
Abstract
Vasculogenic mimicry (VM) describes the process utilized by highly aggressive cancer cells to generate vascular-like structures without the presence of endothelial cells. VM has been vividly described in various tumors and participates in cancer progression dissemination and metastasis. Diverse molecular mechanisms and signaling pathways are involved in VM formation. Furthermore, the patterning characteristics of VM, detected with molecular imaging, are being investigated for use as a tool to aid clinical practice. This review explores the most recent studies investigating the role of VM in tumor induction. Indeed, the recognition of these advances will increasingly affect the development of novel therapeutic target strategies for VM in human cancer.
Introduction
Blood supply contributes to cancer progression, recurrence, and metastasis. With vascular channel structures, nutrients such as glucose could be supplied to tumor tissues; meanwhile, metabolic wastes are evacuated. Previous research has shown that angiogenesis is not an exclusive method to nourish tumor tissues. It has been suggested that cancer stem cells transdifferentiate into endothelial-like cells that support the formation of the extracellular matrix (ECM), eventually inducing tumor-associated neovascularization – this process is called vascular mimicry (VM).Citation1,Citation2 VM represents a new channel that can mimic the embryonic vascular network pattern and provide sufficient blood supply to tumor tissues.Citation3 VM has been depicted in numerous types of malignant tumors, such as glioblastoma, astrocytoma, non-functioning pituitary adenomas, head and neck cancer, lung cancer, breast cancer, esophageal carcinoma, gastric cancer, hepatocellular cancer, colorectal cancer, ovarian carcinoma, prostate cancer, gallbladder cancer, Ewing sarcoma, and osteosarcoma.Citation2,Citation4–Citation11
Over the last few decades, several potential mechanisms of VM formation have been explored, such as epithelial–mesenchymal transition (EMT) and cancer stem cells (CSCs).Citation12 The contribution of several protein factors, including epithelial cell kinase (ephrin type-A receptor 2 [EphA2]), focal adhesion kinase (FAK), hypoxia inducible factor 1-α (HIF-α), integrins laminin 5 (Ln-5) γCitation2 chain, matrix metalloproteinase (MMPs), phosphoinositide 3-kinase (PI3K), and vascular endothelial cadherin (VE-Cad), to VM formation have been investigated.Citation3,Citation13–Citation16 Despite remarkable advances in identifying VM, the key explicit mechanisms of VM are not well characterized, and fully elucidating these mechanisms are of great importance. Lately, advancements in molecular imaging have facilitated the detection, diagnosis, and evaluation of cancer. Qualitative and quantitative investigations on the biological procedures of tumor tissue at the cellular and molecular level can be achieved.
Accordingly, significant features of VM in highly aggressive malignant tumors have been explored widely by various invasive or noninvasive molecular imaging techniques.Citation17 Furthermore, VM formation is associated with unfavorable outcomes of malignant tumor.Citation18 Currently, new drugs and therapeutic strategies targeting VM in cancer are still under development. This review focuses on the advances of concepts, mechanisms, molecular imaging techniques, and targeted therapy in VM that exist at present.
Conceptual progress of VM
Until relatively recently, angiogenesis was considered the sole method of tumor vascularization. Common anti-angiogenesis drugs primarily target endothelial cells, through inducing endothelial cell apoptosis and reducing the proliferation to “starve” tumors. However, the antitumor effects of angiogenesis inhibitors are unsatisfactory. With advances in tumor vascularization, other patterns of blood supply have been detected. In 1999, Maniotis et al first described highly patterned vascular tubes that were formed by highly aggressive uveal melanoma cells without the presence of endothelial cells lining these vessel-like channels.Citation1 Since then, the concept of VM has been solidified as an alternative vascularization method that nourishes tumor tissues.Citation12,Citation19 Recently, choriocarcinoma was shown to utilize VM to increase nutrient retrieval from the blood. In this tumor type, the vessels were surrounded by neoplastic trophoblastic cells. In the marginal regions of tumor tissue, trophoblastic cells invaded host vessels and formed anastomoses.Citation20 VM channels expressed high levels of tissue transglutaminase antigen 2 which enables tubular structure formation.Citation21 CSCs can differentiate into other kinds of cell lineages. Cancer cells in VM express multipotent, stem cell-like phenotypes, including tumor and endothelial phenotypes.Citation22 In melanoma stroma, both fibrovascular septa and VM are observed; VM is distinguished from fibrovascular septa by lamination, thickness, and immunohistochemistry.Citation23
Remarkably, there is indirect evidence suggesting there is circulation of plasma through VM patterns in various types of cancer. Frenkel et al described blood circulation within VM channels with laser scanning confocal angiography in a choroidal melanoma.Citation24 In plastic EW7 Ewing sarcoma tumors in the athymic Tie2-GFP transgenic mouse model, both fluorescent blood vessels and non-fluorescent VM tubes could be observed by intravital microscopy; moreover, the hemodynamics in the cancer-cell-lined channels could be visualized.Citation25 Microstructures of VM by transmission electron microscopy illustrated empty spaces in the central region of the tubes; there were no internal vacuoles with organelle-poor cytoplasm, and desmosomes contributed to the establishment of the junction between VM and endothelial-like cells.Citation26
Additionally, other characteristics of VM have been recently described. In a heterotopic malignant mesothelioma xenograft model in BALB/C node mice, a small number of vessels consisting of mouse endothelial cells were observed; however, further immunohistochemical (IHC) analysis using the human-specific mitochondria antibody MAB1273B revealed VM channels containing erythrocytes at the periphery of the tumor tissue.Citation27 In multiple myeloma, Nico et al showed that mast cells contribute to the formation of VM channels.Citation28 Co-culture of aggressive melanoma cell lines with mesenchymal stromal cells were shown to acquire endothelial cell-like properties, eventually forming VM channels; however, such a phenomenon was not detected in poorly aggressive melanoma cell lines.Citation29 These data suggest that VM is a novel method for providing blood and nutrition to aggressive tumor tissues.
The composition of tumor cells and a basement membrane made VM different from the classical concept of angiogenesis. To date, VM can be characterized in tumor samples with IHC using a positive staining pattern of both CD31 and periodic acid–Schiff (PAS) as markers. Two distinctive types of VM in aggressive malignant tumors have been identified. Type 1, known as the tubular type, is composed of non-endothelial cell-lined blood tubes, and cancer cells were found lining the surface of the channels.Citation30 Type 2, known as the patterned matrix type, comprises a basement membrane rich in fibronectin, collagens, and laminin, where the membrane is surrounded by tumor cells instead of endothelial cells.Citation31
Furthermore, VM is used as a means to support tumor growth in the early stages of tumorigenesis. Both angiogenesis and VM are coordinately used in tumor tissues.Citation10 In the VM–angiogenesis junction, VM serves as part of the functional microcirculation; cancer cells within the tumor-lined vascular channels can easily transfer into endothelial-lined blood vessels, thereby facilitating tumor invasion and metastasis.
Mechanisms of VM activating invasion and metastasis
VM plays an essential role in the process of tumor invasion and metastasis. After the initiation of VM, a mosaic vessel made of tumor cells and endothelial cells gradually forms. Vessels connect and merge with host vessels, and cancer cells could obtain oxygen and nutriment via VM – consequently inducing cancer growth ().Citation32 Moreover, VM contributes to portal vein invasion in hepatocellular carcinoma through the expression of several proteins, including Notch1, MMP-2, and MMP-9.Citation33 In head and neck squamous cell cancer, tumor cells express both VM and endothelial-specific markers and respond to vascular endothelial growth factor (VEGF) and endostatin. These features can be enhanced by transforming growth factor-β1, facilitating the acquisition of an endothelial cell type in a subpopulation of cells. Additionally, tumor cell mobility and invasiveness were heightened.Citation34 In non–small cell lung cancer, Dickkopf-1 significantly induces VM formation and promotes cancer cell growth, migration, and metastasis via overexpression of EMT- and CSC-associated proteins.Citation35 Maspin has a positive correlation with VM; deregulated maspin facilitates tumor cell invasion and metastasis in non–small cell lung cancer.Citation36 In melanoma cells, hypoxia induces the release of mitochondrial reactive oxygen species and promotes activation of the Met proto-oncogene, thereby enhancing VM formation and resulting in tumor cell spread, increased motility, invasion, and metastasis.Citation37 In the hypoxic microenvironment, HIF-1α induces VM formation via the upregulation of lysyl oxidases, such as in hepatocellular cancer, and promotes tumor cell metastasis and progression.Citation38 Overexpression of miRNA-124 represses VM formation via inhibition of MMP-2, MMP-9, and VEGF; further analysis indicated that miRNA-124 is important in preventing tumor cell migration.Citation39 Witkiewicz et al demonstrated that VM resembled lymphatic vessels or veins in morphology; moreover, endothelial cells were replaced by fibroblasts, thereby facilitating tumor metastasis.Citation40 In esophageal squamous cell carcinoma, researchers used RNA silencing, and observed a positive correlation between VM channels and N-cadherin expression.Citation41 Genes involved in metastatic and invasive behavior could be influenced by the tumor microenvironment. On being exposed to a laminin-rich niche, highly invasive uveal melanoma cells can form VM channels; however, a downregulation of genes including CD44, thrombospondin 1, and cyclin E2 were observed, thereby reducing melanoma cell invasion.Citation42
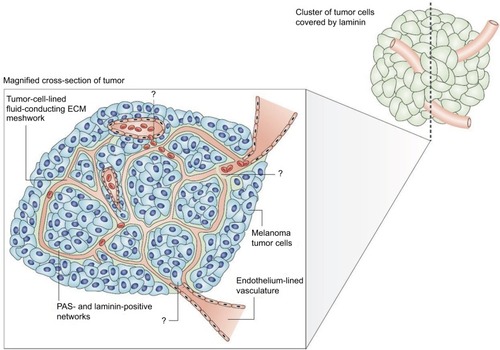
Figure 1 The melanoma fluid-conducting extracellular matrix.
Notes: In mice with aggressive melanoma (blue cells). The endothelium-lined vessels (pink) are closely opposed to the fluid-conducting meshwork formed by tumor cells. Tumor cells can remodel the vasculature, which becomes leaky and leads to the extravascular accumulation of erythrocytes and plasma (red). Reprinted by permission from Springer Nature, Nature Reviews Cancer, Vasculogenic mimicry and tumour-cell plasticity: lessons from melanoma, Hendrix MJ, Seftor EA, Hess AR, Seftor RE, COPYRIGHT 2003.Citation77 The specific mechanism of VM are still under investigation.
Research of VM using molecular imaging
The development of noninvasive imaging is essential for cancer detection and diagnosis during the early stages of the disease. Molecular imaging provides a better way to understand the biologic processes governing VM at a cellular and molecular level in living organisms. This use of this technique has become more applicable for cancer in the past few decades. VM can develop vascular anastomosis with microcirculation in the tumor tissues. Theoretically, contrasting agents can easily enter the VM tube, thereby enabling visualization of imaging patterns. This technique brings hope to a variety of imaging techniques, especially the molecular imaging technology.
Interestingly, evidence of VM has been reported in human tumors by using novel molecular imaging technologies. In a study that included 18 patients diagnosed with choroidal melanomas, patients were injected with indocyanine green as a contrasting agent prior to being imaged with a confocal scanning laser ophthalmoscope. The results demonstrated microvasculature patterns within the tumor; the tubular structures correlated well with the histopathological appearance of VM.Citation43 Frenkel et al conducted indocyanine green laser scanning confocal angiography on patients with posterior choroidal melanoma. They found that fluid enters VM by leakage instead of through endothelial cell-lined vessels; furthermore, fluid accumulated in VM was never characterized as a stagnant pool.Citation24 Polylactic acid – a nanoparticle agent – is showing promising advantages for targeted ultrasound imaging. This nanoparticle could be conjugated to the anti-human epidermal growth factor receptor 2 (HER2) antibody, thereby allowing for HER-positive breast tumor cells to be specifically bound.Citation44 In accordance with this finding, imaging of VM channels by high-resolution ultrasound using targeted nanoparticles may be explored. With the development of intravascular macromolecular contrasting agents for magnetic resonance imaging (MRI), the diagnosis of VM tubes by techniques such as time-course dynamic micro-magnetic resonance angiography analysis could be achieved. Shirakawa et al used this technology to investigate the hemodynamics of VM in a newly established human inflammatory breast cancer xenograft in BALA/c nude mice. The results revealed that tumor center exhibited a signal that gradually increased in intensity, which is consistent with histological features of VM in the same area. The group also observed the presence of a VM-angiogenesis junction by transmission electron microscopy and IHC.Citation45 Similar results were observed in Kobayashi et al’s study, and the data also suggested VM is involved in tumor tissue perfusion.Citation46 Yamamoto et al described the radiological features of VM in malignant gliomas using the newly developed MRI. Anaplastic oligodendroglioma illustrating hyperintensity on fluid-attenuated inversion recovery images as well as histological diagnosis proved these were VM.Citation17 Thus, due to remarkable progress made in this endeavor, imaging features of VM should be further explored as a tool for clinical application.
Significance of VM expression in clinical practice
Identification of VM formation is of paramount importance to daily clinical practice. Previous studies have attempted to measure microvascular density to quantify the degree of angiogenesis and its relationship to prognosis; however, the results are rather controversial.Citation47,Citation48 Part of the rationale is that the tumor microcirculation is lined not only by endothelial cells but also with tumor cells; VM contributes to the intratumor heterogeneity in aggressive cancer. Moreover, standard VM assessment is based on the quantification of positive PAS and negative CD31 staining of vessel-like structures; this is different from the commonly used tumoral microvascular density assessment. In human gliomas, VM formation is highly aggressive and acts as a complementary strategy for providing tumor tissues with blood supply in poorly vascularized areas.Citation49 In prostatic cancer, the correlation between VM and histologic grading was insignificant, and the correlation between VM and perineurial invasion was rather weak. This observation may be due to the composition of prostatic tissue, which contains an abundance of smooth muscle fibers, as well as the staining methods used.Citation50
VM has been demonstrated as an unfavorable survival factor and a marker of poor prognosis in various cancers. In colorectal carcinoma, VM was shown to be a strong, independent, prognostic factor of survival.Citation7 A meta-analysis by Cao et al comprising 15 types of malignant tumors revealed that patients with VM-positive cancer showed a less favorable 5-year overall survival than patients with VM-negative cancer, especially in advanced-stage cancer.Citation18 In patients with non–small cell lung cancer, CD133 expression and VM formation were significantly high and associated with tumor differentiation, lymph node metastasis, clinical stage as well as prognosis; therefore, VM could be used as a prognostic marker in clinical practice.Citation51 Indeed, VM could also be used as a prognostic marker in ovarian cancer, inflammatory breast cancer, and gastric adenocarcinoma.Citation52–Citation54 In hepatocellular carcinoma, VM promotes cancer progression after orthotopic liver explant.Citation55 Data on laryngeal squamous cell carcinoma suggested VM was a poor prognostic factor of disease-specific and metastasis-free survival; further analysis revealed that VM contributed to tumor progression through promoting lymph node metastasis.Citation56 In uveal melanoma samples, Chromosome 3 aberrations played an important role in the formation of VM networks; identification of Chromosome 3 with fine needle aspiration could be used in prognostic assessment.Citation57 There is a strong correlation between increased expression of HER2 and the positive rate of VM in invasive breast cancer. Furthermore, VM was associated with a positive nodal status and advanced clinical stages.Citation58 However, other studies suggested different outcomes. Massi et al indicated that, in patients with pT3 and pT4 cutaneous melanoma, there is no evidence of VM as a prognostic factor.Citation59
In some biopsy samples of malignant cancer or benign diseases, clinical evidence of VM is detected. VM tubes that contained red blood cells were found in malignant mesothelioma, cerebral cavernous malformations, and anaplastic oligodendroglioma.Citation17,Citation27 In benign nevi, IHC analysis of VM was shown in a minority of cases as compared to that of melanomas and was associated with unfavorable survival.Citation60 In a patient diagnosed with multiple probable cerebral cavernous malformations, a slightly enlarged hyperintense region on the right frontal lobe persisted after therapy. Postoperative histological diagnosis demonstrated an anaplastic oligodendroglioma and cerebral cavernous malformations with VM.Citation17 Therefore, early identification of radiological features of VM is essential for clinical decision making.
Blood supply plays an important role in tumor growth and metastasis; however, the therapeutic efficiency of drugs directed toward endothelial cells is unsatisfactory.Citation61 The discovery of VM in tumor blood supply provided new insights in cancer biology. In Merkel cell carcinoma, VM-rich regions are illustrated as resistant to conventional chemotherapeutic agents.Citation62 VM channels decrease cancer latency and increase intratumoral cisplatin delivery but may also reduce drug efficacy.Citation8 Therefore, VM may be considered a useful target for treating cancer.
Therapeutic targeting of VM
Present treatment strategies such as chemotherapy are inefficient in aggressive cancer. Residual tumor cells may form VM channels, thereby providing oxygen and nutrients, which support cell proliferation and cancer progression. Thus, VM is a promising target for developing novel anticancer therapeutics, and many drugs have been investigated. Angiogenesis inhibitors including anginex, TNP-470, and endostatin were delivered to both human melanoma cell lines and human endothelial cell lines in vitro; marked inhibition on the vascular cord and tube formation were observed in endothelial cells as compared to melanoma cells. Detailed analysis showed higher mRNA and protein levels for two putative endostatin receptors – α5 integrin and heparin sulfate proteoglycan 2 – in endothelial cells compared to melanoma cells.Citation63 The Rho kinase inhibitor fasudil induces VM destruction in B 16 melanoma cell xenograft.Citation64 Fasudil blocks the RhoA/ROCK signaling pathway and VM formation is inhibited.Citation11 Nicotinamide can effectively inhibit the formation of VM channels as well as destroy preexisting channels by downregulating VE-Cad expression. VM formation was suppressed for up to a month after exposure to nicotinamide. Although melanoma cell proliferation was significantly inhibited, it is worth noting the opposite effect of increased invasion capacity of the tumor cells by nicotinamide.Citation65 When cultured on an aggressive tumor cell-preconditioned three-dimensional matrix, VM formation was induced in poorly aggressive tumor cells. Chemically modified tetracycline has been reported to be capable of targeting the tumor microenvironment by inhibition of MMP activity. Generation of Laminin-5-γ2 (Ln-5γ2) chain promigratory fragments is suppressed, thereby blocking its gene expression of poorly aggressive melanoma cells; as a consequence, VM formation is inhibited.Citation66 The inhibition of VM by verteporfin is due to the suppression of MMP-2 and VE-Cad in human pancreatic ductal adenocarcinoma cell lines.Citation67 Nanostructured functional drug-loaded liposomes, modified with an HIV peptide lipid-derivative conjugate, containing epirubicin and celecoxib, DSPE-PEG2000-PTDHIV-1, downregulated VM expression via the inhibition of VE-Cad, FAK, EphA2, HIF-1α, and MMP-9 in invasive breast cancer xenografts in nude mice.Citation68 Paclitaxel-loaded liposomes modified with tandem peptides were observed to induce the destruction of VM channels in glioma cells.Citation69 Doxycycline downregulated the expression of VE-Cad and MMPs and inhibited VM in hepatocellular carcinoma.Citation70 In clear-cell renal cell carcinoma, the tumor was efficiently targeted by sunitinib – both in vitro and in vivo; however, resistant tumor cells emerged with a more aggressive phenotype and high level of vascularization during maintenance treatment. VM formation, autonomous pro-tumoral gene expression, a stroma-rich variant, and a favorable tumor microenvironment are the main reasons for this change. Second-line therapy with everolimus enhanced the inhibition of sunitinib-resistant tumors by suppressing VM channels.Citation71 In a malignant glioma tumor model, the bispecific immunotoxin VEGF165-ephrin A1-PE38KDEL was delivered by human mesenchymal stem cells. As a result, VM formation and tumor growth was effectively inhibited; this provided a novel strategy for treatment of malignant gliomas.Citation72 In melanoma cells, cilengitide – an inhibitor of αν integrins – reduced VM formation by decreasing ECM invasion as well as VEGF-A and MMP-9 secretion.Citation73 Similarly, genistein was shown to inhibit VM by downregulation of VE-Cad in uveal melanoma cells.Citation74 Thalidomide is efficient in inhibiting VM channels and induces tumor cell necrosis by regulation of the NF-κB signaling pathway.Citation75 Antioxidants such as resveratrol can decrease the level of VEGF and cas-pase-3 – thus, minimizing capillary formation.Citation76
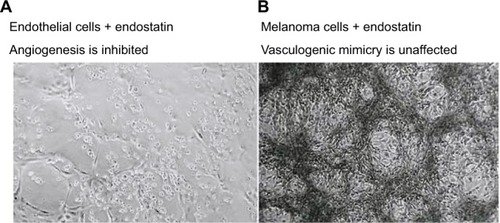
The effects of antiangiogenic compounds on VM formation were also assessed. The inhibitory effects of Endostar – a recombinant human endostatin – on VM formation of melanoma cells () and glioblastoma cells were shown to be insignificant.Citation9,Citation77 In an ovarian cancer model, short-term bevacizumab treatment demonstrated antitumor effects; however, it also contributed to cancer progression due to increased HIF-1α expression and VM formation.Citation78 Apropos the above, angiogenesis inhibitors have failed to prevent progression and growth of malignant tumors, which also demonstrates that VM formation – as a form of microcirculation – plays an important role in tumor progression.
Figure 2 The therapeutic effects of endostatin (an angiogenesis inhibitor) on microvascular endothelial cells (A), lack of vessel networks) compared with melanoma cells (B), VM formation was unaffected) in three-dimensional gels of collagen I in vitro.
Note: Reprinted by permission from Springer Nature, Nature Reviews Cancer, Vasculogenic mimicry and tumour-cell plasticity: lessons from melanoma, Hendrix MJ, Seftor EA, Hess AR, Seftor RE, COPYRIGHT 2003.Citation77
miRNAs are small non-coding RNA molecules, which contain approximately 22 nucleotides, that promote post-transciptional gene silencing.Citation79 Accordingly, miRNAs play a crucial role in modulating cancer cell migration, invasion, and tumor angiogenesis. miRNA-9 is a tissue-specific miRNA localized in tissues of the central nervous system. In glioma cell lines, miRNA-9 inhibits VM formation and tumor growth by controlling Stathmin expression.Citation80 A study by Wan et al revealed miRNA-124 inhibits VM formation of cervical cancer cells by targeting AmotL1 and suppressing the EMT process.Citation39 As noted earlier, the VE-Cad complex is involved in tumor VM formation. miRNA-27b acts as an inhibitor of VM in ovarian cancer cells by suppressing VE-Cad expression.Citation81 The full spectrum of mechanisms of miRNAs to tumor VM inhibition remains to be elucidated.
Over recent years, traditional Chinese medicines have been investigated in treating various cancers. Lycorine hydrochloride is effective against capillary-like tube formations in a melanoma cell line model by hindering VE-Cad expression.Citation82 Dehydroeffusol was found to significantly inhibit VM formation by downregulating MMP-2.Citation83 Other drugs, including grape-seed proanthocyanidins, isoxanthohumol, ginsenoside Rg3, norcantharidin, paris polyphylla, and curcumin have been well characterized and possess antitumor effects ().Citation84–Citation89
Table 1 Summary of traditional Chinese medicine in cancer treatment by targeting VM
Conclusion and future vision
Although initially described in aggressive uveal melanoma, VM has led to profound changes in our understanding of tumor nourishment in a variety of cancers. The phenomenon illustrates that tumor cells can transdifferentiate due to a multipotent, CSC-like phenotype and generate ECM-rich, negative CD31, and positive PAS vascular networks. These channels are distinct microvessels that differ from host vessels and provide adequate blood supply which promotes tumor growth. As such, VM plays an essential role in facilitating tumor invasion and metastasis. Realizing its importance in tumorigenesis, researchers have been investigating VM with the aid of molecular imaging and, thus far, the results have been promising. Furthermore, VM was considered an independent, unfavorable, prognostic factor of clinical practice; VM is associated with aggressive behavior and easily metastasizes to distant sites. To date, anti-angiogenic therapies focus primarily on tumor blood vessels that are formed by endothelial cells, VM formation is one of the reasons for the limited therapeutic effects of existing anticancer drugs; although enormous efforts have been made in an attempt to eliminate VM formation and some drugs demonstrate a promising result, research into treatment strategies to counter VM is still ongoing. Much remains to be learned to delineate the role of VM in tumor progression on the cellular level and to translate these findings into novel therapies that target VM in the treatment of cancer.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (grant nos. 81372436 and 81773230); the Science and Technology Innovative Research Group of Zhengzhou city (grant no. 121PCXTD524), and the Joint Research Funds of Henan province and the Minister of Health of China (grant no. 201201009). The authors would like to thank Kyle Vaugh Laster (Hormel Cancer Institute, USA) for editing the language of the present manuscript.
Author contributions
Both HL and HG have contributed to the conception and design, data acquisition, and interpretation; drafting the article and final approval of the final version to be submitted; and agree to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of the work are appropriately investigated and resolved.
Disclosure
The authors report no conflicts of interest in this work.
References
- ManiotisAJFolbergRHessAVascular channel formation by human melanoma cells in vivo and in vitro: vasculogenic mimicryAm J Pathol1999155373975210487832
- Ricci-VitianiLPalliniRBiffoniMTumour vascularization via endothelial differentiation of glioblastoma stem-like cellsNature2010468732582482821102434
- SeftorRESeftorEAKoshikawaNCooperative interactions of laminin 5 gamma2 chain, matrix metalloproteinase-2, and membrane type-1-matrix/metalloproteinase are required for mimicry of embryonic vasculogenesis by aggressive melanomaCancer Res200161176322632711522618
- Di MicheleJRotondoFKovacsKVasculogenic mimicry in clinically non-functioning pituitary adenomas: a histologic studyPathol Oncol Res201723480380928084580
- SunWFanYZZhangWZGeCYA pilot histomorphology and hemodynamic of vasculogenic mimicry in gallbladder carcinomas in vivo and in vitroJ Exp Clin Cancer Res2011304621529356
- SharmaNSeftorRESeftorEAProstatic tumor cell plasticity involves cooperative interactions of distinct phenotypic subpopulations: role in vasculogenic mimicryProstate200250318920111813211
- BaetenCIHillenFPauwelsPde BruineAPBaetenCGPrognostic role of vasculogenic mimicry in colorectal cancerDis Colon Rectum200952122028203519934926
- WilliamsonSCMetcalfRLTrapaniFVasculogenic mimicry in small cell lung cancerNat Commun201671332227827359
- LiuZLiYZhaoWMaYYangXDemonstration of vasculogenic mimicry in astrocytomas and effects of Endostar on U251 cellsPathol Res Pract20112071064565121940112
- UpileTJerjesWRadhiHVascular mimicry in cultured head and neck tumour cell linesHead Neck Oncol201135522196216
- XiaYCaiXFanJRhoA/ROCK pathway inhibition by fasudil suppresses the vasculogenic mimicry of U2OS osteosarcoma cells in vitroAnticancer Drugs201728551452128225457
- LiuQQiaoLLiangNThe relationship between vasculogenic mimicry and epithelial-mesenchymal transitionsJ Cell Mol Med20162091761176927027258
- HessARSeftorEAGrumanLMKinchMSSeftorREHendrixMJVE-cadherin regulates EphA2 in aggressive melanoma cells through a novel signaling pathway: implications for vasculogenic mimicryCancer Biol Ther20065222823316481735
- LiuKSunBZhaoXHypoxia induced epithelial-mesenchymal transition and vasculogenic mimicry formation by promoting Bcl-2/Twist1 cooperationExp Mol Pathol201599238339126318343
- ZangMZhangYZhangBCEACAM6 promotes tumor angiogenesis and vasculogenic mimicry in gastric cancer via FAK signalingBiochim Biophys Acta2015185251020102825703140
- HessARSeftorEASeftorREHendrixMJPhosphoinositide 3-kinase regulates membrane Type 1-matrix metalloproteinase (MMP) and MMP-2 activity during melanoma cell vasculogenic mimicryCancer Res200363164757476212941789
- YamamotoJShimajiriSMiyaokaRNishizawaSPitfalls of conservative treatments of multiple probable cerebral cavernous malformations (CCMs): clinicopathological features of CCMs coexisting with vasculogenic mimicry in an anaplastic oligodendrogliomaBrain Tumor Pathol201431321522124337536
- CaoZBaoMMieleLSarkarFHWangZZhouQTumour vasculogenic mimicry is associated with poor prognosis of human cancer patients: a systemic review and meta-analysisEur J Cancer201349183914392323992642
- LiuZSunBQiLLiHGaoJLengXZinc finger E-box binding homeobox 1 promotes vasculogenic mimicry in colorectal cancer through induction of epithelial-to-mesenchymal transitionCancer Sci2012103481382022212097
- ShihIeMTrophoblastic vasculogenic mimicry in gestational choriocarcinomaMod Pathol201124564665221217646
- JonesRAWangZDookieSGriffinMThe role of TG2 in ECV304-related vasculogenic mimicryAmino Acids20134418910122231926
- Valyi-NagyKKormosBAliMShuklaDValyi-NagyTStem cell marker CD271 is expressed by vasculogenic mimicry-forming uveal melanoma cells in three-dimensional culturesMol Vis20121858859222419851
- LinAYManiotisAJValyi-NagyKDistinguishing fibrovascular septa from vasculogenic mimicry patternsArch Pathol Lab Med2005129788489215974811
- FrenkelSBarzelILevyJDemonstrating circulation in vasculogenic mimicry patterns of uveal melanoma by confocal indocyanine green angiographyEye (Lond)200822794895217363922
- HillenFKaijzelELCastermansKoude EgbrinkMGLöwikCWGriffioenAWA transgenic Tie2-GFP athymic mouse model; a tool for vascular biology in xenograft tumorsBiochem Biophys Res Commun2008368236436718237547
- ClementeMPérez-AlenzaMDIlleraJCPeñaLHistological, immunohistological, and ultrastructural description of vasculogenic mimicry in canine mammary cancerVet Pathol201047226527420106772
- PulfordEHockingAGriggsKVasculogenic mimicry in malignant mesothelioma: an experimental and immunohistochemical analysisPathology201648765065927956272
- NicoBMangieriDCrivellatoEVaccaARibattiDMast cells contribute to vasculogenic mimicry in multiple myelomaStem Cells Dev2008171192218205547
- VartanianAKarshievaSDombrovskyVBelyavskyAMelanoma educates mesenchymal stromal cells towards vasculogenic mimicryOncol Lett20161164264426827313776
- El HallaniSBoisselierBPeglionFA new alternative mechanism in glioblastoma vascularization: tubular vasculogenic mimicryBrain2010133Pt 497398220375132
- LiuWBXuGLJiaWDPrognostic significance and mechanisms of patterned matrix vasculogenic mimicry in hepatocellular carcinomaMed Oncol201128Suppl 1S228S23820957524
- SeftorREHessARSeftorEATumor cell vasculogenic mimicry: from controversy to therapeutic promiseAm J Pathol201218141115112522944600
- JueCZhifengWZhishengZVasculogenic mimicry in hepatocellular carcinoma contributes to portal vein invasionOncotarget2016747779877799727793002
- TongMHanBBHolpuchASPeiPHeLMallerySRInherent phenotypic plasticity facilitates progression of head and neck cancer: endotheliod characteristics enable angiogenesis and invasionExp Cell Res201331971028104223370231
- YaoLZhangDZhaoXDickkopf-1-promoted vasculogenic mimicry in non-small cell lung cancer is associated with EMT and development of a cancer stem-like cell phenotypeJ Cell Mol Med20162091673168527240974
- WuSYuLChengZSongWZhouLTaoYExpression of maspin in non-small cell lung cancer and its relationship to vasculogenic mimicryJ Huazhong Univ Sci Technolog Med Sci201232334635222684556
- ComitoGCalvaniMGiannoniEHIF-1α stabilization by mitochondrial ROS promotes Met-dependent invasive growth and vasculogenic mimicry in melanoma cellsFree Radic Biol Med201151489390421703345
- WangMZhaoXZhuDHIF-1α promoted vasculogenic mimicry formation in hepatocellular carcinoma through LOXL2 up-regulation in hypoxic tumor microenvironmentJ Exp Clin Cancer Res20173616028449718
- WanHYLiQQZhangYMiR-124 represses vasculogenic mimicry and cell motility by targeting amotL1 in cervical cancer cellsCancer Lett2014355114815825218344
- WitkiewiczHOhPSchnitzerJEII. Capsular vaso-mimicry formed by transgenic mammary tumor spheroids implanted ectopically into mouse dorsal skin fold: implications for cellular mechanisms of metastasisF1000Res20132924555024
- WangFLiXKXuHYN-cadherin participated in invasion and metastasis of human esophageal squamous cell carcinoma via taking part in the formation of vasculogenic mimicryMed Oncol201532248025575439
- FolbergRArbievaZMosesJTumor cell plasticity in uveal melanoma: microenvironment directed dampening of the invasive and metastatic genotype and phenotype accompanies the generation of vasculogenic mimicry patternsAm J Pathol200616941376138917003493
- MuellerAJBartschDUFolbergRImaging the microvasculature of choroidal melanomas with confocal indocyanine green scanning laser ophthalmoscopyArch Ophthalmol1998116131399445206
- LiuJLiJRosolTJPanXVoorheesJLBiodegradable nanoparticles for targeted ultrasound imaging of breast cancer cells in vitroPhys Med Biol200752164739474717671332
- ShirakawaKKobayashiHSobajimaJHashimotoDShimizuAWakasugiHInflammatory breast cancer: vasculogenic mimicry and its hemodynamics of an inflammatory breast cancer xenograft modelBreast Cancer Res20035313613912793894
- KobayashiHShirakawaKKawamotoSRapid accumulation and internalization of radiolabeled herceptin in an inflammatory breast cancer xenograft with vasculogenic mimicry predicted by the contrast-enhanced dynamic MRI with the macromolecular contrast agent G6-(1B4M-Gd)(256)Cancer Res200262386086611830544
- ShinHJKimEKMoonHJYoonJHHanKHKwakJYCan increased tumoral vascularity be a quantitative predicting factor of lymph node metastasis in papillary thyroid microcarcinoma?Endocrine201447127328224366642
- IakovlevVVGabrilMDubinskiWMicrovascular density as an independent predictor of clinical outcome in renal cell carcinoma: an automated image analysis studyLab Invest2012921465622042086
- LiuXMZhangQPMuYGClinical significance of vasculogenic mimicry in human gliomasJ Neurooncol2011105217317921533525
- AhmadiSAMoinfarMGohari MoghaddamKBahadoriMPractical application of angiogenesis and vasculogenic mimicry in prostatic adenocarcinomaArch Iran Med201013649850321039005
- WuSYuLWangDAberrant expression of CD133 in non-small cell lung cancer and its relationship to vasculogenic mimicryBMC Cancer20121253523170850
- LiangJYangBCaoQWuXAssociation of vasculogenic mimicry formation and CD133 expression with poor prognosis in ovarian cancerGynecol Obstet Invest201681652953627160772
- ShirakawaKWakasugiHHeikeYVasculogenic mimicry and pseudo-comedo formation in breast cancerInt J Cancer200299682182812115483
- LiMGuYZhangZVasculogenic mimicry: a new prognostic sign of gastric adenocarcinomaPathol Oncol Res201016225926620016961
- GuzmanGCotlerSJLinAYManiotisAJFolbergRA pilot study of vasculogenic mimicry immunohistochemical expression in hepatocellular carcinomaArch Pathol Lab Med2007131121776178118081435
- WangWLinPHanCCaiWZhaoXSunBVasculogenic mimicry contributes to lymph node metastasis of laryngeal squamous cell carcinomaJ Exp Clin Cancer Res2010296020525189
- MeirTZeschnigkMMasshöferLPe’erJChowersIThe spatial distribution of monosomy 3 and network vasculogenic mimicry patterns in uveal melanomaInvest Ophthalmol Vis Sci20074851918192217460242
- LiuTSunBZhaoXHER2/neu expression correlates with vasculogenic mimicry in invasive breast carcinomaJ Cell Mol Med201317111612223279650
- MassiDFranchiAPaglieraniMVasculogenic mimicry has no prognostic significance in pT3 and pT4 cutaneous melanomaHum Pathol200435449650215116332
- SpiliopoulosKPeschosDBatistatouAImmunohistochemical study of vasculogenic mimicry and angiogenesis in melanocytic tumors of the eye and the periocular areaAnticancer Res20173731113112028314272
- PotenteMGerhardtHCarmelietPBasic and therapeutic aspects of angiogenesisCell2011146687388721925313
- LezcanoCKleffelSLeeNMerkel cell carcinoma expresses vasculogenic mimicry: demonstration in patients and experimental manipulation in xenograftsLab Invest201494101092110225111691
- van der SchaftDWSeftorRESeftorEAEffects of angiogenesis inhibitors on vascular network formation by human endothelial and melanoma cellsJ Natl Cancer Inst200496191473147715467037
- XiaYCaiXYFanJQRho kinase inhibitor fasudil suppresses the vasculogenic mimicry of B16 mouse melanoma cells both in vitro and in vivoMol Cancer Ther20151471582159025934709
- ItzhakiOGreenbergEShalmonBNicotinamide inhibits vasculogenic mimicry, an alternative vascularization pathway observed in highly aggressive melanomaPLoS One201382e5716023451174
- SeftorRESeftorEAKirschmannDAHendrixMJTargeting the tumor microenvironment with chemically modified tetracyclines: inhibition of laminin 5 gamma2 chain promigratory fragments and vasculogenic mimicryMol Cancer Ther20021131173117912479698
- WeiHWangFWangYVerteporfin suppresses cell survival, angiogenesis and vasculogenic mimicry of pancreatic ductal adenocarcinoma via disrupting the YAP-TEAD complexCancer Sci2017108347848728002618
- JuRJLiXTShiJFLiposomes, modified with PTD(HIV-1) peptide, containing epirubicin and celecoxib, to target vasculogenic mimicry channels in invasive breast cancerBiomaterials201435267610762124912818
- LiuYMeiLYuQMultifunctional tandem peptide modified paclitaxel-loaded liposomes for the treatment of vasculogenic mimicry and cancer stem cells in malignant gliomaACS Appl Mater Interfaces2015730167921680126173814
- MengJSunBZhaoXDoxycycline as an inhibitor of the epithelial-to-mesenchymal transition and vasculogenic mimicry in hepatocellular carcinomaMol Cancer Ther201413123107312225277383
- SerovaMTijeras-RaballandADos SantosCEverolimus affects vasculogenic mimicry in renal carcinoma resistant to sunitinibOncotarget2016725384673848627509260
- ZhangYSunXHuangMKeYWangJLiuXA novel bispecific immunotoxin delivered by human bone marrow-derived mesenchymal stem cells to target blood vessels and vasculogenic mimicry of malignant gliomasDrug Des Devel Ther2015929472959
- RuffiniFGrazianiGLevatiLTentoriLD’AtriSLacalPMCilengitide downmodulates invasiveness and vasculogenic mimicry of neuropilin 1 expressing melanoma cells through the inhibition of αvβ5 integrinInt J Cancer20151366E545E55825284767
- CongRSunQYangLGuHZengYWangBEffect of Genistein on vasculogenic mimicry formation by human uveal melanoma cellsJ Exp Clin Cancer Res20092812419735546
- ZhangSLiMGuYThalidomide influences growth and vasculogenic mimicry channel formation in melanomaJ Exp Clin Cancer Res2008276018983651
- VartanianAABurovaOSStepanovaEVBaryshnikovAYLichinitserMRMelanoma vasculogenic mimicry is strongly related to reactive oxygen species levelMelanoma Res200717637037917992120
- HendrixMJSeftorEAHessARSeftorREVasculogenic mimicry and tumour-cell plasticity: lessons from melanomaNat Rev Cancer20033641142112778131
- XuYLiQLiXYYangQYXuWWLiuGLShort-term anti-vascular endothelial growth factor treatment elicits vasculogenic mimicry formation of tumors to accelerate metastasisJ Exp Clin Cancer Res2012311622357313
- BartelDPMicroRNAs: genomics, biogenesis, mechanism, and functionCell2004116228129714744438
- SongYMuLHanXMicroRNA-9 inhibits vasculogenic mimicry of glioma cell lines by suppressing Stathmin expressionJ Neurooncol2013115338139024043603
- LiuWLvCZhangBZhouQCaoZMicroRNA-27b functions as a new inhibitor of ovarian cancer-mediated vasculogenic mimicry through suppression of VE-cadherin expressionRNA20172371019102728396577
- LiuRCaoZTuJLycorine hydrochloride inhibits metastatic melanoma cell-dominant vasculogenic mimicryPigment Cell Melanoma Res201225563063822781316
- LiuWMengMZhangBDehydroeffusol effectively inhibits human gastric cancer cell-mediated vasculogenic mimicry with low toxicityToxicol Appl Pharmacol201528729811025982451
- LuanYYLiuZMZhongJYYaoRYYuHSEffect of grape seed pro-anthocyanidins on tumor vasculogenic mimicry in human triple-negative breast cancer cellsAsian Pac J Cancer Prev201516253153525684483
- SerweARudolphKAnkeTErkelGInhibition of TGF-β signaling, vasculogenic mimicry and proinflammatory gene expression by isoxanthohumolInvest New Drugs201230389891521340508
- GuoJQZhengQHChenHGinsenoside Rg3 inhibition of vasculogenic mimicry in pancreatic cancer through downregulation of VE-cadherin/EphA2/MMP9/MMP2 expressionInt J Oncol20144531065107224938458
- ZhangJTSunWZhangWZNorcantharidin inhibits tumor growth and vasculogenic mimicry of human gallbladder carcinomas by suppression of the PI3-K/MMPs/Ln-5γ2 signaling pathwayBMC Cancer20141419324628713
- YaoNRenKWangYParis polyphylla suppresses proliferation and vasculogenic mimicry of human osteosarcoma cells and inhibits tumor growth in vivoAm J Chin Med201745357559828385078
- ChiablaemKLirdprapamongkolKKeeratichamroenSSuraritRSvastiJCurcumin suppresses vasculogenic mimicry capacity of hepatocellular carcinoma cells through STAT3 and PI3K/AKT inhibitionAnticancer Res20143441857186424692720