 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Cortical spreading depression (CSD) involves a slowly-propagating depolarization wave in the cortex, which can appear in numerous pathophysiological conditions, such as migraine with aura, stroke, and traumatic brain injury. Neurons and glial cells are also depolarized transiently during the phenomena. CSD is followed by a massive increase in glutamate release and by changes in the brain microcirculation. The aim of this study was to investigate the effects of two N-methyl-D-aspartate receptor antagonists, endogenous kynurenic acid (KYNA) and dizocilpine, on CSD and the related blood–brain barrier (BBB) permeability in rats. In intact animals, KYNA hardly crosses the BBB but has some positive features as compared with its precursor L-Kynurenine, which is frequently used in animal studies (KYNA cannot be metabolized to excitotoxic agents such as 3-hydroxy-L-kynurenine and quinolinic acid). We therefore investigated the possible effects of peripherally administered KYNA. Repetitive CSD waves were elicited by the application of 1 M KCl solution to the cortex. Direct current-electrocorticograms were measured for 1 hour. Four parameters of the waves were compared. Evans blue dye and fluorescent microscopy were used to study the possible changes in the permeability of the BBB. The results demonstrated that N-methyl-D-aspartate receptor antagonists can reduce the number of CSD waves and decrease the permeability of the BBB during CSD. These results suggest that KYNA itself or its derivatives may offer a new approach in the therapy of migraines.
Introduction
Cortical spreading depression (CSD), discovered by Leao in 1944, is a slowly-propagating, transient, negative direct current (DC) shift accompanied by depressed neuronal bioelectrical activity. The main event during CSD is the depolarization of a critical mass of brain tissue and a massive elevation in glutamate release associated with an increase in extracellular potassium ion concentration, water influx into the cells, and shrinkage of the extracellular spaceCitation1–Citation4 There are also changes in immediate early genes, neurotransmitters, growth factors, etc.Citation5
Some years ago, it was observed that a high glutamate level reduced the function of the blood–brain barrier (BBB) via endothelium-expressed N-methyl-D-aspartate receptor (NMDAR)-dependent occludin phosphorylation.Citation6 Moreover, CSD initiates disruption of the BBB via a matrix metalloprotease-dependent mechanism.Citation7
Despite intensive research, the exact pathomechanism of migraine and its connection with CSD are not completely understood. Various results have indicated that CSD is associated with changes in the diameter of blood vessels in the cortical surface,Citation8 while other authors have described it as a phenomenon following acute cortical injury, including brain ischemia,Citation9 and yet others consider it to be an electrophysiological substrate of migraine aura, which may induce headache.Citation10,Citation11
In recent years, we have studied the effects of L-Kynurenine (KYN) and some derivatives of kynurenic acid (KYNA) on different animal models of migraine.Citation12–Citation14 It was found, for instance, that KYN suppressed CSD waves and could mitigate some of the labeling thought to be experimental signs of migraine (eg, c-fos immunoreactivity and neuronal nitric oxide synthase).Citation12–Citation15 It is widely accepted that KYN (the precursor of KYNA) can readily cross the BBB, but that KYNA itself, the only known endogenous NMDAR antagonist, hardly crosses the BBB at all,Citation16 whereas there is evidence that peripherally administered KYNA can penetrate the intact BBB.Citation17,Citation18 Furthermore, published data indicate that CSD results in a disruption of the BBB.Citation7,Citation19 Since NMDARs play a decisive role in the generation and propagation of CSD,Citation3,Citation20 and data are accumulating which suggest that endogenous KYNA may act as a modulator at various levels of the pathomechanism of migraine,Citation21 it seemed plausible to study the effects on CSD of exogenously administered KYNA.
Although little is yet known about the connections between migraine, CSD, and changes in BBB permeability, such a relationship appears more than probable. Accordingly, we have now studied the KCl-induced CSD and changes in BBB permeability after the administration of KYNA and, as a positive control, dizocilpine (MK-801; a well-known noncompetitive antagonist of NMDARs) in order to elucidate whether or not these neuroactive molecules act in parallel on CSD and BBB permeability.
Materials and methods
Animals
The National Institutes of Health principles of animal care and the protocol for animal care, approved by both the Hungarian Health Committee (1998) and the European Communities Council Directive (86/609/EEC), were strictly followed. Every effort was made to limit the number of animals and their suffering. Fifteen male Wistar rats (270 ± 21 g) (Charles River, Wilmington, MA, USA) were used for the electrophysiological portion of this study, and an additional 17 for histology. The animals were kept under 12-hour light and 12-hour dark conditions and were raised with free access to water and food pellets. The room temperature was 22°C ± 1°C.
Surgery
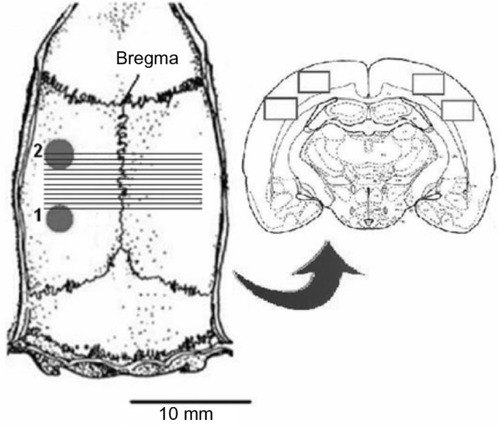
All the procedures were carried out under deep nembutal anesthesia, achieved with a dose of 60 mg/kg Euthasol® (AST Farma, Oudewater, the Netherlands), followed by 20 mg/kg hourly. A trachea cannula was inserted and the head of the animals was fixed in a stereotaxic frame (David Kopf Instruments, Tujunga, CA, USA). After exposure of the skull, two holes were drilled through the left side of the skull under cooling with saline; one hole facilitated DC-electrocorticogram (DC-ECoG) recording above the primary somatosensory cortex (3 mm caudally; 5 mm laterally from the bregma) and the other allowed access for the induction of CSD (7 mm caudally and 5 mm laterally) (). The dura mater and arachnoidea were carefully removed from the posterior hole.
Figure 1 Explanatory figure relating to the experimental protocol.
Abbreviation: KCl, potassium chloride.
Electrophysiology
A glass microelectrode filled with 150 mM saline (resistance 0.8–1.2 MΩ) connected to an Ag/AgCl wire was inserted into the cortex to a depth of 1000–1200 μm, corresponding to cortical layer V The control group (n = 5) received 1 mL of phosphate buffer (0.2 M, pH = 7.4). The KYNA group (n = 5) was treated with 300 mg/kg KYNA (Sigma-Aldrich, St Louis, MO, USA) dissolved in 400 μL of 1 M NaOH and 600 μL of phosphate buffer. MK-801 (Tocris Bioscience, Bristol, UK) was dissolved in phosphate buffer and administered at 2 mg/kg to the animals in the positive control group (n = 5).
All of these drugs were injected intraperitoneally. The cortex was allowed to recover for 30 minutes following the surgical preparation and treatment. CSD was induced by placing a cotton ball (diameter 1 mm) soaked with 1 M KCl solution on the cortex through the posterior hole. To keep the cotton ball moist, it was impregnated with 5 μL of KCl solution every 15 minutes. The DC-ECoG was measured for 1 hour. Following offline data processing with laboratory-developed software, four parameters of the CSD waves were compared between groups: the numbers of CSD episodes per hour, the peak-to-peak amplitude of the waves, the time at half-amplitude, and the area of the waves, calculated via integration. All of these parameters were measured for all waves and then averaged within an animal. The two-sample t-test statistical analyses were carried out with Origin 8.1 software (OriginLab, Northampton, MA, USA).
Histology
For the histological part of the study, the surgical preparation and treatment were the same as described above (n = 5 per group). After the 30-minute recovery, 5 mL/kg of 2% Evans blue (EB) dye, dissolved in saline, was administered intravenously at 0.1 mL/min. CSD waves were evoked by the same method as described above. Two additional animals served as sham-operated controls. The sham preparation was similar to that for experimental animals except that a saline-impregnated cotton ball was applied to the cortex. After 1 hour induction of CSD by KCl or saline irrigation, the animals were perfused with cold saline and fixed with 4% paraformaldehyde dissolved in 0.1 M phosphate buffer. The brains were removed and postfixed overnight in the same paraformaldehyde solution. Sections were cut at a thickness of 20 μm with a vibratome (Leica VT 1000S; Leica Microsystems, Wetzlar, Germany). Twelve sections were cut in steps of 500 μm from the area of CSD initiation, in the rostral direction. The first section was situated 6 mm caudally from the bregma. These preparations were mounted on 2% gelatine-coated slides and covered with Fluoromount (SERVA, Heidelberg, Germany). The slides were examined under a fluorescent microscope (Olympus BX51; Olympus Corporation, Tokyo, Japan) equipped with a DP70 digital imaging system (Olympus Corporation). A Texas red filter (532–587 nm excitation wavelength and 608–683 nm emission wavelength) was used for the detection of EB fluorescence. Two 1360 × 1024 red green, blue color model pictures were captured from each hemisphere of the sections (). After data collection, the pictures were transformed to 8-bit gray-scale images, and the mean intensity was calculated via the following equation:
where I is the intensity of pixels, n is the number of pixels in a row and m is the number of pixels in a column. After quantification, the ratio of the intensities of the two hemispheres was calculated and plotted (). Repeated-measures one-way analysis of variance (ANOVA) was carried out with Origin 8.1 software.
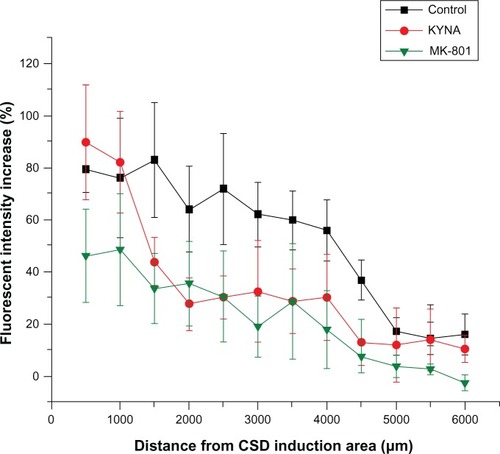
Figure 2 Fluorescence intensity changes caused by plasma extravasation in the three groups.
Abbreviations: ANOVA, analysis of variance; CSD, cortical spreading depression; KYNA, kynurenic acid; MK-801, dizocilpine; KCl, potassium chloride.
Results
Electrophysiology
KCl administration to the cortex resulted in CSD waves with a frequency of 15 ± 1.1/hour (mean ± standard error of the mean). KYNA suppressed the CSD activity significantly (9.8/hour ± 0.9/hour; P = 0.0084), while MK-801 administration completely eliminated the CSD waves ().
Figure 3 Diagrams of cortical spreading depression (CSD) wave parameters.
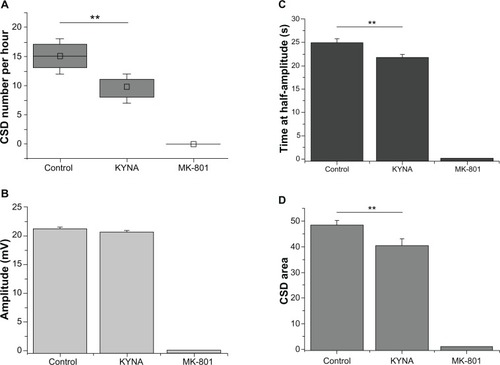
Detailed data analysis showed that the application of 1 M KCl to the cortex in the control group resulted in CSD waves with an amplitude of 21 mV ± 0.3 mV However, KYNA (20.6 mV ± 0.7 mV) did not cause a significant change in the amplitude of the CSD waves, and no CSD wave was observed following the administration of MK-801 ().
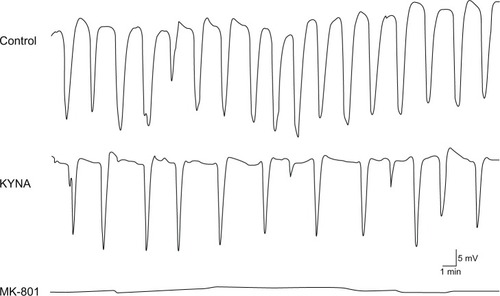
Though KYNA did not induce a significant change in the amplitude of the CSD waves, the time at half-amplitude was significantly shorter after KYNA administration (from 24.9 seconds ± 0.7 seconds to 21.7 seconds ± 0.6 seconds; P = 0.0014) (). This is because the CSD waves became sharper as compared with the controls after drug administration ().
Figure 4 Representative cortical spreading depression (CSD) waves recorded in the barrel cortex.
After integration of the waves, the CSD area in the control group was 48.3 ± 1.7. The KYNA treatment produced a significant decline, to 40.1 ± 2.4 (P = 0.008). MK-801 completely eliminated the CSD activity ().
Histology
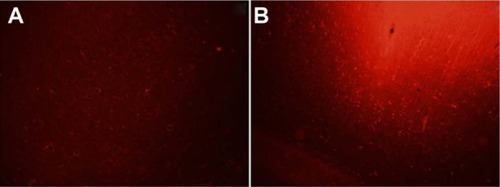
There were no detectable differences between the two hemispheres after 1 hour of saline irrigation in the sham-operated control group (n = 2). In all groups, KCl was administered to the cortex, as detailed above. In the control and the KYNA-treated group, massive plasma extravasation was detected as intense EB fluorescence within 1000 μm of the site of KCl application (). The only exception was the MK-801 group, where the level of fluorescence (extravasation) was half that in the control and KYNA groups. However, the intensity of fluorescence decreased with increasing distance from the site of CSD induction. Fluorescence in the KYNA-treated animals was only reduced to the levels observed with MK-801 treatment at ∼1500 μm (). Repeated-measures one-way ANOVA did not reveal any significant difference between the groups.
Figure 5 Representative photomicrographs of the cerebral cortex.
Abbreviation: KCl, potassium chloride.
Discussion
The mechanism of generation of CSD waves, and especially those aspects relating to pathological conditions such as migraine, brain ischemia, and alterations in the BBB, is not yet clear.
A decade ago, it was reported that ischemia and hypoxic cortical depolarization are associated with an intense vasospasm and vasogenic edema.Citation19 It was later found that neuronal and glial depolarization and an increase in extracellular glutamate level initiate a cascade that disrupts the BBB in numerous ways.Citation6,Citation7
We have previously investigated the effects of kynurenines on different animal models of migraine. In all of the studied models, KYN (a precursor of KYNA) and other KYNA derivatives proved effective in mitigating the stimulation-induced increase in c-fos immunoreactivity in the caudal trigeminal nucleusCitation12,Citation13 or nitroglycerine-induced neuronal nitric oxide synthase.Citation14
It was recently described that the administration of KYN significantly decreases the frequency of CSD, an effect due to the increased level of KYNA in the brain.Citation15 Although KYN seems to be neuroprotective in many animal models of migraine and stroke, it sometimes facilitates pathological pathways (our unpublished data).Citation22 This may be caused by the metabolization of KYN; it can be converted to excitotoxic 3-hydroxy-L-kynurenine and quinolinic acid in the brain.Citation23 Kynurenine aminotransferase converts KYN to KYNA in a unidirectional manner, and peripherally administered KYNA can therefore not be metabolized to excitotoxic agents.Citation24 Accordingly, and in light of the fact that CSD increases the permeability of the BBB, it appeared worthwhile to investigate whether systemically administered KYNA influences the CSD activity. In parallel with KYNA, we studied the effects of MK-801 as a positive control.
The aims of this study were to learn whether or not peripherally administered KYNA is able to influence the CSD activity, and whether these neuroactive molecules (which influence the glutamatergic system) act in parallel on CSD and the permeability of the BBB. Our results using these molecules were in accordance with earlier observations and inferences that glutamate is involved in CSD.Citation25,Citation26 Results of this study also implicated elements of the pathophysiology of the disorder, including trigeminovascular activation and central sensitization.Citation27
The fluorescent dye EB binds to the serum albumin in the blood. Under physiological conditions, albumin cannot pass through the brain capillaries. The intracellular calcium concentration [Ca2+]i demonstrates a laminar difference in the response of the pyramidal neurons to CSD.Citation28 Unfortunately, the situation concerning the [Ca2+]i in the endothelial cells is not known, but in general, the linkage between [Ca2+]i, nitrous oxide, and the integrity of the BBB is well-based:Citation29,Citation30 elevated [Ca2+]i and enhanced neuronal nitric oxide synthase activation lead to robust vasodilatation and protein extravasation, when the albumin-EB complex may appear in the brain tissue following KCl-induced CSD activity.
Although it cannot be excluded that KCl administration itself may result in increased extravasation, the fact that KYNA and MK-801 administration resulted in reduced or eliminated CSD activity in parallel with decreased extravasation strongly suggests that the waves of cortical depolarization and the level of BBB permeability are related.
CSD waves, the typical electrophysiological signs of cortical depolarizing waves, disappeared completely after MK-801 administration. A similar, but less pronounced effect was observed after KYNA administration. The changes in the electrical signal of the CSD (ie, the decrease in frequency of the DC shifts and the sharpening of the waves) were observed at the beginning of recording, but CSD was elicited only at the beginning of the DC-ECoG measurement. This suggests that peripherally administered KYNA is able to cross the intact BBB and change the parameters of the waves. In parallel with the electrophysiological results, the administration of these NMDAR antagonists resulted in a gradually declining level of fluorescence, in accordance with increasing distance from the source of the CSD waves (the site of KCl application).
The reason for this relatively moderate effect of KYNA, as compared with that of MK-801, is probably its increased but limited permeability through the BBB. Under physiological conditions, KYNA is barely able to cross the BBB,Citation16 but our results indicate that, in KCl-treated animals, KYNA may pass through the BBB and influence the CSD activity. KYNA presumably mitigates the effects of the glutamate system. KYNA is not only an endogenous NMDAR blocker, but also a noncompetitive inhibitor of the α-7-nicotinic acetylcholine receptor,Citation31,Citation32 and through this mechanism, KYNA can decrease glutamate release.Citation33 In this manner, KYNA can reduce the pathological glutamate level in the cortex during CSD activity and protect the BBB structure.Citation6
Our results suggest that, in the course of CSD-induced glutamate release, the permeability of the BBB increases, and systemically administered KYNA can therefore reduce the consequences of CSD.
Recent findings indicated that kynurenines, and particularly KYNA, exhibit a strong modulatory potential on the neuronal structures in the brainstem, which may play a crucial role in the pathogenesis of migraine.Citation34 The present data reveal that KYNA may also act in the cortex. These results provide an experimental tool by which to understand the pathomechanism of migraine. However, further studies are needed to clarify the possible role of the kynurenines in the therapy of migraine.
Acknowledgments
This work was supported by TÁMOP 4.2.2-A-11/KONV-2012-0052, OTKA K105077 and TÁMOP 4.2.4. A/2-11-1-2012-0001 “National Excellence Program – Elaborating and operating an inland student and researcher personal support system”.
Disclosure
The authors report no conflicts of interest in this work.
References
- Gardner-Medwin AR Possible roles of vertebrate neuroglia in potassium dynamics, spreading depression and migraine J Exp Biol 1981 95 111 127 7334315
- Kraig RP Nicholson C Extracellular ionic variations during spreading depression Neuroscience 1978 3 11 1045 1059 745780
- Somjen GG Mechanisms of spreading depression and hypoxic spreading depression-like depolarization Physiol Rev 2001 81 3 1065 1096 11427692
- Fabricius M Jensen LH Lauritzen M Microdialysis of interstitial amino acids during spreading depression and anoxic depolarization in rat neocortex Brain Res 1993 612 1–2 61 69 8330214
- Jander S Schroeter M Peters O Witte OW Stoll G Cortical spreading depression induces proinflammatory cytokine gene expression in the rat brain J Cereb Blood Flow Metab 2001 21 3 218 225 11295876
- András IE Deli MA Veszelka S Hayashi K Hennig B Toborek M The NMDA and AMPA/KA receptors are involved in glutamate-induced alterations of occludin expression and phosphorylation in brain endothelial cells J Cereb Blood Flow Metab 2007 27 8 1431 1443 17245419
- Gursoy-Ozdemir Y Qiu J Matsuoka N Cortical spreading depression activates and upregulates MMP-9 J Clin Invest 2004 113 10 1447 1455 15146242
- Brennan KC Beltrán-Parrazal L López-Valdés HE Theriot J Toga AW Charles AC Distinct vascular conduction with cortical spreading depression J Neurophysiol 2007 97 6 4143 4151 17329631
- Dohmen C Sakowitz OW Fabricius M Spreading depolarizations occur in human ischemic stroke with high incidence Ann Neurol 2008 63 6 720 728 18496842
- Santos E Sánchez-Porras R Dohmen C Hertle D Unterberg AW Sakowitz OW Spreading depolarizations in a case of migraine-related stroke Cephalalgia 2012 32 5 433 436 22407661
- Zhang X Levy D Noseda R Kainz V Jakubowski M Burstein R Activation of meningeal nociceptors by cortical spreading depression: implications for migraine with aura J Neurosci 2010 30 26 8807 8814 20592202
- Knyihár-Csillik E Toldi J Krisztin-Péva B Prevention of electrical stimulation-induced increase of c-fos immunoreaction in the caudal trigeminal nucleus by kynurenine combined with probenecid Neurosci Lett 2007 418 2 122 126 17382471
- Knyihár-Csillik E Toldi J Mihály A Kynurenine in combination with probenecid mitigates the stimulation-induced increase of c-fos immunoreactivity of the rat caudal trigeminal nucleus in an experimental migraine model J Neural Transm 2007 114 4 417 421 16897600
- Vámos E Párdutz A Varga H l-kynurenine combined with probenecid and the novel synthetic kynurenic acid derivative attenuate nitroglycerin-induced nNOS in the rat caudal trigeminal nucleus Neuropharmacology 2009 57 4 425 429 19580819
- Chauvel V Vamos E Párdutz A Vecsei L Schoenen J Multon S Effect of systemic kynurenine on cortical spreading depression and its modulation by sex hormones in rat Exp Neurol 2012 236 2 207 214 22587906
- Fukui S Schwarcz R Rapoport SI Takada Y Smith QR Blood-brain barrier transport of kynurenines: implications for brain synthesis and metabolism J Neurochem 1991 56 6 2007 2017 1827495
- Heyes MP Quearry BJ Quantification of kynurenic acid in cerebrospinal fluid: effects of systemic and central L-kynurenine administration J Chromatogr 1990 530 1 108 115 2177481
- Scharfman HE Goodman JH Effects of central and peripheral administration of kynurenic acid on hippocampal evoked responses in vivo and in vitro Neuroscience 1998 86 3 751 764 9692715
- Koroleva VI Vinogradova LV [Ischemic and hypoxic depolarization in the rat neocortex.] Zh Vyssh Nerv Deiat Im I P Pavlova 2000 50 4 612 623 Russian [with English abstract] 10984906
- Lauritzen M Hansen AJ The effect of glutamate receptor blockade on anoxic depolarization and cortical spreading depression J Cereb Blood Flow Metab 1992 12 2 223 229 1312539
- Fejes A Párdutz A Toldi J Vécsei L Kynurenine metabolites and migraine: experimental studies and therapeutic perspectives Curr Neuropharmacol 2011 9 2 376 387 22131946
- Roussel S Pinard E Seylaz J Kynurenate does not reduce infarct size after middle cerebral artery occlusion in spontaneously hypertensive rats Brain Res 1990 518 1–2 353 355 2390725
- Schwarcz R Pellicciari R Manipulation of brain kynurenines: glial targets, neuronal effects, and clinical opportunities J Pharmacol Exp Ther 2002 303 1 1 10 12235226
- Turski WA Schwarcz R On the disposition of intrahippocampally injected kynurenic acid in the rat Exp Brain Research 1988 71 3 563 567
- Longoni M Ferrarese C Inflammation and excitotoxicity: role in migraine pathogenesis Neurol Sci 2006 27 Suppl 2 S107 S110 16688611
- Ramadan NM The link between glutamate and migraine CNS Spectr 2003 8 6 446 449 12858134
- Andreou AP Goadsby PJ Therapeutic potential of novel glutamate receptor antagonists in migraine Expert Opin Investig Drugs 2009 18 6 789 803
- Gniel HM Martin RL Changes in membrane potential and the intracellular calcium concentration during CSD and OGD in layer V and layer II/III mouse cortical neurons J Neurophysiol 2010 104 6 3203 3212 20810684
- Chen Y McCarron RM Azzam N Endothelin-1 and nitric oxide affect human cerebromicrovascular endothelial responses and signal transduction Acta Neurochir Suppl 2000 76 131 135 11449992
- Thiel VE Audus KL Nitric oxide and blood-brain barrier integrity Antioxid Redox Signal 2001 3 2 273 278 11396481
- Hilmas C Pereira EF Alkondon M Rassoulpour A Schwarcz R Albuquerque EX The brain metabolite kynurenic acid inhibits alpha7 nicotinic receptor activity and increases non-alpha7 nicotinic receptor expression: physiopathological implications J Neurosci 2001 21 19 7463 7473 11567036
- Stone TW Forrest CM Mackay GM Stoy N Darlington LG Tryptophan, adenosine, neurodegeneration and neuroprotection Metab Brain Dis 2007 22 3–4 337 352 17712616
- Konradsson-Geuken A Wu HQ Gash CR Cortical kynurenic acid bi-directionally modulates prefrontal glutamate levels as assessed by microdialysis and rapid electrochemistry Neuroscience 15 169 4 1848 1859
- Tajti J Szok D Párdutz A Where does a migraine attack originate? In the brainstem J Neural Transm 2012 119 5 557 568 22426834