Abstract
Glycine is a nonessential amino acid that is reversibly converted from serine intracellularly by serine hydroxymethyltransferase. Glyphosate and its degradation product, aminomethylphosphonic acid (AMPA), are analogs to glycine, thus they may inhibit serine hydroxymethyltransferase to decrease intracellular glycine synthesis. In this study, we found that glyphosate and AMPA inhibited cell growth in eight human cancer cell lines but not in two immortalized human normal prostatic epithelial cell lines. AMPA arrested C4-2B and PC-3 cancer cells in the G1/G0 phase and inhibited entry into the S phase of the cell cycle. AMPA also promoted apoptosis in C4-2B and PC-3 cancer cell lines. AMPA upregulated p53 and p21 protein levels as well as procaspase 9 protein levels in C4-2B cells, whereas it downregulated cyclin D3 protein levels. AMPA also activated caspase 3 and induced cleavage of poly (adenosine diphosphate [ADP]-ribose) polymerase. This study provides the first evidence that glyphosate and AMPA can inhibit proliferation and promote apoptosis of cancer cells but not normal cells, suggesting that they have potentials to be developed into a new anticancer therapy.
Introduction
Glycine is a nonessential amino acid that can be converted directly from serine in mammalian cells. Serine is derived from 3-phosphoglycerate. Serine hydroxymethyltransferase (SHMT) reversibly catalyzes the conversion of serine to glycine and vice versa. In the liver of vertebrates, glycine can be synthesized from N5,N10-methylene tetrahydrofolate by glycine synthase.Citation1 Glycine is the precursor material for biosynthesis of protein, purine, and glutathione. In addition, glycine is converted into sarcosine (N-methylglycine) by glycine N-methyltransferase,Citation2 and reversely, sarcosine can be converted into glycine by sarcosine dehydrogenase.Citation3 The serum half-life of intravenously administered glycine varies from 1/2 hour to 4 hours.Citation4 Recently, the importance of glycine in cancer cell biology has been revealed.Citation5 In a human melanoma LOX IMVI cell line, about two-thirds of intracellular glycine is synthesized intracellularly, while one-third of intracellular glycine is taken from extracellular glycine sources. In a panel of 60 human cancer cell lines that the National Cancer Institute uses for anticancer drug screening, rapidly proliferating cancer cells consumed extracellular glycine due to increased demand for glycine. In contrast, rapidly proliferating human normal cell lines released glycine.Citation5 These findings suggest that targeting glycine consumption may become a new strategy in killing rapidly proliferating cancer cells, while not harming rapidly proliferating normal cells. Indeed, the use of short hairpin RNA (shRNA) to knockdown expression of SHMT2, thus blocking endogenous glycine synthesis, effectively halted proliferation of LOX IMVI cells in the absence of extracellular glycine. In contrast, slowly proliferating cells were not affected by SHMT2 knockdown and deprivation of extracellular glycine.Citation5
There are two isozymes of SHMT. SHMT1 encodes for the cytoplasmic and SHMT2 encodes for the mitochondrial isozyme.Citation6–Citation8 In mammalian cells, SHMT2 gene has an alternative promoter within intron 1, thus SHMT2 encodes for two transcripts, SHMT2 and SHMT2α.Citation9 SHMT2 protein containing exon 1 (with mitochondrial-targeting sequence) is localized in mitochondria. SHMT2α protein without exon 1 is not imported into mitochondria efficiently and is localized predominantly in the cytoplasm and nucleus. SHMT1 protein, like SHMT2α, is also localized in the cytoplasm and nucleus, and both SHMT1 and SHMT2α catalyze production of one-carbon units from serine for nuclear de novo thymidylate biosynthesis.Citation9 Interestingly, a glycine analog, aminomethylphosphonate (aminomethylphosphonic acid [AMPA]) (molecular formula CH6NO3P []), inhibits more than 95% of nuclear thymidylate biosynthesis that requires SHMT1 and SHMT2α, suggesting that AMPA is an effective inhibitor of SHMT1 and SHMT2α, as well as SHMT2.Citation9
Figure 1 Chemical structure of glycine, AMPA, and glyphosate.
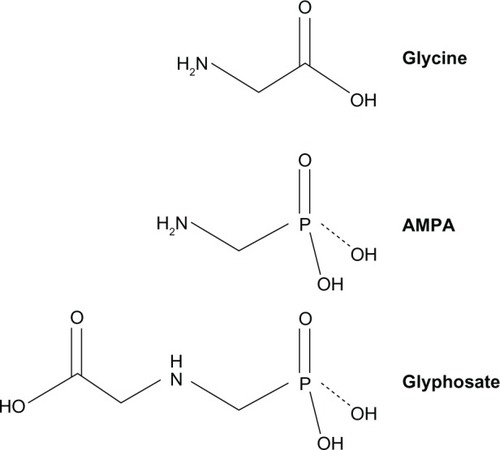
AMPA is the primary degradation product of glyphosate (N-(phosphonomethyl)glycine; molecular formula C3H3NO5P []). Glyphosate is water soluble and chemically stable and is degraded by microbes to produce AMPA.Citation10 Glyphosate is a weak organic acid consisting of a glycine moiety and a phosphonomethyl moiety. Glyphosate is a broad-spectrum herbicide that is used worldwide in agriculture, forestry, and aquatic weed control. It is applied to many crops in various commercial formulations. The major formulation is Roundup® (Monsanto Co, St Louis, MO, USA), in which glyphosate is formulated as the isopropylamine salt,Citation11 whereas AMPA has no commercial use.Citation10 In animal studies using [14C]glyphosate in rats, rabbits, and goats, approximately 30% of the oral dose was absorbed through the gastrointestinal tract. On day 7 after the oral dose of [14C]glyphosate in rats, the isotope was distributed throughout the animal body, with the highest concentration found in the bones. Almost all of the isotope was eliminated in urine and feces, with a very low level exhaled in air. The only metabolite was AMPA, which accounted for about 0.2% to 0.3% of the administered dose of glyphosate.Citation10 In a study in rats, approximately 20% of the oral dose of AMPA was absorbed, which was excreted almost exclusively through the urine, with less than 0.1% of the dose expired as CO2.Citation10 Glyphosate and AMPA have been found to present no significant toxicity in acute, subchronic, and chronic animal studies, nor any genotoxicity, teratogenicity, or carcinogenicity.Citation10,Citation12 In this study, we assessed the effects of glyphosate and AMPA on cancer cell growth. Our results suggest that glyphosate and AMPA inhibit cell growth in eight cancer cell lines but not in two immortalized human normal prostatic epithelial cell lines, at concentrations up to 50 mM.
Materials and methods
Cell culture
The immortalized human normal prostatic epithelial cell lines RWPE-1 and pRNS-1-1 were obtained from Johng S Rhim (Uniformed Services University of the Health Sciences, Bethesda, MD, USA).Citation13 Human castration-resistant prostate cancer cell line C4-2B was obtained from Leland WK Chung (Cedars-Sinai Medical Center, Los Angeles, CA, USA).Citation14 Human prostate cancer cell lines (LNCaP, DU-145, and PC-3), human ovarian cancer cell lines (SKOV-3 and OVCAR-3), human cervical cancer HeLa cell line, and human lung cancer A549 cell line were purchased from the American Type Culture Collection (Manassas, VA, USA). RWPE-1 and pRNS-1-1 cells were cultured in Dulbecco’s Modified Eagle’s medium (DMEM) (Mediatech, Inc, Manassas, VA, USA) containing 10% fetal bovine serum (FBS) (HyClone Standard Fetal Bovine Serum; Thermo Fisher Scientific Inc, Waltham, MA, USA), 100 IU/mL penicillin/streptomycin, 5 μg/mL bovine insulin, 25 μg/mL bovine pituitary extract, and 6 ng/mL recombinant human epidermal growth factor (Sigma-Aldrich Corp, St Louis, MO, USA). LNCaP cells were cultured in T-Medium (Life Technologies Corp, Carlsbad, CA, USA) containing 5% FBS and 100 IU/mL penicillin/streptomycin. C4-2B and SKOV-3 cells were cultured in Roswell Park Memorial Institute (RPMI)-1640 medium (Thermo Fisher Scientific) containing 10% FBS and 100 IU/mL penicillin/streptomycin. OVCAR-3 cells were cultured in RPMI-1640 medium containing 20% FBS, 100 IU/mL penicillin/streptomycin, 5 μg/mL bovine insulin, and 1 mM sodium pyruvate. DU-145, PC-3, HeLa, and A549 cells were cultured in DMEM medium containing 10% FBS and 100 IU/mL penicillin/streptomycin. DMEM contains 30 mg/L glycine and 42 mg/L L-serine; RPMI-1640 medium contains 10 mg/L glycine and 30 mg/L L-serine; and T-Medium contains 27 mg/L glycine and 37.8 mg/L L-serine. The cells were cultured in a 5% CO2 humidified incubator at 37°C.
Cell viability assay
The live cell numbers were determined using the CellTiter-Glo® Luminescent Cell Viability Assay (Promega Corp, Fitchburg, WI, USA). This assay is based on quantitation of the adenosine triphosphate (ATP), an indicator of metabolically active cells, which is a well-established method for cell proliferation and cytotoxicity assays.Citation15–Citation17 To optimize the experimental conditions, we performed pilot experiments and determined that there was a linear relationship (r Citation2 = 0.98) between the luminescent signal and the number of cells, from 1,000 to 40,000 cells per well. Therefore, we plated 4,000 cells per well, so that the cell number was less than 40,000 cells per well after 72 hours in culture. The cells were plated in 100 μL complete culture medium with FBS in Costar® opaque-walled 96-well plates (Thermo Fisher Scientific Inc). After overnight incubation, the cells were treated with glyphosate or AMPA (Sigma-Aldrich Corp) at final concentrations of 0, 15, 25, or 50 mM for 72 hours. Both glyphosate and AMPA were used in their native forms in all the experiments, and they were dissolved in complete medium with FBS prior to use. Each treatment group had triplicate wells. Wells containing the same medium without cells were used to assess the background luminescence. The amount of 100 μL CellTiter-Glo® Reagent was added into each well according to the instructions. Luminescence was read using a FLUOstar OPTIMA (BMG Labtech GmbH, Ortenberg, Germany) microplate reader. Cell viability was calculated as (luminescence of the treatment group − background luminescence) ÷ (luminescence of the control group − background luminescence) × 100%. The data are presented as the mean and standard error of the mean (SEM) of three independent experiments.
Cell cycle analysis
C4-2B and PC-3 cells were plated in complete culture medium with FBS, into 60 mm dishes. After 16 hours, the cells were treated without or with AMPA at a final concentration of 50 mM for 24 hours. The cells were trypsinized washed once with phosphate buffered saline (PBS), pelleted and resuspended in 70% ice-cold ethanol and stored at −20°C until the cell cycle analysis. The fixed cells were stained in a solution containing 0.1% Triton™ X-100 (Sigma-Aldrich Corp), 0.2 mg/mL DNase-free RNase A, and 20 μg/mL propidium iodide for 30 min at room temperature in the dark. The percentages of cells at G1/G0, S, and G2/M phases were determined by flow cytometry analysis using DNA content frequency histogram deconvolution software (Becton, Dickinson and Company, Franklin Lakes, NJ, USA).
Cell apoptosis assay
C4-2B and PC-3 cells were plated in complete culture medium with FBS into 60 mm dishes. Sixteen hours later, the cells were treated without or with AMPA at a final concentration of 50 mM for 0, 24, 48, and 72 hours. The cells were trypsinized washed once with PBS, pelleted and resuspended in annexin-binding buffer. The cells were stained with Annexin-V Fluorescein isothiocyanate (FITC) Conjugate (1:20 dilution; Life Technologies Corp) and propidium iodide (100 μg/mL) according to the manufacturers protocol, and the percentages of apoptotic cells were determined by flow cytometry analysis.Citation18,Citation19
Western blot analysis
C4-2B cells were cultured in 60 mm dishes overnight. The cells were treated with AMPA at a final concentration of 50 mM for 0, 12, 24, 48, and 72 hours. Proteins were extracted from the treated cells in radioimmunoprecipitation assay (RIPA) lysis buffer (50 mM sodium fluoride, 0.5% Igepal® CA-630 [NP-40], 10 mM sodium phosphate, 150 mM sodium chloride, 25 mM Tris pH 8.0, 1 mM phenylmethylsulfonyl fluoride, 2 mM ethylenediaminetetraacetic acid [EDTA], 1.2 mM sodium vanadate) supplemented with protease inhibitor cocktail (Sigma-Aldrich Corp). An equal amount of proteins was subjected to 10% SDS-polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes (Bio-Rad Laboratories, Hercules, CA, USA). The membranes were blocked with 5% nonfat dry milk in TBST buffer (25 mM Tris-HCl, 125 mM NaCl, 0.1% Tween 20) for 2 hours and probed with the indicated primary antibodies overnight and then IRDye®800CW- or IRDye®680-conjugated secondary antibodies (LI-COR Biosciences Inc, Lincoln, NE, USA) for 1 hour. The results were visualized using an Odyssey® Infrared Imager (LI-COR Biosciences Inc). For loading control, the membranes were stripped and probed for glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The antibodies used were as follows: rabbit anti-poly (adenosine diphosphate [ADP]-ribose) polymerase (PARP), rabbit anti-caspase 3, rabbit anti-caspase 9, and mouse anti-cyclin D3 antibodies, purchased from Cell Signaling Technology Inc (Danvers, MA, USA); mouse anti-p21 and goat anti-p53 antibodies, obtained from Santa Cruz Biotechnology Inc (Dallas, TX, USA); and mouse anti-GAPDH antibodies, ordered from EMD Millipore Corp (Billerica, MA, USA).
Statistical analysis
All experiments were repeated three times, and the results represent mean ± SEM of three independent experiments. Statistical analysis was made using two-tailed Student’s t test. A P-value < 0.05 was considered statistically significant.
Results
Glyphosate inhibits cell growth in cancer cell lines but not in normal cell lines
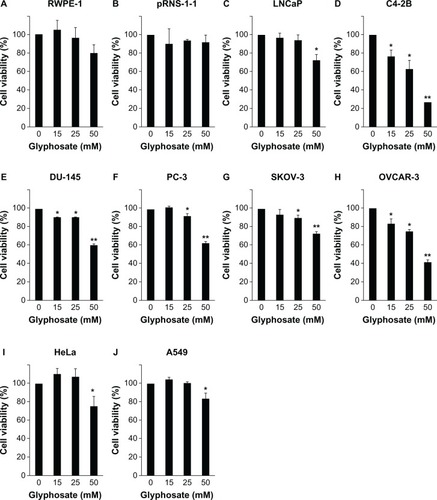
Glyphosate, at concentrations of 15, 25, and 50 mM, did not significantly decrease the cell viability in the RWPE-1 and pRNS-1-1 cell lines compared with the untreated control group (P > 0.05) (). Glyphosate, at concentrations of 15 and 25 mM, did not decrease the cell viability in the LNCaP cell line; however, it decreased 27% of the cell viability at a concentration of 50 mM (P < 0.05) (). Glyphosate, at concentrations of 15, 25, and 50 mM, significantly decreased the cell viability in the C4-2B and DU-145 cell lines (P < 0.05 or P < 0.01) (), with a 73.4% and 39.3% decrease at the dose of 50 mM, respectively. Glyphosate, at a concentration of 15 mM, did not decrease the cell viability in the PC-3 and SKOV-3 cell lines; however, it significantly decreased the cell viability at concentrations of 25 and 50 mM (P < 0.05 or P < 0.01) (), with a 36.9% and 28% decrease at the dose of 50 mM in the PC-3 and SKOV-3 cell lines, respectively. Glyphosate, at concentrations of 15, 25, and 50 mM, significantly decreased the cell viability in the OVCAR-3 cell line (P < 0.05 or P < 0.01) (), with a 58.8% decrease at the dose of 50 mM. However, at a concentration of 50 mM, glyphosate only decreased about 25% and 17% of the cell viability in the HeLa and A549 cell lines, respectively, though the decrease was statistically significant (P < 0.05) (). Based on the percentages of inhibition caused by different concentrations of glyphosate, we estimated the half maximal (50%) inhibitory concentrations (IC50) of glyphosate in the cell lines, using a linear regression model ().
Table 1 Half maximal inhibitory concentrations (IC50) of glyphosate and AMPA in inhibition of the cell growth in the normal and cancer cell lines
Figure 2 Glyphosate inhibits cell growth in cancer cell lines but not in normal cell lines.
Abbreviation: SEM, standard error of the mean.
AMPA inhibits cell growth in cancer cell lines but not in normal cell lines
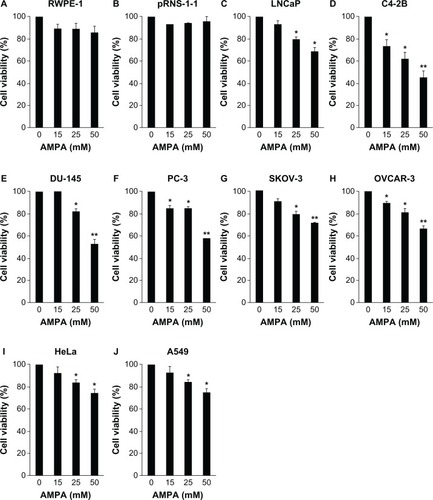
AMPA, at concentrations of 15, 25, and 50 mM, did not significantly decrease the cell viability in the RWPE-1 and pRNS-1-1 cell lines (P > 0.05) (). In contrast, AMPA, at concentrations of 25 and 50 mM, significantly decreased the cell viability in the LNCaP, DU-145, SKOV-3, HeLa, and A549 cell lines (P < 0.05 or P < 0.01) (), while AMPA at concentrations of 15, 25, and 50 mM significantly decreased the cell viability in the C4-2B, PC-3, and OVCAR-3 cell lines (P < 0.05 or P < 0.01) (). The percentages of decrease in cell viability at 50 mM AMPA were 32% in LNCaP, 54.5% in C4-2B, 47% in DU-145, 41.7% in PC-3, 28.5% in SKOV-3, 33.6% in OVCAR-3, 25% in HeLa, and 31.4% in the A549 cell lines. Of note, we found that at a concentration of 100 mM, AMPA decreased the cell viability in the RWPE-1 and pRNS-1-1 cell lines by 59.5% and 57.6%, respectively. In contrast, this high concentration of AMPA decreased cell viability by 54% in LNCaP, 91% in C4-2B, 67% in DU-145, 85.1% in PC-3, 42% in SKOV-3, 79.9% in OVCAR-3, 50.6% in HeLa, and 53.1% in the A549 cell lines. The IC50 concentrations of AMPA in inhibition of the cell growth in the normal and cancer cell lines are shown in .
Figure 3 AMPA inhibits cell growth in cancer cell lines but not in normal cell lines.
Abbreviations: AMPA, aminomethylphosphonic acid; SEM, standard error of the mean.
AMPA inhibits entry into the s phase of cell cycle and increases apoptosis
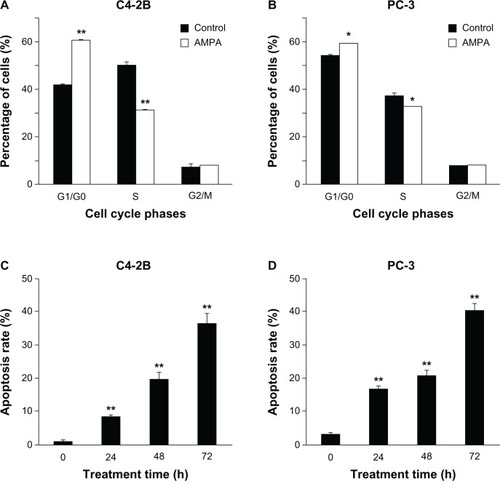
AMPA, at a concentration of 50 mM, significantly increased the number of C4-2B and PC-3 cells in the G1/G0 phase of cell cycle, compared with the control group (P < 0.05 or P < 0.01) (). In contrast, AMPA significantly decreased the number of C4-2B and PC-3 cells in the S phase of cell cycle (P < 0.05 or P < 0.01), whereas the number of cells in the G2/M phase was not affected (). In addition, AMPA, at a concentration of 50 mM, significantly increased apoptosis of C4-2B and PC-3 cells in a time-dependent manner (P < 0.01) ().
Figure 4 AMPA inhibits entry into the s phase of cell cycle and increases apoptosis.
Abbreviations: AMPA, aminomethylphosphonic acid; FITC, Fluorescein isothiocyanate; SEM, standard error of the mean.
AMPA induces changes in expression levels of genes involved in cell cycle and apoptosis
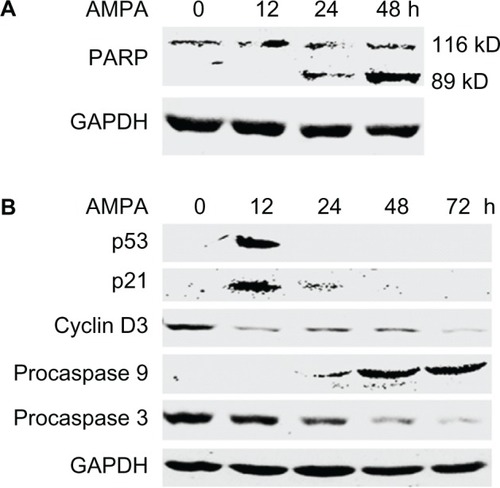
AMPA, at a concentration of 50 mM, increased the levels of cleaved PARP in the C4-2B cells in a time-dependent manner (). AMPA transiently increased the levels of p53 and its downstream gene p21, at 12 hours after treatment (). It also decreased cyclin D3 protein levels, starting from 12 hours after treatment (). Further, AMPA increased the levels of procaspase 9, starting from 24 hours after treatment (). In contrast, AMPA decreased the levels of procaspase 3, starting from 24 hours after treatment ().
Figure 5 AMPA induces changes in expression levels of genes involved in cell cycle and apoptosis.
Abbreviations: AMPA, aminomethylphosphonic acid; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; h, hours; PARP, poly(ADP-ribose) polymerase; ADP, adenosine diphosphate.
Discussion
A previous study identified glycine as being consumed by rapidly proliferating cancer cell lines. Among the 60 human cancer cell lines tested in the previous study, five were included in the present study, namely, DU-145, PC-3, SKOV-3, OVCAR-3, and A549. Previously, it was shown that cancer cell proliferation was impaired by knocking down the expression levels of SHMT2 that is responsible for intracellular glycine synthesis.Citation5 Here we present data showing that glycine analogs, glyphosate and AMPA, inhibited cell growth in eight cancer cell lines, including four human prostate cancer cell lines (LNCaP, C4-2B, DU-145, and PC-3), two human ovarian cancer cell lines (SKOV-3 and OVCAR-3), one human cervical cancer cell line (HeLa), and one human lung cancer cell line (A549). In contrast, glyphosate and AMPA did not impair the growth of two immortalized human normal prostatic epithelial cell lines (RWPE-1 and pRNS-1-1) at concentrations up to 50 mM. These findings suggest that glyphosate and AMPA can differentially affect cancer cell growth but not normal cell growth at concentrations up to 50 mM. However, at a higher concentration of 100 mM, AMPA decreased cell viability of the two normal cell lines, suggesting that potential adverse side effects may arise when the doses are too high. A safe therapeutic window may be limited to a drug concentration of between 50 mM to 100 mM. However, in clinical practice, it may be difficult to administer the drugs to reach this high concentration. Therefore, the use of glyphosate and AMPA in patients may be limited if the high IC50 concentrations are difficult to achieve in patients’ blood. Nevertheless, glyphosate, particularly AMPA, may be a good lead compound for developing more potent inhibitors with low IC50 concentrations.
Among the human cancer cell lines, the castration-resistant prostate cancer cell lines (C4-2B, DU-145, and PC-3) and ovarian cancer cell line (OVCAR-3) appear to be more sensitive to glyphosate and AMPA than are the other human cancer cell lines. The IC50 concentrations varied from 34.2 to 68.1 mM among the four sensitive cancer cell lines (C4-2B, DU-145, PC-3, and OVCAR-3). In contrast, the IC50 concentrations were between 89 and 136.7 mM in other cancer cell lines, including LNCaP, SKOV-3, HeLa, and A549 (), suggesting that these four cell lines are resistant to glyphosate and AMPA. The differences in sensitivity may be caused by many factors that are not clearly understood because these cell lines are derived from different genetic backgrounds. The rate of cell proliferation may be one of the factors, as suggested by previous study.Citation5 Indeed, the C4-2B cells were more sensitive to glyphosate and AMPA than LNCaP cells. Coincidentally, LNCaP cells grow slower than C4-2B cells, which are castration-resistant cells derived from the hormone-sensitive LNCaP cells.Citation14 This observation indicates that glyphosate and AMPA are more effective in inhibiting growth of rapidly proliferating cancer cells.
Cell growth in a population of cells represents the net outcome of proliferation and apoptosis. Our data indicate that AMPA can arrest cancer cells in the G1/G0 phase of cell cycle, thus inhibiting entry into the S phase. On the other hand, AMPA can enhance apoptosis of cancer cells, as shown by the increased rates of annexin-V-positive cells and increased levels of cleaved PARP, an indicator of apoptosis. Therefore, AMPA inhibits cancer cell growth through inhibition of cellular proliferation and promotion of apoptosis. The molecular mechanism may be that AMPA upregulates the p53 protein level, which subsequently increases p21 protein level. Activation of the p53-p21 pathway is known to cause G1-phase arrest and apoptosis in mammalian cells.Citation20–Citation22 AMPA downregulates the expression of cyclin D3, which may also contribute to the cell cycle arrest.Citation23 AMPA increases procaspase 9 levels and simultaneously decreases procaspase 3 levels, which may mediate apoptosis, as shown in a previous study.Citation24 However, how AMPA initiates these molecular changes and whether these changes apply to other cancer cell lines remain to be determined.
To our best knowledge, this is the first study showing that glycine analogs can inhibit proliferation and promote apoptosis of cancer cells but not normal cells, in vitro at concentrations up to 50 mM. Higher concentrations of the chemicals may affect normal cells, thus producing adverse side effects. These findings suggest that animal studies are warranted to assess the efficacy of glyphosate and AMPA in the treatment of tumors growing in animals and to test whether the effective inhibitory concentrations can be achieved in animal blood. If a positive outcome is obtained in preclinical animal study, it will be feasible to conduct human clinical trials because glyphosate and AMPA are of little toxicity to animals and humans.Citation10 On the other hand, more potent inhibitors may be developed using glyphosate and AMPA as lead compounds. Based on our findings and the previous report,Citation5 it appears promising to develop a new anticancer therapy targeting glycine metabolism.
Acknowledgments
This work was partly supported by a grant from the Department of Defense (PC121647), two grants from the National Institute of General Medical Sciences (P20GM103518) and the National Cancer Institute (R01CA174714) of the National Institutes of Health, the Developmental Fund of Tulane Cancer Center (TCC), and the Louisiana Cancer Research Consortium (LCRC) Fund (to ZY). Dr Zongbing You conceived the idea and designed the experiments. The content of this article is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. The TCC and LCRC FACS Core Facilities were used to conduct this study. Dr Qingli Li received a scholarship from the State Scholarship Fund via China Scholarship Council (CSC) for her training under Dr Zongbing You at Tulane University.
Disclosure
The authors report no conflicts of interest in this work.
References
- Nelson DL Cox MM 2008 Principles of Biochemistry New York, NY WH Freeman and Company 2008
- Kerr SJ Competing methyltransferase systems J Biol Chem 1972 247 13 4248 4252 4338482
- Porter DH Cook RJ Wagner C Enzymatic properties of dimethylglycine dehydrogenase and sarcosine dehydrogenase from rat liver Arch Biochem Biophys 1985 243 2 396 407 2417560
- Hahn RG Dose-dependent half-life of glycine Urol Res 1993 21 4 289 291 8212419
- Jain M Nilsson R Sharma S Metabolite profiling identifies a key role for glycine in rapid cancer cell proliferation Science 2012 336 6084 1040 1044 22628656
- Garrow TA Brenner AA Whitehead VM Cloning of human cDNAs encoding mitochondrial and cytosolic serine hydroxymethyltransferases and chromosomal localization J Biol Chem 1993 268 16 11910 11916 8505317
- Girgis S Nasrallah IM Suh JR Molecular cloning, characterization and alternative splicing of the human cytoplasmic serine hydroxymethyltransferase gene Gene 1998 210 2 315 324 9573390
- Stover PJ Chen LH Suh JR Stover DM Keyomarsi K Shane B Molecular cloning, characterization, and regulation of the human mitochondrial serine hydroxymethyltransferase gene J Biol Chem 1997 272 3 1842 1848 8999870
- Anderson DD Stover PJ SHMT1 and SHMT2 are functionally redundant in nuclear de novo thymidylate biosynthesis PLoS One 2009 4 6 e5839 19513116
- World Health Organization (WHO) Glyphosate and AMPA in Drinking-Water. Background Document for Development of WHO Guidelines for Drinking-Water Quality Geneva WHO 2005
- World Health Organization (WHO) IPCS, Environmental Health Criteria 159: Glyphosate Geneva WHO 1994
- Williams GM Kroes R Munro IC Safety evaluation and risk assessment of the herbicide Roundup and its active ingredient, glyphosate, for humans Regul Toxicol Pharmacol 2000 31 2 Pt 1 117 165 10854122
- You Z Dong Y Kong X Zhang Y Vessella RL Melamed J Differential expression of IL-17RC isoforms in androgen-dependent and androgen-independent prostate cancers Neoplasia 2007 9 6 464 470 17603628
- Thalmann GN Sikes RA Wu TT LNCaP progression model of human prostate cancer: androgen-independence and osseous metastasis Prostate 2000 44 2 91 103 10881018
- Crouch SP Kozlowski R Slater KJ Fletcher J The use of ATP bioluminescence as a measure of cell proliferation and cytotoxicity J Immunol Methods 1993 160 1 81 88 7680699
- Kangas L Grönroos M Nieminen AL Bioluminescence of cellular ATP: a new method for evaluating cytotoxic agents in vitro Med Biol 1984 62 6 338 343 6543460
- Elisia I Popovich DG Hu C Kitts DD Evaluation of viability assays for anthocyanins in cultured cells Phytochem Anal 2008 19 6 479 486 18435529
- Vermes I Haanen C Steffens-Nakken H Reutelingsperger C A novel assay for apoptosis. Flow cytometric detection of phosphatidylserine expression on early apoptotic cells using fluorescein labelled Annexin V J Immunol Methods 1995 184 1 39 51 7622868
- Mead RN Ryu J Liu S Supraphysiologic temperature enhances cytotoxic effects of bupivacaine on bovine articular chondrocytes in an in vitro study Arthroscopy 2012 28 3 397 404 22169763
- He G Siddik ZH Huang Z Induction of p21 by p53 following DNA damage inhibits both Cdk4 and Cdk2 activities Oncogene 2005 24 18 2929 2943 15735718
- el-Deiry WS Harper JW O’Connor PM WAF1/CIP1 is induced in p53-mediated G1 arrest and apoptosis Cancer Res 1994 54 5 1169 1174 8118801
- el-Deiry WS Tokino T Velculescu VE WAF1, a potential mediator of p53 tumor suppression Cell 1993 75 4 817 825 8242752
- Fimognari C Nusse M Berti F Iori R Cantelli-Forti G Hrelia P Cyclin D3 and p53 mediate sulforaphane-induced cell cycle delay and apoptosis in non-transformed human T lymphocytes Cell Mol Life Sci 2002 59 11 2004 2012 12530531
- Sakai T Liu L Teng X Nucling recruits Apaf-1/pro-caspase-9 complex for the induction of stress-induced apoptosis J Biol Chem 2004 279 39 41131 41140 15271982