from the nude mice originated from CLC13 cells were paraffin embedded and IF stained. The level of CEBPβ was monitored using a rabbit CEBPβ antibody, and the overexpression of LAP1 was confirmed. Scale bars: 100 µm. (C–E) The level of CD13 or CD133 or Ki67 was compared and analyzed with corresponding antibodies and then counterstained with Hoechst 33342 for confocal microscopy. The images are representative of the results obtained. Scale bars, 100 µm.
Abbreviations: CEBPβ, CCAAT/enhancer binding protein beta; HCC, hepatocellular carcinoma; IF, immunofluorescence; LAP1, liver-enriched activator protein 1; oe, overexpression.
Discussion
Several studies have highlighted the crucial role of CEBPβ in suppressing tumor features. For example, forced expression of CEBPβ in HepG2 hepatocarcinoma cells induced arrest of cells at or near the G1–S boundary.Citation28 CEBPβ knockout mice developed a lymphoproliferative disorder, suggesting that CEBPβ inhibited the expansion of the lymphoid cell compartment.Citation29 CEBPβ was found to be expressed at a low level in hematological malignancies in chronic myelogenous leukemia. CEBPβ played a role in the BCR/ABL of bone marrow progenitor cells by activating the transcription and expression of the downstream target gene GADD45A, thus promoting cell differentiation and inhibiting cell proliferation.Citation30
However, growth arrest induced by CEBPβ is highly context specific because CEBPβ was found to display growth-promoting activity in several cases. CEBPβ can function as a pro-oncogenic transcription factor that promotes proliferation and/or survival of some tumor cells. For example, deletion of the CEBPβ gene rendered mice totally resistant to carcinogen-induced skin tumor development.Citation31 Shimizu et alCitation32 reported that CEBPβ inhibited apoptosis and improved the survival rate of pancreatic cancer cells. Gardiner et alCitation33 found that CEBPβ-1 overexpression increased transformation and upregulated the expression of the CSC marker ALDH1A1, leading to chemoresistance in Ewing sarcoma. Therefore, CEBPβ can function as a pro-oncogenic transcription factor that promotes proliferation and survival of some tumor cells. How it stimulates mitotic growth and why it elicits a completely opposite effect on proliferation in different cellular contexts are intriguing questions for future investigation.
The incidence and mortality of HCC are at the forefront of all malignancies.Citation1,Citation3 Although great progress has been made in the diagnosis and treatment of liver cancer, tumor recurrence and metastasis in patients with liver cancer remain the main reasons for poor prognosis.Citation4 CSCs have some characteristics including self-renewal, differentiation, and the ability to form a new tumor.Citation34 Studies have confirmed that CSCs may be responsible for cancer development, relapse, and metastasis. Human HCC is driven and maintained by LCSCs. Thus, targeting CSCs may become a promising therapeutic strategy to cure deadly malignancies.
Furthermore, CEBPβ expression has been linked to a variety of rodent and human cancers.Citation20 CEBPβ appears to play a critical role in the development of both the mammary gland and cancers through its involvement in the development, differentiation, and proliferation of mammary epithelial cells.Citation35–Citation37
LAP1 is the most highly expressed isoform of CEBPβ, which is a liver-enriched transcription factor involved in cellular metabolism, development, proliferation, and differentiation. In this study, our observations were confirmed by Park’s cohort dataset GSE62232. We found that LAP1 was weakly expressed in HCC tumors and LCSCs, indicating that LAP1 may play a negative regulatory role in CSCs. However, no study has explored the function of LAP1 in LCSCs and other kinds of CSCs.
The self-renewal capacity of CSCs is known to be one of the important features for the development of cancer. To explore the function of LAP1 on the self-renewal capacity of LCSCs, we first measured the effect of LAP1 on the proliferative capacity of HCC cell lines. The results showed that LAP1 overexpression was able to inhibit the proliferative capacity of HCC cells, and that LAP1 could attenuate the stemness of HCC. Moreover, stem cell–associated membrane markers can be used to measure the stemness of HCC cells. Knowing that CD133, CD13, and EpCAM are the most useful stemness biomarkers, we conducted qRT-PCR and IF to measure their expressions, and found that LAP1 could decrease the expression of CD133, CD13, and EpCAM, thus suggesting that LAP1 could attenuate the stemness of HCC and inhibit the self-renewal ability of LCSCs. Furthermore, the sphere-formation experiment was conducted, knowing that it is an important experiment to measure the self-renewal capacity of HCC. We found that LAP1 overexpression could inhibit the sphere formation of HCC in terms of the size and number of the spheres. According to the theory of CSCs, the subcutaneous tumor-formation ability is an important method to measure the self-renewal capacity of CSCs in mice. To explore the influence of LAP1 on the stemness of LCSCs, we transplanted HCC to the mice and built a subcutaneous tumor mice model. After approximately 1 month, differences between the LAP1 overexpression and control groups were observed. It was found that LAP1 could significantly inhibit the formation and growth of subcutaneous tumors; furthermore, when the tumors were paraffin embedded and IF stained, we found the intensities of CD13, CD133, and Ki67 were obviously lower in LAP1 overexpressing CLC13 cells than in the control, which strongly supports the conclusion that LAP1 can attenuate the stemness of LCSCs and HCCs. At the same time, we measured other tumor biological functions of LAP1 by the Transwell assay and scratch test, and found that LAP1 overexpression could inhibit the migration and invasion of HCC in vitro. Furthermore, LAP1 overexpression could cause cell cycle arrest in the G1/G0 phase, which also indicates the tumor suppressor gene function of this LAP1.
The above results demonstrated that LAP1 could suppress the self-renewal capability of LCSCs and may prove to be a target for eradicating LCSCs and a novel promising therapeutic approach for inhibiting liver cancer.
Conclusion
LAP1 was weakly expressed in HCC and LCSCs. LAP1 could suppress the self-renewal capability of LCSCs and possessed a tumor-suppressive effect on HCC cells, suggesting that LAP1 may serve as an optimal target in liver CSC-targeted therapy.
Acknowledgments
This study was supported by the National Natural Science Foundation of China (grant numbers 31771511 and 31401166), Shanghai Committee of Science and Technology of China (grant number 15JC1403900), and China Postdoctoral Science Foundation (grant number 2013M542434).
Supplementary materials
Figure S1 After infection by LAP1 and AcGFP for 48 and 72 h, the cell apoptosis of Huh7 and CLC13 was examined. Cell apoptosis was not observed in both cells. The experiments were repeated three times, and representative data are shown. Each value represents the mean ± SD for triplicate samples (Student’s t-test). No significance (P>0.05).
Abbreviations: LAP1, liver-enriched activator protein 1; oe, overexpression.
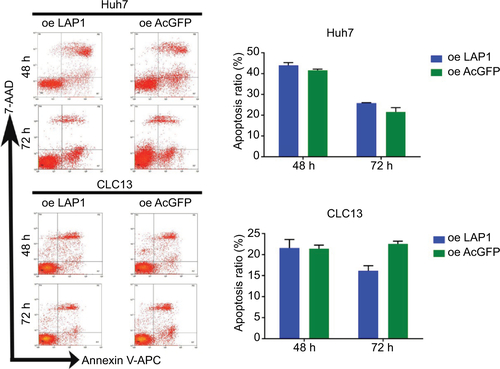
Table S1 Real-time PCR primers used in this study
Author contributions
All authors contributed to the conception and design of the study, acquisition of data, or analysis and interpretation of the data, and to the critical revision of the manuscript for important intellectual content, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
- TorreLABrayFSiegelRLFerlayJLortet-TieulentJJemalAGlobal cancer statistics, 2012CA Cancer J Clin20156528710825651787
- WangRSunQWangPNotch and Wnt/β-catenin signaling pathway play important roles in activating liver cancer stem cellsOncotarget2016755754576826735577
- SiegelRLMillerKDJemalACancer statistics, 2016CA Cancer J Clin201666173026742998
- MaluccioMCoveyARecent progress in understanding, diagnosing, and treating hepatocellular carcinomaCA Cancer J Clin201262639439923070690
- HaoJZhangYDengMMicroRNA control of epithelial-mesenchymal transition in cancer stem cellsInt J Cancer201413551019102724500893
- ChibaTIwamaAYokosukaOCancer stem cells in hepatocellular carcinoma: therapeutic implications based on stem cell biologyHepatol Res2016461505726123821
- Charafe-JauffretEGinestierCIovinoFBreast cancer cell lines contain functional cancer stem cells with metastatic capacity and a distinct molecular signatureCancer Res20096941302131319190339
- MackillopWJCiampiATillJEBuickRNA stem cell model of human tumor growth: implications for tumor cell clonogenic assaysJ Natl Cancer Inst19837019166571928
- BonnetDDickJEHuman acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cellNat Med199737307379212098
- MacDonaghLGraySGBreenELung cancer stem cells: the root of resistanceCancer Lett2016372214715626797015
- TangDGPatra WalaLCalhounTProstate cancer stem/progenitor cells: identification, characterization, and implicationsMoll Carcinogen2007461114
- LathiaJDMackSCMulkearns-HubertEEValentimCLRichJNCancer stem cells in glioblastomaGenes Dev201529121203121726109046
- LeeCJDoschJSimeoneDMPancreatic cancer stem cellsJ Clin Oncol200826172806281218539958
- TongCMMaSGuanXYBiology of hepatic cancer stem cellsJ Gastroenterol Hepatol20112681229123721557770
- JiJWangXWClinical implications of cancer stem cell biology in hepatocellular carcinomaSemin Oncol201239446147222846863
- YamashitaTWangXWCancer stem cells in the development of liver cancerJ Clin Invest201312351911191823635789
- YangQWangYYangQCuprous oxide nanoparticles trigger ER stress-induced apoptosis by regulating copper trafficking and overcoming resistance to sunitinib therapy in renal cancerBiomaterials2017146728528898759
- YuBWangYYuXCuprous oxide nanoparticle-inhibited melanoma progress by targeting melanoma stem cellsInt J Nanomedicine2017122553256728435246
- DescombesPSchiblerUA liver-enriched transcriptional activator protein, LAP, and a transcriptional inhibitory protein, LIP, are translated from the same mRNACell19916735695791934061
- SebastianTJohnsonPFStop and go: anti-proliferative and mitogenic functions of the transcription factor C/EBPbetaCell Cycle20065995395716687924
- RamjiDPFokaPCCAAT/enhancer-binding proteins: structure, function and regulationBiochem J2002365Pt 356157512006103
- Saint-AuretGDananJLHironMCharacterization of the transcriptional signature of C/EBPbeta isoforms (LAP/LIP) in Hep3B cells: implication of LIP in pro-survival functionsJ Hepatol20115461185119421145827
- LiWKesslerPYegerHA gene expression signature for relapse of primary wilms tumorsCancer Res20056572592260115805255
- OyaMHoriguchiAMizunoRMarumoKMuraiMIncreased activation of CCAAT/enhancer binding protein-beta correlates with the invasiveness of renal cell carcinomaClin Cancer Res2003931021102712631601
- HommaJYamanakaRYajimaNIncreased expression of CCAAT/ enhancer binding protein beta correlates with prognosis in glioma patientsOncol Rep200615359560116465418
- FangTCuiMSunJOrosomucoid 2 inhibits tumor metastasis and is upregulated by CCAAT/enhancer binding protein β in hepatocellular carcinomasOncotarget2015618161061611925965830
- QiuZZouKZhuangLHepatocellular carcinoma cell lines retain the genomic and transcriptomic landscapes of primary human cancersSci Rep201662741127273737
- BuckMTurlerHChojkierMLAP (NF-IL-6), a tissue-specific transcriptional activator, is an inhibitor of hepatoma cell proliferationEMBO J19941348518607906646
- ScrepantiIRomaniLMusianiPLymphoproliferative disorder and imbalanced T-helper response in C/EBP beta-deficient miceEMBO J1995149193219417744000
- GuerzoniCBardiniMMarianiSAInducible activation of CEBPB, a gene negatively regulated by BCR/ABL, inhibits proliferation and promotes differentiation of BCR/ABL-expressing cellsBlood2006107104080408916418324
- ZhuSYoonKSterneckEJohnsonPFSmartRCCCAAT/enhancer binding protein-beta is a mediator of keratinocyte survival and skin tumorigenesis involving oncogenic Ras signalingProc Natl Acad Sci U S A200299120721211756662
- ShimizuYKishimotoTOhtsukaMCCAAT/enhancer binding protein-beta promotes the survival of intravascular rat pancreatic tumor cells via antiapoptotic effectsCancer Sci200798111706171317727681
- GardinerJDAbegglenLMHuangXC/EBPβ-1 promotes transformation and chemoresistance in Ewing sarcoma cellsOncotarget2017816260132602628148901
- ReyaTMorrisonSJClarkeMFWeissmanILStem cells, cancer, and cancer stem cellsNature2001414685910511111689955
- ZahnowCACCAAT/enhancer binding proteins in normal mammary development and breast cancerBreast Cancer Res20024311312112052253
- GrimmSLRosenJMThe role of C/EBPbeta in mammary gland development and breast cancerJ Mammary Gland Biol Neoplasia20038219120414635794
- ZahnowCACCAAT/enhancer-binding protein beta: its role in breast cancer and associations with receptor tyrosine kinasesExpert Rev Mol Med200911e1219351437