Abstract
There is evidence that proteases and antiproteases participate in the iron homeostasis of cells and living systems. We tested the postulate that α-1 antitrypsin (A1AT) polymorphism and the consequent deficiency of this antiprotease in humans are associated with a systemic disruption in iron homeostasis. Archived plasma samples from Alpha-1 Foundation (30 MM, 30 MZ, and 30 ZZ individuals) were analyzed for A1AT, ferritin, transferrin, and C-reactive protein (CRP). Plasma samples were also assayed for metals using inductively coupled plasma atomic emission spectroscopy (ICPAES). Plasma levels of A1AT in MZ and ZZ individuals were approximately 60% and 20% of those for MM individuals respectively. Plasma ferritin concentrations in those with the ZZ genotype were greater relative to those individuals with either MM or MZ genotype. Plasma transferrin for MM, MZ, and ZZ genotypes showed no significant differences. Linear regression analysis revealed a significant (negative) relationship between plasma concentrations of A1AT and ferritin while that between A1AT and transferrin levels was not significant. Plasma CRP concentrations were not significantly different between MM, MZ, and ZZ individuals. ICPAES measurement of metals confirmed elevated plasma concentrations of nonheme iron among ZZ individuals. Nonheme iron concentrations correlated (negatively) with levels of A1AT. A1AT deficiency is associated with evidence of a disruption in iron homeostasis with plasma ferritin and nonheme iron concentrations being elevated among those with the ZZ genotype.
Introduction
Alpha-1 antitrypsin (A1AT; also known as α-1 proteinase inhibitor) is a serum glycoprotein which predominantly (80%) originates from hepatocytes.Citation1 A1AT inhibits a wide variety of proteases (eg, neutrophil elastase and proteinase-3) to contribute to a proteinase/antiproteinase balance.Citation2 It is postulated that this acute phase protein participates in limiting host tissue injury by proteases at sites of inflammation;Citation3 a proposed critical function is the prevention of lung injury associated with disruption of connective tissue by neutrophil elastase enzyme.Citation1
A1AT deficiency is an autosomal recessive genetic disorder caused by its defective production leading to both decreased activity in body tissues and deposition of excessive abnormal protein in the liver.Citation1,Citation2 While M is the normal allele for A1AT, there are over 80 variant mutations of the gene. The Z allele results in a glutamate to lysine mutation at position 342 while the S allele produces a glutamate to valine mutation at position 264. In individuals with SS, MZ and SZ genotypes, blood levels of A1AT are reduced to between 40% and 60% of normal levels (ie, those observed among MM individuals). Among those who do not smoke, this A1AT concentration is almost always sufficient to protect the lungs from the effects of elastase. However, among individuals with the ZZ genotype, A1AT levels are usually less than 15% of normal and these deficient patients can develop lung disease at a young age. In addition, those with the ZZ genotype can develop liver disease; this is associated with impaired A1AT secretion and its consequent accumulation in this specific tissue. Therefore, A1AT deficient patients can present with lung disease (eg, panacinar emphysema [chronic obstructive pulmonary disease], pneumothorax, asthma, and bronchiectasis) and liver disease (eg, hepatitis, cirrhosis, and hepatocellular carcinoma).Citation4 People of northern and western European ancestry are at the greatest risk for A1AT deficiency and its associated diseases;Citation5 4% carry the PiZ allele and between 1 in 1,500 and 1 in 2,500–3,000 are ZZ.
In addition to siderophores and ferrireductases, microbials employ proteases to mobilize host sources of iron to support their growth and replication.Citation6 These proteases cleave iron-transport and -storage proteins allowing utilization of the metal by the microbe.Citation7,Citation8 Corroborating a potential interaction between metal homeostasis and proteases, elevated iron concentrations impact expression of proteases and their activities.Citation9–Citation12 Animal and human investigation similarly suggests a participation of proteases in iron homeostasis.Citation13–Citation15 The intratracheal instillation of a single dose of neutrophil elastase in an animal model increased lung iron concentrations. Iron homeostasis is disrupted among cystic fibrosis patients in whom airway elastase content and activity is excessive; cystic fibrosis patients have elevated iron and ferritin concentrations in both the sputum and bronchoalveolar lavage.
A participation of proteases also suggests a potential involvement of anti-proteases in iron homeostasis. A1AT is the antiprotease in greatest concentration in humans. A participation of A1AT in the iron homeostasis of humans has been previously suggested with observations of deficient patients developing cirrhosis characterized by significant iron accumulation.Citation16 An involvement of neutrophil elastase and A1AT in disrupting iron homeostasis of the lung was supported by investigation which demonstrated a proteolytic cleavage of both iron-transport and -storage proteins with subsequent increase in nonheme tissue iron concentrations.Citation17 Finally, patients treated with antiproteases, and thus altering protease activity, can show evidence of disruptions in iron homeostasis.Citation18 We further explored the relationship between A1AT and iron homeostasis in humans by testing the postulate that its deficiency is associated with a systemic disruption in iron homeostasis.
Methods
Samples from MM, MZ, and ZZ individuals
Archived plasma samples were kindly provided by the Alpha-1 Foundation (Miami, FL). These included 90 nonfasting plasma specimens from 30 MM, 30 MZ, and 30 ZZ individuals who had been screened as family members of a patient with A1AT deficiency. The health status of the donor was not known. Institutional Review Board exemption was granted from the University of North Carolina for analyses of these blood samples.
Plasma assays
Samples were analyzed for A1AT, ferritin, iron, transferrin, and C-reactive protein (CRP). A1AT and CRP concentrations were measured using immunoturbidimetric assay kits (DiaSorin Inc, Stillwater, MN). Ferritin was quantified by also using an immunoturbidimetric assay (Kamiya Biomedical Company, Seattle, WA). Transferrin concentrations were analyzed using an immunoprecipitin analysis (INCSTAR Corporation, Stillwater, MN). Assays were modified for use on the Konelab Arena 30 (ThermoLab Systems, Espoo, Finland).
Measurements of metals in plasma
Plasma samples were hydrolyzed at 70°C in 3 N HCl/10% trichloroacetic acid for 24 hours. After centrifugation, metals in the supernatant were measured using inductively coupled plasma optical emission spectroscopy (ICPOES; Model Optima 4300D; Perkin Elmer, Norwalk, CT).
Statistics
Data are expressed as mean values ± standard error unless otherwise specified. Differences between groups were compared using one-way analysis of variance. The post-hoc test employed was Duncan’s Multiple Range test. General linear regression modeling was used in delineating a relationship between two continuous variables. Significance was assumed at P < 0.05.
Results
Demographics of healthy individuals providing archived plasma samples are listed (). There were differences in the gender distributions between the MM, MZ, and ZZ cohorts and subsequently results were provided for males and females. Disparities in age and ethnicity between the cohorts were not significant.
Table 1 Demographics of healthy individuals providing plasma samples
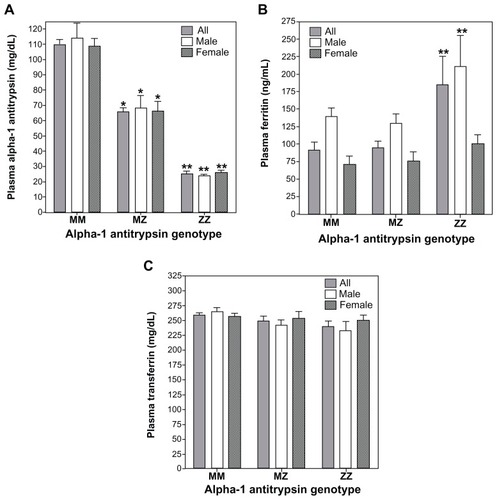
Among the samples collected by the Alpha-1 Foundation, plasma levels of A1AT for MZ and ZZ individuals were approximated as 60% and 20% of those for MM individuals respectively (). This concentration of A1AT approximates those serum values reported in other study populations in which ZZ individual demonstrate levels 15% of MM subjects.Citation19 There were significant differences in plasma ferritin concentrations between MM, MZ, and ZZ cohorts (). Those with the ZZ genotype showed greater ferritin values relative to both MM and MZ genotypes while there were no differences between MM and MZ (; F = 5.54; P = 0.02). When separate analyses of variance were conducted for males and females, the ferritin concentration in the ZZ cohort was greater than both MM and MZ for males while only greater than MM subjects for females. Plasma transferrin for MM, MZ, and ZZ cohorts showed no significant differences between them regardless of whether all were included in the analysis or sex-specific values were considered (; F = 2.97; P = 0.09).
Figure 1 Concentrations of A1AT, ferritin, and transferrin among MM, MZ, and ZZ cohorts. Relative to the MM genotype, significant decrements in plasma A1AT concentrations were observed in both MZ and ZZ subjects (A). Relative to MM subjects, plasma ferritin levels were significantly increased only in the ZZ cohort (B). Transferrin did not demonstrate differences between the three genotypes (C).
Abbreviation: A1AT, α-1 antitrypsin.
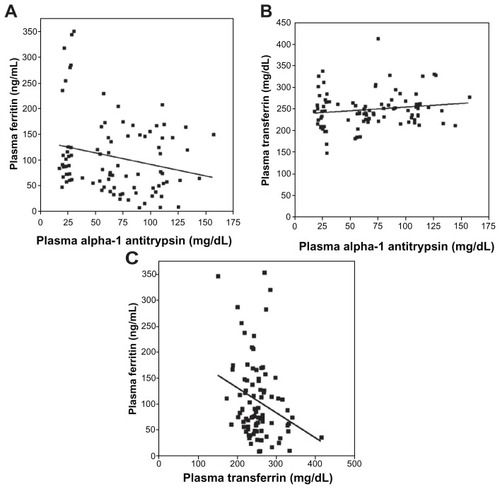
Linear regression analysis revealed a significant (negative) relationship between plasma concentrations of A1AT and ferritin (; r = 0.26; P = 0.01). Linear regression between A1AT and transferrin levels was not significant (; r = 0.18; P = 0.09). The linear regression between ferritin and transferrin was significant (negative) (; r = 0.39; P < 0.01).
Figure 2 Linear correlations between plasma concentrations of A1AT, ferritin, and transferrin. The correlation between A1AT and ferritin concentrations (A) reached significance while that between A1AT and transferrin (B) did not. The relationship between plasma ferritin and transferrin was negative and significant (C).
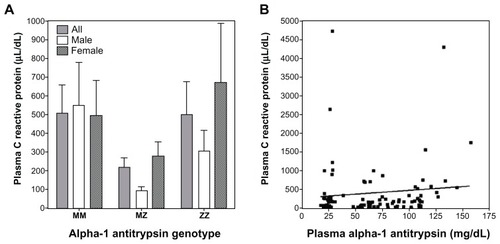
Plasma CRP concentrations were not significantly different between MM, MZ, and ZZ individuals (). Linear regression analysis did not show a significant association between CRP and A1AT (; r = 0.10; P = 0.36). Regarding the relationships between CRP and indices of iron homeostasis, there was no significant association with either ferritin (r = 0.17; P = 0.11) or transferrin (r = 0.20; P = 0.06).
Figure 3 C-reactive protein (CRP) levels in MM, MZ, and ZZ individuals and its correlation with A1AT concentration. There were no significant differences in CRP among the MM, MZ, and ZZ genotypes (A). Similarly, there was no significant correlation between plasma A1AT and CRP concentrations (B).
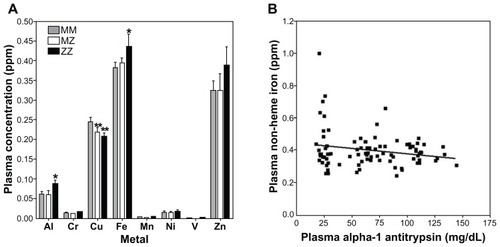
Plasma metal concentrations were significantly different between MM and ZZ cohorts for aluminum (F = 3.10; P = 0.05) and iron (F = 3.37; P = 0.04) (). Zinc also appeared to increase in the ZZ cohort, but this did not reach significance (F = 0.49; P = 0.61). Aluminum and iron were elevated in those individuals with the ZZ genotype relative to the MM genotype; disparities between MM and MZ were not significant. In contrast, copper levels among those with both MZ and ZZ genotypes were significantly lower relative to the MM cohort (F = 4.33; P = 0.02) (). Correlations of plasma metal concentration with levels of A1AT were significant only for iron (; r = 0.21; P = 0.05).
Figure 4 Plasma metal concentrations among MM, MZ, and ZZ cohorts. Aluminum and iron concentrations were observed to increase in ZZ individuals relative to those with MM and MZ genotype (A). In contrast, copper levels decreased in both MZ and ZZ cohorts (A). Only the correlation between A1AT and iron concentration was significant (B).
Abbreviation: A1AT, α-1 antitrypsin.
Discussion
Ferritin is considered the best available index to measure total body stores of iron.Citation20–Citation22 Significant differences in plasma ferritin concentrations between MM and ZZ individuals support an impact of A1AT deficiency on iron homeostasis in humans. Similarly, plasma iron levels were increased in the ZZ genotype. While the MZ genotype decreased A1AT levels, it did not influence ferritin and iron concentrations; this may reflect either a true lack of effect of A1AT genotype on iron homeostasis or the sample number may have been insufficient to demonstrate a difference. The significant correlations between both plasma levels of ferritin and iron with A1AT also suggest a participation of this anti-protease in iron homeostasis. A relationship between A1AT and ferritin can possibly be predicted since both would increase with inflammation. However, CRP levels were not significantly different between genotypes (MM, MZ, and ZZ) and there was no significant relationship between CRP and ferritin suggesting the increase in ferritin levels among ZZ individuals was not the result of disparities in inflammatory status. In addition, while both A1AT and ferritin are positive acute phase reactants, the relationship between them in our study cohort was an inverse one; that is, as A1AT decreased, ferritin increased. It is not possible to argue that inflammation is responsible for an increase in one index while the other is decreased.
This investigation does not address the molecular basis for a disruption in iron homeostasis in patients with A1AT deficiency. Proteases are employed as a third pathway of iron acquisition by microbials (in addition to siderophores and ferrireductases).Citation23,Citation24 These proteins have a capacity to cleave iron-transport and -storage proteins thus allowing utilization of the metal by the microbe.Citation25–Citation27 Reflecting potential interactions between metal availability and proteases, increased iron concentrations can impact expression and/or activity of collagenase, elastase, alkaline proteinase, and metalloproteases.Citation9,Citation10,Citation12,Citation28,Citation29 In addition, in vitro exposure to the protease inhibitor A1AT alters iron homeostasis in eukaryotic cells.Citation30–Citation34 This serpin, which primarily inhibits neutrophil elastase, can elevate intracellular ferritin concentrations in cultured cells. In vivo exposure to protease disrupted iron homeostasis in an animal model.Citation17 Finally, among patients with A1AT deficiency, iron homeostasis is disrupted.Citation16,Citation35,Citation36 Accordingly, one mechanism through which A1AT deficiency can affect a disruption in iron homeostasis is by allowing an unrestricted activity of specific proteases which would result in increased iron availability. This will lead to elevated tissue concentrations of iron especially in the liver and spleen, which are major cites of storage. Reflecting this, blood ferritin will be increased among individuals with A1AT deficiency.
This disruption in iron homeostasis associated with A1AT deficiency can assist in understanding the clinical presentation of such patients. Clinical manifestations of A1AT deficiency most frequently include liver and lung disease. Liver tissue is one of the major sites for iron storage. With a lack of A1AT, there will be an accumulation of iron with subsequent elevations in liver concentrations. Elevated liver iron is associated with hepatitis, cirrhosis, and hepatocarcinoma, which are the manifestations of disease in A1AT deficiency among ZZ individuals. Inhaled exposures (eg, particles) precipitate a biological effect in the lung by disrupting the normal iron homeostasis and increasing the availability of host iron.Citation37 Subsequently, particle exposure in A1AT deficient individuals is predicted to be associated with a higher incidence of disease (eg, chronic obstructive pulmonary disease among smokers and those exposed to elevated particle levels).Citation38–Citation40
We conclude that A1AT deficiency is associated with evidence of a disruption in iron homeostasis in ZZ individuals with plasma ferritin and iron concentrations being elevated. Such a disruption in iron homeostasis can contribute to understanding the clinical presentation of A1AT deficiency.
Acknowledgments
We thank the Alpha-1 Foundation for allowing access to archived plasma samples.
Disclosure
Bernard Fischer was supported in part by an Alpha-1 Foundation research grant. The other authors have no conflict of interests to declare.
References
- BlancoILaraBde SerresFEfficacy of alpha1-antitrypsin augmentation therapy in conditions other than pulmonary emphysemaOrphanet J Rare Dis201161421486454
- LomasDAThe selective advantage of alpha1-antitrypsin deficiencyAm J Respir Crit Care Med2006173101072107716439713
- TuderRMJanciauskieneSMPetracheILung disease associated with alpha1-antitrypsin deficiencyProc Am Thorac Soc20107638138621030517
- StollerJKAboussouanLSAlpha1-antitrypsin deficiencyLancet200536594782225223615978931
- LuisettiMSeersholmNAlpha1-antitrypsin deficiency. 1: epidemiology of alpha1-antitrypsin deficiencyThorax200459216416914760160
- MarxJJIron and infection: competition between host and microbes for a precious elementBest Pract Res Clin Haematol200215241142612401315
- BritiganBEHayekMBDoebbelingBNFickRBJrTransferrin and lactoferrin undergo proteolytic cleavage in the Pseudomonas aeruginosa-infected lungs of patients with cystic fibrosisInfect Immun19936112504950558225581
- MillerRABritiganBEProtease-cleaved iron-transferrin augments oxidant-mediated endothelial cell injury via hydroxyl radical formationJ Clin Invest1995956249125007769095
- GardiCArezziniBFortinoVComportiMEffect of free iron on collagen synthesis, cell proliferation and MMP-2 expression in rat hepatic stellate cellsBiochem Pharmacol20026471139114512234617
- ShigematsuTFukushimaJOyamaMTsudaMKawamotoSOkudaKIron-mediated regulation of alkaline proteinase production in Pseudomonas aeruginosaMicrobiol Immunol200145857959011592632
- BrumlikMJStoreyDGZinc and iron regulate translation of the gene encoding Pseudomonas aeruginosa elastaseMol Microbiol1992633373441552848
- OkazakiIBrinckerhoffCESinclairJFSinclairPRBonkowskyHLHarrisEDJrIron increases collagenase production by rabbit synovial fibroblastsJ Lab Clin Med19819733964026257809
- StitesSWPlautzMWBaileyKO’Brien-LadnerARWesseliusLJIncreased concentrations of iron and isoferritins in the lower respiratory tract of patients with stable cystic fibrosisAm J Respir Crit Care Med1999160379680110471599
- ReidDWLamQTSchneiderHWaltersEHAirway iron and iron-regulatory cytokines in cystic fibrosisEur Respir J200424228629115332399
- LuceyECStonePJChristensenTGBreuerRSniderGLAn 18-month study of the effects on hamster lungs of intratracheally administered human neutrophil elastaseExp Lung Res19881456716863224598
- RabinovitzMGavalerJSKellyRHVan ThielDHAssociation between heterozygous alpha 1-antitrypsin deficiency and genetic hemochromatosisHepatology19921611451481618466
- FischerBMDomowiczDAZhengSNeutrophil elastase increases airway epithelial nonheme iron levelsClin Transl Sci20092533333920411049
- McNabbJCCappaJARossJWDisorders of iron metabolism associated with protease inhibitor therapyClin Infect Dis200133341341411438916
- Alpha 1-antitrypsin deficiency: memorandum from a WHO meetingBull World Health Organ19977553974159447774
- JacobsEMHendriksJCvan DeursenCTSeverity of iron overload of proband determines serum ferritin levels in families with HFE-related hemochromatosis: the HEmochromatosis FAmily StudyJ Hepatol200950117418319008010
- HerbertVJayatillekeEShawSSerum ferritin iron, a new test, measures human body iron stores unconfounded by inflammationStem Cells19971542912969253113
- PowellLWHallidayJWCowlishawJLRelationship between serum ferritin and total body iron stores in idiopathic haemochromatosisGut1978196538542680587
- RauscherLExpertDMatzankeBFTrautweinAXChrysobactin-dependent iron acquisition in Erwinia chrysanthemi. Functional study of a homolog of the Escherichia coli ferric enterobactin esteraseJ Biol Chem200227742385239511694506
- BrochuVGrenierDNakayamaKMayrandDAcquisition of iron from human transferrin by Porphyromonas gingivalis: a role for Arg- and Lys-gingipain activitiesOral Microbiol Immunol2001162798711240860
- MillerRARasmussenGTCoxCDBritiganBEProtease cleavage of iron-transferrin augments pyocyanin-mediated endothelial cell injury via promotion of hydroxyl radical formationInfect Immun19966411821888557338
- WolzCHohlochKOcaktanAIron release from transferrin by pyoverdin and elastase from Pseudomonas aeruginosaInfect Immun1994629402140278063422
- BrinesRDBrockJHThe effect of trypsin and chymotrypsin on the in vitro antimicrobial and iron-binding properties of lactoferrin in human milk and bovine colostrum. Unusual resistance of human apolactoferrin to proteolytic digestionBiochim Biophys Acta198375932292356349699
- SantosJAGonzalezCJLopezTMOteroAGarcia-LopezMLHemolytic and elastolytic activities influenced by iron in Plesiomonas shigelloidesJ Food Prot199962121475147710606155
- SokolPACoxCDIglewskiBHPseudomonas aeruginosa mutants altered in their sensitivity to the effect of iron on toxin A or elastase yieldsJ Bacteriol198215127837876807961
- GraziadeiIKahlerCMWiedermannCJVogelWThe acute-phase protein alpha 1-antitrypsin inhibits transferrin-receptor binding and proliferation of human skin fibroblastsBiochim Biophys Acta1998140121701769531972
- WeissGGraziadelIUrbanekMGrunewaldKVogelWDivergent effects of alpha 1-antitrypsin on the regulation of iron metabolism in human erythroleukaemic (K562) and myelomonocytic (THP-1) cellsBiochem J1996319 Pt 38979028920996
- GraziadeiIWeissGBohmAWerner-FelmayerGVogelWUnidirectional upregulation of the synthesis of the major iron proteins, transferrin-receptor and ferritin, in HepG2 cells by the acute-phase protein alpha1-antitrypsinJ Hepatol19972747167259365048
- GraziadeiIKaserbacherRBraunsteinerHVogelWThe hepatic acute-phase proteins alpha 1-antitrypsin and alpha 2-macroglobulin inhibit binding of transferrin to its receptorBiochem J1993290Pt 11091137679893
- GraziadeiIWeissGEggerCNiederwieserDPatschJRVogelWModulation of iron metabolism in monocytic THP-1 cells and cultured human monocytes by the acute-phase protein alpha1-antitrypsinExp Hematol19982611105310609766445
- KaserbacherRPropstTPropstAGraziadeiIJudmaierGVogelWAssociation between heterozygous alpha 1-antitrypsin deficiency and genetic hemochromatosisHepatology19931837077088359811
- BruntEMTavillASBaconBRA 49-year-old man with alpha 1-antitrypsin deficiency and abnormal iron study resultsSemin Liver Dis1996161971018723327
- GhioAJDisruption of iron homeostasis and lung diseaseBiochim Biophys Acta20091790773173919100311
- MayerASStollerJKBucher BartelsonBJames RuttenberASandhausRANewmanLSOccupational exposure risks in individuals with PI*Z alpha(1)-antitrypsin deficiencyAm J Respir Crit Care Med20001622 Pt 155355810934086
- BanauchGIBrantlyMIzbickiGAccelerated spirometric decline in New York City firefighters with alpha-antitrypsin deficiencyChest201013851116112420634282
- WoodAMHarrisonRMSempleSAyresJGStockleyRAOutdoor air pollution is associated with disease severity in alpha1-antitrypsin deficiencyEur Respir J200934234635319324958