
Abstract
Background
In patients with gastric cancer, one of the greatest obstacles to effective chemotherapy is the development of chemoresistance. It has been previously reported that hypoxia-inducible factor-1 alpha (HIF-1α) is associated with acquisition of chemoresistance, and more recent studies have also noted an association of pyruvate kinase muscle 1 (PKM1) and chemoresistance. The purpose of this study was to identify the effect of HIF-1α and PKM1 expression on the development of acquired chemoresistance using a paclitaxel (PTX)-resistant gastric cancer cell line.
Materials and methods
A cancer cell line resistant to PTX was established from MKN45 cells by stepwise exposure to drug (rMKN45-PTX). The expressions of HIF-1α, apoptosis, vascular endothelial growth factor (VEGF), multidrug transporters and glycolytic enzyme were examined by Western blotting, enzyme-linked immunosorbent assay and immunohistochemistry. We also assessed the tumor proliferation by subcutaneous tumor and peritoneal dissemination of mouse xenograft model.
Results
The resistance index was 6.1 by determining as the ratio of the 50% growth inhibition (IC50) of rMKN45-PTX/IC50 of MKN45. Expression of nuclear factor kappa B and HIF-1α was increased in rMKN45-PTX cells compared with the parent cells. Expression of Bax and caspase-3 was significantly downregulated, whereas expression of Bcl-xL, P-glycoprotein, multidrug resistance-associated protein and VEGF was increased in rMKN45-PTX. The expression level of PKM1 was upregulated in rMKN45-PTX, leading to an increase in the PKM1/PKM2 ratio. Using xenograft models, we demonstrated that mouse subcutaneous tumors derived from rMKN45-PTX were significantly larger than those derived from MKN45 cells.
Conclusion
Under the stress of chemotherapeutic agent exposure, high expression of HIF-1α affects various downstream genes. Although the underlying mechanism is unknown, our data suggest that PKM1 is also a molecular target for gastric cancer treatment.
Keywords:
Introduction
Gastric cancer is the fourth most commonly diagnosed cancer and the second leading cause of cancer death worldwide.Citation1 Most patients in the advanced stages of disease are treated with chemotherapy to prolong their survival. However, even if the regimen is effective, the cancer becomes chemoresistant with repeated treatmentCitation2 and new strategies to counter chemoresistance in gastric cancer are therefore required.
Acquisition of chemoresistance is a complex and multifactorial phenomenon related to the tumor microenvironment, and the mechanism has not been fully elucidated. Cancer cell lines with resistance to anticancer drugs can provide valuable model systems to investigate the detailed molecular mechanisms. However, to date there have been few reports about the establishment of cancer cell lines resistant to anticancer drugs.Citation3–Citation6
Paclitaxel (PTX), a member of the taxane family, is a diterpenoid compound obtained from Taxus brevifolia.Citation7 It disturbs depolymerization of microtubules in malignant cellsCitation8 and thereby inhibits cell division and triggers apoptosis.Citation9 Because PTX is one of the most commonly used therapeutic agents in gastric cancer patients, it is necessary to develop new methods of overcoming resistance to PTX in clinical practice.
Previous experimental studies showed that hypoxia-inducible factor-1 alpha (HIF-1α) may be a factor involved in acquisition of chemoresistance.Citation10–Citation16 In various cancer, HIF-1α is the aryl hydrocarbon receptor nuclear translocator that forms a functional complex with β subunits under hypoxia, which transactivates more than 60 genes associated with angiogenesis, apoptosis and cell proliferation.Citation10–Citation12
Furthermore, nuclear factor kappa B (NF-κB) also may be linked to chemoresistance.Citation13 The NF-κB family is composed of the transcription factors that regulate the HIF system (NF-κB/HIF-1α pathway).Citation14 Liu et al demonstrated an association between HIF-1α expression and activation of the NF-κB signal pathway in chemoresistance acquisition using the vincristine-resistant gastric cancer cell line SGC7901/VCR.Citation15 It was recently noted that genes related to cancer metabolism such as pyruvate kinase muscle 1 (PKM1) may also play an important role in chemoresistance acquisition.Citation16
The purpose of this study was to elucidate the molecular mechanisms associated with HIF-1α and PKM1 that lead to acquired chemoresistance using a newly established PTX-resistant gastric cancer cell line.
Materials and methods
Antineoplastic agent
PTX was kindly provided by Bristol-Myers Squibb Co. (Tokyo, Japan). PTX was reconstituted in distilled water at appropriate concentrations and stored at −20°C until use. The reagent was used as recommended by this supplier.
Cell lines and cell culture
The gastric cancer cell line used in this study was MKN45, which was purchased from American Type Culture Collection (Rockville, MD, USA). Cells were maintained in RPMI-1640 medium supplemented with 10% fetal bovine serum, penicillin (100 U/mL) and streptomycin (100 μg/mL). The cells were maintained in a humidified atmosphere of 5% CO2 in air at 37°C.
Establishment of PTX-resistant cancer cell lines
PTX-resistant cells were established by exposure to increasing concentrations of PTX similar to the previously described method.Citation4–Citation6 MKN45 cells were initially cultured in RPMI containing PTX at a concentration of 0.1 nM and subsequently subcultured with increasing concentrations of PTX. Once surviving cells reached 80% confluence, they were passaged twice in the same concentration of PTX, after which the process was repeated with increasing doses of PTX until a cell population was selected that demonstrated more than three times of 50% growth inhibition (IC50) to PTX than the parental cell line. The resultant cell line that grew under the high concentrations of PTX was made as a drug-resistant gastric cancer cell line and named rMKN45-PTX. rMKN45-PTX used in this study was selected from a culture dish almost 1 year later. Experiments were performed after culturing for 2 weeks under the absence of PTX.
3-(4,5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay
The viability of cancer cells was determined by the MTT assay. Cells were seeded at 4 × 103 per well in 96-well plates and incubated overnight at 37°C in a humidified environment containing 5% CO2. PTX was dissolved in phosphate-buffered saline (PBS) and added to the cell culture medium at various concentrations (0–100 nM). After 48 h of treatment, the supernatant was discarded and MTT solution was added to each well (500 μg/mL final concentration) and incubated at 37°C for 3 h. Then, the supernatant was removed, and 150 μL of DMSO (Wako, Tokyo, Japan) was added. The absorbance of the solution was read at a wavelength of 535 nm using a microplate reader (Bio-Rad 550; Bio-Rad, Hercules, CA, USA). Cell viability was calculated as follows: viability = (absorbance of experimental wells)/(absorbance of control wells). All experiments were repeated at least three times. The IC50 of each chemotherapeutic drug was determined as the drug concentration showing 50% cell growth inhibition compared with the control cell growth. The resistance index (RI) was calculated as the IC50 of rMKN45-PTX/the IC50 of MKN45. Chemoresistance of cancer cell lines was defined by a RI of 3.0 or greater.
Flow cytometric analysis of cell cycle distribution
Single-cell suspension was collected, washed with cold PBS three times and fixed with 70% ethanol at 4°C overnight. After centrifugation, the supernatant was discarded and the cells were washed twice with cold PBS. The cells were then stained with propidium iodide. The sub-G0/G1, S and G2/M phase fractions of 2 × 104 cells were determined by flow cytometry using a FACS Caliber (Becton Dickinson).
Western blotting
Approximately 5 × 106 cells were lysed in RIPA buffer containing 1% protease inhibitor cocktail (Sigma-Aldrich Co.). The protein concentration of each sample was measured using a BCA protein assay kit (Pierce Biotechnology, Waltham, MA, USA). In total, 20 μg of protein from each sample was loaded onto 12.5% SDS-PAGE gels and subjected to electrophoresis. Proteins were transferred to polyvinylidene fluoride membranes (Bio-Rad Laboratories Inc., Hercules, CA, USA) and blocked with commercial gradient buffer (0.1% Tween-20; EZ Block; ATTO Corporation, Tokyo, Japan) at room temperature for 30 min. Blots were incubated overnight at 4°C with each primary antibody (see following text) and then incubated for 1 h with the appropriate horseradish peroxidase (HRP)-conjugated secondary antibodies. The antibody–antigen complexes were detected with an ECL Western blotting detection kit (GE Healthcare Japan Ltd., Japan) and a Light-Capture system (ATTO Corporation, NY, USA). To ensure equal protein loading, β-actin levels were measured using an anti-β-actin monoclonal antibody. We performed Western blot analysis by using same cell lysate. Primary antibodies against the following proteins were used: HIF-1α (rabbit polyclonal IgG, diluted 1:500; Santa Cruz Biotechnology, Inc., Dallas, TX, USA), Bax (rabbit monoclonal IgG, diluted 1:200; Abcam, Cambridge, UK), Bcl-xL (rabbit monoclonal IgG, diluted 1:1000; Abcam), caspase-3 (rabbit polyclonal IgG, diluted 1:1000; Abcam), multidrug resistance-associated protein (MRP; mouse polyclonal IgG, diluted 1:500; Abcam), P-glycoprotein (P-gp; rabbit polyclonal IgG, diluted 1:1000; Abcam), PKM1 (rabbit polyclonal IgG, diluted 1:500; Novus Biologicals), PKM2 (rabbit polyclonal IgG, diluted 1:1000; Abcam), vascular endothelial growth factor (VEGF; rabbit polyclonal IgG, diluted 1:1000; Santa Cruz Biotechnology, Inc.) and β-actin (mouse monoclonal IgG, diluted 1:10,000; Sigma-Aldrich Co.).
Enzyme-linked immunosorbent assay (ELISA)
Serum-free conditioned media (SF-CM) was prepared from rMKN45-PTX and MKN45 cells as previously reported.Citation17 Briefly, 1.0 × 106 cells were seeded into 100 mm tissue culture dishes with 10 mL RPMI supplemented with 10% FBS and incubated at 37°C for 3 days. To obtain SF-CM, the cells were washed twice with PBS and then incubated for 2 days with 5 mL of serum-free RPMI. SF-CM was harvested, centrifuged at 1500 × g for 5 min and passed through a filter (pore size: 0.45 μm). The concentration of VEGF was measured by ELISA (Quantikine; R&D Systems, Wiesbaden, Germany) according to the manufacturer’s instructions.
Mouse xenograft model
All animal experiments were performed according to standard guidelines of Kanazawa University. All procedures followed were in accordance with the ethical standards of the responsible committee on human experimentation (institutional and national) and with the Helsinki Declaration of 1964 and later versions. The study was approved by the Research Ethics Committee of Kanazawa University. Female immunocompromised BALB/c-nu/nu mice (Charles River Laboratories Inc., Yokohama-shi, Japan) aged 4–6 weeks were maintained in a sterile environment. For the subcutaneous model, a total of 5 × 106 MKN45 and rMKN45-PTX cells in 100 μL of RPMI were subcutaneously injected into the dorsal side of each mouse on day 0. The left dorsal side was injected with MKN45 and the right side was injected with rMKN45-PTX. Animals were carefully monitored and tumors of five sample were measured every 4 days. The tumor volume (V) was calculated according to the formula V = AB2/2, where A is the length of the major axis and B is the length of the minor axis. For the peritoneal dissemination model, 5 × 106 cells in 1,000 μL of RPMI were intraperitoneally injected into nude mice on day 0. After 14 days, mice were anesthetized with diethyl-ether and sacrificed, and the tumors and abdominal organs were excised together. Tumor specimens were then collected for immunohistochemical examination.
Immunohistochemistry
Tumor specimens were fixed in 10% neutral buffered formalin and embedded in paraffin. The expression levels of NF-κB p65 (mouse monoclonal IgG, diluted 1:100; Santa Cruz Biotechnology, Inc.), VEGF (rabbit polyclonal IgG, diluted 1:100; Santa Cruz Biotechnology, Inc.), Ki-67 (mouse monoclonal IgG, diluted 1:100; DakoCytomation, Glostrup, Denmark), PKM1 (rabbit polyclonal IgG, diluted 1:500; Novus Biologicals) and PKM2 (rabbit polyclonal IgG, diluted 1:1000; Abcam) were assessed immunohistochemically. Deparaffinized sections were pretreated by autoclaving in 10% citric acid buffer (pH 8.0) at 120°C for 15 min. Following treatment with protein block serum (DakoCytomation, Kyoto, Japan) for 10 min and incubation with 2% skim milk for 30 min to block non-specific reactions, sections were incubated with primary antibody at 4°C overnight. The Envision-polymer solution (horseradish peroxidase, DakoCytomation) was then applied for 1 h. Sections were examined using a fluorescence microscope (Olympus, Tokyo, Japan). The Ki-67 index was calculated as a percentage of expression within the whole section in all samples using a BZ-9000 BZII microscope (Keyence, Osaka, Japan).
Statistical analysis
Values are expressed as mean ± SD. Comparisons were made using one-way analysis of variance or Student’s t-test using SPSS statistical software, version 11.0 (SPSS). In all analyses, P < 0.05 indicated statistical significance.
Results
Establishment of PTX-resistant gastric cancer cell line
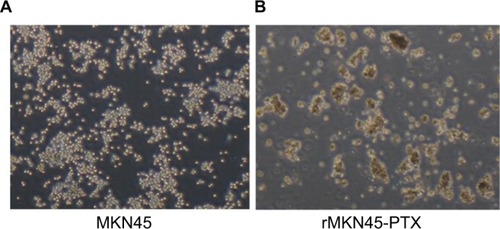
To obtain a PTX-resistant cell line, MKN45 cells were treated with increasing concentrations of PTX up to 20 nM. The IC50 value of PTX for MKN45 was 5.9 nM. By contrast, the IC50 level for rMKN45-PTX was 36 nM. RI calculated as the ratio of the IC50 level of rMKN45-PTX/IC50 of MKN45 was 6.1 (, ). Since the RI of rMKN45-PTX was greater than 3.0, this cell line was successfully established as a chemoresistant cancer cell line and was used for further investigation. Regarding morphologic appearance under a light microscope, rMKN45-PTX formed spheroid bodies, in contrast to MKN45 ().
Table 1 Establishment of drug-resistant cancer cell line
Figure 1 Drug sensitivity of rMKN45-PTX and MKN45 by MTT assay.
Note: Results are mean values from five experiments.
Abbreviations: MTT, 3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide; PTX, paclitaxel.
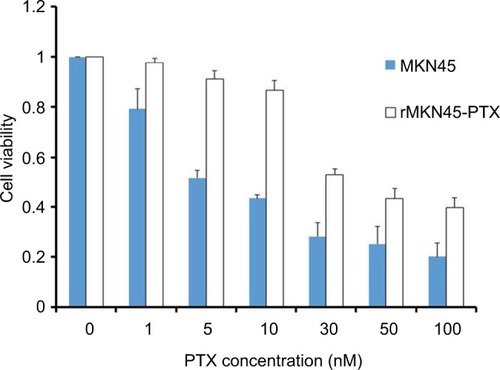
Figure 2 Morphologic differences between MKN45 (A) and rMKN45-PTX (B) under light microscope.
Note: rMKN45-PTX formed spheroid bodies (magnification ×40).
Abbreviation: PTX, paclitaxel.
Cell cycle distribution
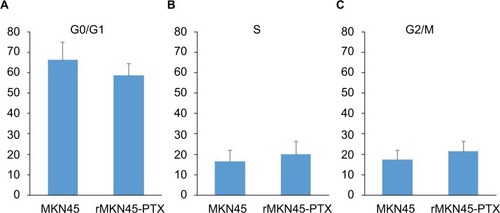
Our results showed no significant differences between rMKN45-PTX and the parent cell line in the proportion of cells in G0/G1 phase (66.2% vs. 58.6%, N.S.) and G2/M phase (17.3% vs. 21.4%, N.S.) (). Results are representative data of three different experiments.
Figure 3 Cell cycle distribution.
Notes: There were no significant differences in the percentage of cells in G0/G1 phase (A) and G2/M phase (C) between rMKN45-PTX and MKN45 compared with S phase (B). Results are the mean of three experiments.
Abbreviation: PTX, paclitaxel.
Expression of HIF-1α and chemoresistance-related proteins
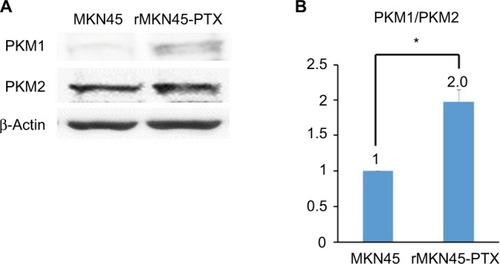
Western blotting results revealed that the expression level of HIF-1α in rMKN45-PTX was significantly higher than that in MKN45 as shown in . rMKN45-PTX cells showed upregulation of the antiapoptotic protein Bcl-xL and downregulation of the proapoptotic protein Bax and the effector enzyme caspase-3 in comparison with the parental cells (). Also, the expression of P-gp, MRP and VEGF was increased in rMKN45-PTX compared with the parent cells (). We also used ELISA to measure the level of VEGF in the culture media and showed that rMKN45-PTX had elevated levels of VEGF compared with MKN45 (). The expression level of PKM1 was upregulated in rMKN45-PTX compared with the parent cells. By contrast, the expression level of PKM2 was almost identical to that of the parent cells (). As a result, the PKM1/PKM2 ratio was significantly increased in rMKN45-PTX (). β-Actin was used as an internal control loading in each lane. Results are representative data of three different experiments.
Figure 4 Western blot analyses of HIF-1α. β-Actin was used as an internal control.
Note: Increased expression of HIF-1α was observed in rMKN45-PTX cells.
Abbreviations: PTX, paclitaxel; HIF-1α, hypoxia-inducible factor-1 alpha.
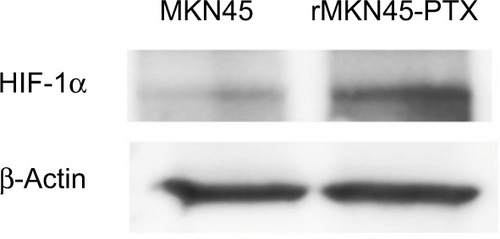
Figure 5 (A) Western blot analysis of Bax, Bcl-xL, caspase-3, P-gp, MRP, VEG and β-actin in MKN45 and rMKN45-PTX. Expression of Bax and caspase-3 was significantly downregulated, whereas expression of Bcl-xL, P-gp, MRP and VEGF was increased in rMKN45-PTX. The average signal intensity was standardized to that of β-Actin. (B) VEGF levels in gastric cancer cells were measured by an ELISA system.
Notes: Values represent mean ± SD (n = 3). *P < 0.05.
Abbreviations: P-gp, P-glycoprotein; MRP, multidrug resistance-associated protein; VEGF, vascular endothelial growth factor; PTX, paclitaxel.
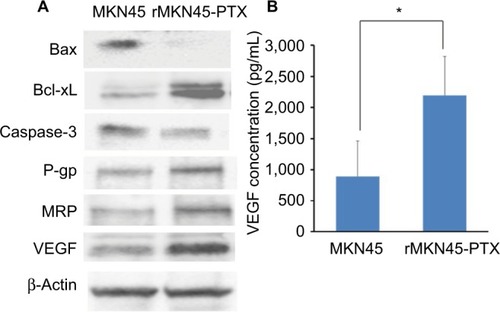
Figure 6 (A) Western blot analysis of PKM1 and PKM2. β-Actin was used as an internal control. (B) PKM1/PKM2 ratio calculated based on densitometric values of PKM1 and PKM2.
Notes: The PKM1/PKM2 ratio is presented relative to that of MKN45 cells (taken as 1.0). *P < 0.05.
Abbreviations: PKM, pyruvate kinase muscle; PTX, paclitaxel.
Tumor proliferation and progression
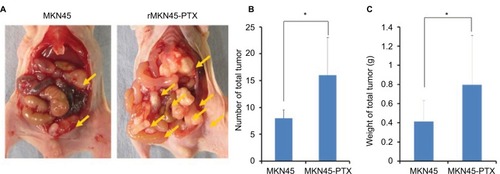
To examine proliferation and carcinogenesis of the drug-resistance cell line, we used a mouse xenograft model and evaluated the subcutaneous tumor volume. The time course of tumor growth is shown by the quantification of volume (). Tumors derived from rMKN45-PTX cells were significantly larger than those derived from MKN45 cells when measured at day 28 (P < 0.05). As shown in , the number and size of peritoneal dissemination nodules tended to be larger in rMKN45-PTX-injected mice than in MKN45-injected mice. Both tumor number and weight were significantly higher in rMKN45-PTX mice.
Figure 7 (A) Growth of tumors derived from rMKN45-PTX and MKN45. Tumor volume was measured every fourth day. (B) Representative images depicting the macroscopic appearance of the tumors at day 28.
Notes: Results are expressed as the mean ± SD (n = 5). *P < 0.05.
Abbreviation: PTX, paclitaxel.
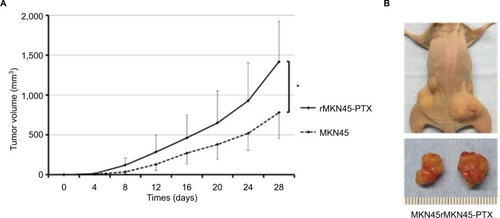
Figure 8 (A) Representative images depicting the macroscopic appearance of the tumors at day 14. Tumors were present in the peritoneal cavity (arrow). (B) Number and (C) weight (g) of disseminated nodules.
Notes: Results are mean ± SD of five experiments. *P < 0.05.
Abbreviation: PTX, paclitaxel.
Histological examination of xenograft tumors
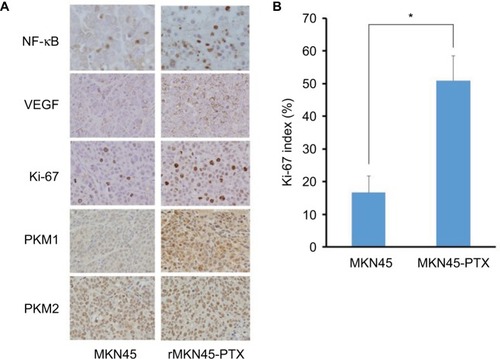
NF-κB immunoreactivity was increased in rMKN45-PTX tumors compared with MKN45 tumors (). VEGF expression in subcutaneous tumors derived from rMKN45-PTX cells was significantly higher than that in tumors from MKN45 cells (). Tumors from rMKN45-PTX exhibited increased expression of Ki-67 () and PKM1, although the expression of PKM2 was almost the same in both tumors (). Peritoneal dissemination tumors showed the same pattern of expression (data not shown).
Figure 9 (A) Immunohistochemical examination of NF-κB, VEGF, Ki-67, PKM1 and PKM2 in subcutaneous xenograft tumors (original magnification ×400). (B) The Ki-67 index is shown as a percentage.
Notes: Results are mean ± SD of five experiments. *P < 0.05.
Abbreviations: NF-κB, nuclear factor kappa B; VEGF, vascular endothelial growth factor; PKM, pyruvate kinase muscle; PTX, paclitaxel.
Discussion
Improving the efficacy of chemotherapy will require a better understanding of the mechanisms by which tumors become chemoresistant and development of strategies to overcome this resistance. The primary approach to this problem has been the establishment of chemoresistant cancer cell lines and identification of the underlying resistance mechanisms. In the present study, we established a PTX-resistant gastric cancer cell line (rMKN45-PTX) by stepwise exposure to PTX. The cell cycle distribution was similar in resistant and parental cell lines, but NF-κB and HIF-1α were over-expressed in rMKN45-PTX under normoxic conditions. Furthermore, we showed that the growth factor VEGF, the antiapoptosis marker Bcl-xL, multidrug transporters P-gp and MDR, and the glycolytic enzyme PKM1 were upregulated in rMKN45-PTX. Consequently, rMKN45-PTX-derived tumors showed significantly enhanced proliferation and progression in subcutaneous and peritoneal dissemination models.
Our study showed that the cell cycle distribution was not related to acquisition of PTX resistance. Wang et al reported that cell cycle distribution may be an integral part of mechanisms responsible for chemoresistance.Citation4 Furthermore, previous studies demonstrated that alteration of cell cycle distribution might be associated with chemoresistance to specific drugs including oxaliplatin and irinotecan, although no studies have demonstrated an association between taxan and cell cycle distribution.Citation18 These findings suggest that there may be a different mechanism underlying acquisition of PTX resistance.
In the present study, we showed that NF-κB and HIF-1α expression was increased during exposure of the parent cell line to PTX. Previous studies suggested that HIF-1α might be a contributor to chemoresistance acquired by hypoxia and in part explain the increased level of intracellular HIF-1α associated with resistance to therapy in head and neck, ovarian, esophageal, prostate and gastric cancer.Citation15,Citation19–Citation22 However, HIF-1α expression is induced not only in hypoxic conditions but also in non-hypoxic conditions and in response to other stressors such as chemotherapy and radiation.Citation3,Citation23 Also, Montagut et al demonstrated that NF-κB activation was significantly correlated with chemoresistance and the number of breast cancer patients with NF-κB activation increased after chemotherapy exposure.Citation13
HIF-1α is known to transcriptionally upregulate VEGF expression.Citation10,Citation24 In this study, we showed that VEGF was overexpressed in rMKN45-PTX cells by Western blotting, ELISA and immunohistochemistry. VEGF is a multifunctional cellular factor that can induce neovascularization and increase capillary permeability.Citation25,Citation26 As a result, the tumor vasculature shows an abnormal structure that is inefficient for oxygen and drug delivery.Citation27 Moreover, VEGF is one of the major factors that contributes to metastasis in various cancers, and VEGF overexpression has been associated with tumor progression and poor prognosis.Citation10,Citation28–Citation30 We demonstrated that rMKN45-PTX-derived tumors showed significantly enhanced tumor proliferation and progression in a subcutaneous model and a peritoneal dissemination model. In our clinical experience, patients with failure of cancer chemotherapy often rapidly develop tumor progression.
Apoptosis is an important mechanism for cell death following treatment with various types of chemotherapy. Caspase-3 is a proapoptotic effector enzyme that interacts with caspase-8 and caspase-9. Bax is a proapoptotic member of the Bcl-2 family that plays a key role in the induction of mitochondria-dependent apoptosis. By contrast, Bcl-xL is an antiapoptotic family member that can neutralize Bax function in the initiation of cell death. Overexpression of Bcl-xL protein occurs in many types of cancer cell and prevents cell death induced by anticancer drugs and radiation.Citation31 Our findings demonstrated that the expression of Bcl-xL was markedly increased in rMKN45-PTX, which is in line with previous studies.Citation32–Citation34 The involvement of HIF-1α in chemoresistance acquisition is possibly due to its role in the regulation of some apoptosis-related genes.
Accumulating evidence has shown that the mechanisms responsible for chemoresistance are associated with overexpression of two ATP-dependent drug transporter proteins, P-gp and MRP.Citation35–Citation37 These proteins act as ATP-dependent outward transport pumps and decrease the intracellular accumulation of drugs by reducing the cotransport mechanism of glutathione.Citation38 In particular, Sparreboom et al showed that P-gp at the luminal side of the intestinal epithelium appears to be an important component of the defense against PTX, and P-gp was therefore associated with chemoresistance acquisition.Citation39 Furthermore, a previous study showed that HIF-1α expression is correlated with MDR1/P-gp expression in colon carcinoma tissue and a colon cancer cell line.Citation40 We found that both P-gp and MRP were expressed in rMKN45-PTX, indicating that both P-gp and MRP were induced by HIF-1α and play an important role in PTX transportation.
In the present study, we examined the association of pyruvate kinase muscle (PKM) and chemoresistance acquisition. PKM has two splicing variants: PKM1 is expressed in most normal tissues, whereas PKM2 is expressed in the embryonic stage cells and cancer cells.Citation41 Recently, it was noted that cancer metabolism-related genes play an important role in chemoresistance acquisition. Especially, the Warburg effect has been reported in cancer cells.Citation42 This phenomenon is that cancer cells use glycolysis for energy for energy production even when an adequate amount of oxygen is present.Citation42 A previous study showed that PKM2 promotes the Warburg effect; the coordinated control of metabolism and proliferation by PKM2 is essential for tumorigenesis.Citation43,Citation44 As a result, it was previously thought that inhibitors and activators of PKM2 are well underway to evaluate their anticancer effects and suitability.Citation45 On the other hand, PKM1 promotes oxidase phosphorylation (OXPHOS).Citation43,Citation46 It was also reported that metabolic switching from glycolysis to OXPHOS contributes to chemoresistance acquisition.Citation47 Taniguchi et al reported that PKM2 was expressed in both parent and chemoresistant cells, whereas PKM1 was commonly upregulated in various chemoresistant cells, and knockdown of PKM1 sensitized chemoresistant cells to chemotherapy.Citation16 They concluded that PKM1 plays important roles in the chemoresistance acquisition of cancer cells.Citation16 In the present study, we similarly demonstrated that PKM1 was upregulated in rMKN45-PTX cells in vivo and in vitro, although PKM2 was expressed in both cells. Their results suggested that PKM1 expression at the same time as PKM2 might be one of the key regulators of chemoresistance acquisition in gastric cancer.
There were several limitations to our study. First, we only evaluated one cell line and one chemotherapeutic agent. It is therefore necessary to evaluate other gastric cancer cell lines, other cancer types and other anticancer drugs. Second, we have no data on HIF-1α knockdown in parental and chemoresistant cells using si-RNA specific for HIF-1α. Third, we did not study the associated with HIF-1α and PKM1 expression under conditions of chemotherapy. Nevertheless, this study demonstrated HIF-1α expression in our chemoresistant cancer cell line and showed a relationship between PKM1 expression and chemoresistance.
Conclusion
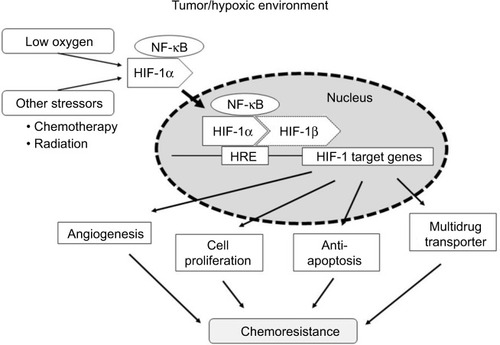
rMKN45-PTX cells showed overexpression of HIF-1α and enhanced tumor proliferation and progression. HIF-1α is a key molecule in chemoresistance acquisition, most likely through its impact on angiogenesis, cell proliferation, apoptosis and multidrug transporters (). Furthermore, PKM1 might also be important in chemoresistance acquisition. Further research into the relationship between chemotherapy and chemoresistance may provide additional novel treatment strategies. In addition to inhibition of HIF-1α, our data suggest that PKM1 might also be a target for tumor-specific molecular-based therapy.
Figure 10 Scheme illustrating a chemoresistance mechanism involving HIF-1α.
Abbreviations: HIF-1α, hypoxia-inducible factor-1 alpha; HRE, hypoxic response element; NF-κB, nuclear factor kappa B.
Acknowledgments
We are grateful to members of the Department of Gastroenterological Surgery of Kanazawa University for their helpful suggestions. We thank Mary Derry, PhD ELS, from Edanz Group (www.edanzediting.com/ac) for editing a draft of this manuscript.
Disclosure
The authors report no conflicts of interest in this work.
References
- OrdituraMGaliziaGSforzaVTreatment of gastric cancerWorld J Gastroenterol20142071635164924587643
- RothADChemotherapy in gastric cancer: a never ending sagaAnn Oncol200314217517712562640
- LiuLNingXSunLInvolvement of MGr1-Ag/37LRP in the vincristine-induced HIF-1 expression in gastric cancer cellsMol Cell Biochem20073031–215116017476462
- ZhangXYashiroMQiuHNishiiTMatsuzakiTHirakawaKEstablishment and characterization of multidrug-resistant gastric cancer cell linesAnticancer Res201030391592120393015
- ChungYMParkSParkJKKimYKangYYooYDEstablishment and characterization of 5-fluorouracil-resistant gastric cancer cellsCancer Lett200015919510110974411
- QiuHYashiroMZhangXMiwaAHirakawaKA FGFR2 inhibitor, Ki23057, enhances the chemosensitivity of drug-resistant gastric cancer cellsCancer Lett20113071475221482024
- DonaldsonKLGoolsbyGLKienerPAWahlAFActivation of p34cdc2 coincident with taxol-induced apoptosisCell Growth Differ1994510104110507848905
- GabizonAPriceDCHubertyJBresalierRSPapahadjopoulosDEffect of liposome composition and other factors on the targeting of liposomes to experimental tumors: biodistribution and imaging studiesCancer Res19905019637163781698120
- WardMWHuberHJWeisováPDüssmannHNichollsDGPrehnJHMitochondrial and plasma membrane potential of cultured cerebellar neurons during glutamate-induced necrosis, apoptosis, and toleranceJ Neurosci200727318238824917670970
- StoeltzingOMcCartyMFWeyJSRole of hypoxia-inducible factor 1α in gastric cancer cell growth, angiogenesis, and vessel maturationJ Natl Cancer Inst2004961294695615199114
- McMahonSGrondinFMcDonaldPPRichardDEDuboisCMHypoxia-enhanced expression of the proprotein convertase furin is mediated by hypoxia-inducible factor-1: impact on the bioactivation of proproteinsJ Biol Chem200528086561656915611046
- Muñoz-NájarUMNeurathKMVumbacaFClaffeyKPHypoxia stimulates breast carcinoma cell invasion through MT1-MMP and MMP-2 activationOncogene200625162379239216369494
- MontagutCTusquetsIFerrerBActivation of nuclear factor-kappa B is linked to resistance to neoadjuvant chemotherapy in breast cancer patientsEndocr Relat Cancer200613260761616728586
- RiusJGumaMSchachtrupCNF-kappaB links innate immunity to the hypoxic response through transcriptional regulation of HIF-1 alphaNature2008453719680781118432192
- LiuLNingXSunLHypoxia-inducible factor-1 alpha contributes to hypoxia-induced chemoresistance in gastric cancerCancer Sci200899112112817953712
- TaniguchiKSakaiMSugitoNPKM1 is involved in resistance to anti-cancer drugsBiochem Biophys Res Commun2016473117418027012213
- OkazakiMFushidaSHaradaSThe angiotensin II type 1 receptor blocker candesartan suppresses proliferation and fibrosis in gastric cancerCancer Lett20143551465325224569
- DonaldsonKLGoolsbyGLWahlAFCytotoxicity of the anticancer agents cisplatin and taxol during cell proliferation and the cell cycleInt J Cancer19945768478557911457
- QuinteroMMackenzieNBrennanPAHypoxia-inducible factor 1 (HIF-1) in cancerEur J Surg Oncol200430546546815135470
- RussellDLIanSAdrianLThe role of hypoxia-inducible factor-1 in three-dimensional tumor growth, apoptosis, and regulation by the insulin-signaling pathwayCancer Res200565104147415215899805
- UmJHKangCDBaeJHAssociation of DNA-dependent protein kinase with hypoxia inducible factor-1 and its implication in resistance to anticancer drugs in hypoxic tumor cellsExp Mol Med200436323324215272235
- SemenzaGLSignal transduction to hypoxia-inducible factor 1Biochem Pharmacol2002645–699399812213597
- HaradaHInoueMItasakaSCancer cells that survive radiation therapy acquire HIF-1 activity and translocate towards tumour blood vesselsNat Commun2012378322510688
- KitajimaYMiyazakiKThe critical impact of HIF-1a on gastric cancer biologyCancers (Basel)201351152624216696
- ConnollyDTHeuvelmanDMNelsonRTumor vascular permeability factor stimulates endothelial cell growth and angiogenesisJ Clin Invest1989845147014782478587
- LeungDWCachianesGKuangWJGoeddelDVFerraraNVascular endothelial growth factor is a secreted angiogenic mitogenScience19892464935130613092479986
- TrédanOLacroix-TrikiMGuiuSAngiogenesis and tumor microenvironment: bevacizumab in the breast cancer modelTarget Oncol201510218919825185646
- ChenWTHuangCJWuMTYangSFSuYCChaiCYHypoxia-inducible factor 1 alpha is associated with risk of aggressive behavior and tumor angiogenesis in gastrointestinal stromal tumorJpn J Clin Oncol200535420721315845570
- MizokamiKKakejiYOdaSClinicopathological significance of hypoxia-inducible factor 1 alpha overexpression in gastric carcinomasJ Surg Oncol200694214915416847924
- FukudaRKellyBSemenzaGLVascular endothelial growth factor gene expression in colon cancer cells exposed to prostaglandin E2 is mediated by hypoxia-inducible factor 1Cancer Res20036392330233412727858
- BargouRCWagenerCBommertKOverexpression of the death-promoting gene Bax-alpha which is downregulated in breast cancer restores sensitivity to different apoptotic stimuli and reduces tumor growth in SCID miceJ Clin Invest19969711265126598647960
- CrawfordANahtaRTargeting Bcl-2 in herceptin-resistant breast cancer cell linesCurr Pharmacogenomics Person Med20119318419022162984
- ShajahanANDobbinZCHickmanFEDakshanamurthySClarkeRTyrosine phosphorylated caveolin-1 (Tyr-14) increases sensitivity to paclitaxel by inhibiting BCL2 and BCLxL proteins via c-Jun N-terminal kinase (JNK)J Biol Chem201228721176821769222433870
- ZhaoQTanBBLiYFanLQYangPGTianYEnhancement of drug sensitivity by knockdown of HIF-1α in gastric carcinoma cellsOncol Res201623312913626931435
- EndoKMaeharaYKusumotoTIchiyoshiYKuwanoMSugimachiKExpression of multidrug-resistance-associated protein (MRP) and chemo-sensitivity in human gastric cancerInt J Cancer19966833723778903480
- FanKFanDChengLFLiCExpression of multidrug resistance-related markers in gastric cancerAnticancer Res2000206C4809481411205224
- ZhaoYXiaoBChenBQiaoTFanDUpregulation of drug sensitivity of multidrug-resistant SGC7901/VCR human gastric cancer cells by bax gene transductionChin Med J (Engl)20001131197798011776130
- XuHWXuLHaoJHQinCYLiuHExpression of P-glycoprotein and multidrug resistance-associated protein is associated with multidrug resistance in gastric cancerJ Int Med Res2010381344220233511
- SparreboomAvan AsperenJMayerULimited oral bioavailability and active epithelial excretion of paclitaxel (Taxol) caused by P-glycoprotein in the intestineProc Natl Acad Sci U S A1997945203120359050899
- ChenJDingZPengYHIF-1α inhibition reverses multidrug resistance in colon cancer cells via downregulation of MDR1/P-glycoproteinPLoS One201496e9888224901645
- ChristofkHRVander HeidenMGWuNAsaraJMCantleyLCPyruvate kinase M2 is a phosphotyrosine-binding proteinNature2008452718418118618337815
- Vander HeidenMGCantleyLCThompsonCBUnderstanding the Warburg effect: the metabolic requirements of cell proliferationScience200932459301029103319460998
- ChenMZhangJManleyJLTurning on a fuel switch of cancer: hnRNP proteins regulate alternative splicing of pyruvate kinase mRNACancer Res201070228977898020978194
- YangWXiaYJiHNuclear PKM2 regulates β-catenin transactivation upon EGFR activationNature2011480737511812222056988
- TamadaMSuematsuMSayaHPyruvate kinase M2: multiple faces for conferring benefits on cancer cellsClin Cancer Res201218205554556123071357
- ChristofkHRVander HeidenMGHarrisMHThe M2 splice isoform of pyruvate kinase is important for cancer metabolism and tumour growthNature2008452718423023318337823
- WolfDAIs reliance on mitochondrial respiration a “chink in the armor” of therapy-resistant cancer?Cancer Cell201426678879525490445