
Abstract
It is well documented that diabetes mellitus (DM) is strongly associated with cognitive decline and structural damage to the brain. Cognitive deficits appear early in DM and continue to worsen as the disease progresses, possibly due to different underlying mechanisms. Normal iron metabolism is necessary to maintain normal physiological functions of the brain, but iron deposition is one of the causes of some neurodegenerative diseases. Increasing evidence shows that iron overload not only increases the risk of DM, but also contributes to the development of cognitive impairment. The current review highlights the role of iron overload in diabetic cognitive impairment (DCI), including the specific location and regulation mechanism of iron deposition in the diabetic brain, the factors that trigger iron deposition, and the consequences of iron deposition. Finally, we also discuss possible therapies to improve DCI and brain iron deposition.
Introduction
Cognitive impairment is a common complication of diabetes mellitus (DM), manifested in severe deficits in attention, processing speed, learning and memory functions, among others.Citation1–3 Epidemiological studies estimate that the decline in cognitive function in DM patients is as high as 19% over 20 years.Citation4 In addition to high complication rates, poorly controlled DM can exacerbate cognitive impairment and may induce neurodegeneration, including Alzheimer’s diseases and Parkinson’s diseases.Citation5–7 Thus, further understanding of the molecular targets and pathways leading to cognitive impairment in DM is urgently needed, which is particularly important for the improvement of therapeutic strategies.
As the most abundant transition metal in the human body, iron plays an important role in maintaining normal growth and development, and physiological metabolism. Specifically, iron is involved in important functions such as oxygen transport of hemoglobin, muscle oxygenation of myoglobin, protein synthesis of the mitochondrial respiratory chain, and DNA synthesis of ribonucleotide reductase, also participates in several enzymatic reactions.Citation8,Citation9
To note, improperly low or high levels of iron are harmful and lead to a variety of diseases. In fact, previous studies have confirmed a strong association between pathologic iron overload and DM risk,Citation10 which is mainly reflected in iron deposition -induced oxidative stress/lipid peroxidation, insulin resistance, abnormal glucose and lipid metabolism, and chronic inflammation.Citation10–15 Iron overload is also associated with many complications of DM,Citation16–18 including diabetic cognitive impairment (DCI).Citation19
As reported, increased iron levels are observed in neurodegenerative diseases.Citation20,Citation21 Considering that DM promotes and aggravates central nervous system (CNS) injury, iron overload may be an important intermediate target. Therefore, we reviewed and collated existing evidence with the aim of gaining a clearer understanding of the contribution of iron overload to DCI. The key question is: where and in what cells in the brain does the iron overload occur? What triggers iron overload and what are its ultimate consequences? What are the current interventions?
Location of Brain Iron Deposits in DCI
As an important part of CNS, hippocampus has the function of learning and memory formation.Citation22 Diabetic models exhibited marked hippocampal atrophy and impaired synaptogenesis leading to memory deficits.Citation23,Citation24 Recent studies have confirmed that there is obvious iron deposition in the hippocampus of type 1 diabetes mellitus (T1DM) model, through an iron assay kit.Citation25 Similarly, type 2 diabetes mellitus (T2DM) has demonstrated increased iron content throughout the hippocampus, including cornu ammonis (CA) 1 region, CA3 region, and the dentate gyrus (DG) region,Citation19,Citation26 while neurons in the hippocampal DG–CA3-CA1 circuit form neural representations of space and support spatial navigation and episodic memory.Citation27 Noteworthy, increased iron uptake in hippocampal neurons in T2DM triggers cell death, and damage to multiple regions of the hippocampus may be involved.Citation28 Similar to neurons, hippocampal astrocytes in T2DM models have been found to exhibit iron hyperuptake and to be associated with astrocyte polarization.Citation29 This is an important finding, as iron transfer from the periphery into neurons is tightly regulated by astrocytes.Citation30 Additional evidence suggests that iron supplementation increases the activation of microglia in the hippocampus of T2DM rats, which also leads to exacerbation of existing neuroinflammation and exacerbates the risk of neurodegenerative diseases.Citation31 In short, iron overload occurs in both neurons and glia (astrocytes and microglia) as an exacerbating factor in DM-induced hippocampal damage. This extensive iron deposition may be a notable factor triggering memory cognitive dysfunction.
The basal ganglia, including the striatum (putamen, caudate, and pallidum), the substantia nigra, and the subthalamic nucleus, are responsible for executive function, working memory tasks, adaptive motor, and sensorimotor learning.Citation32 Using quantitative sensitivity mapping (QSM), T2DM patients with mild cognitive impairment were found to have greater iron deposition in the right caudate nucleus, substantia nigra, and left putamen, which are strongly associated with cognitive impairment.Citation33 In addition, excessive iron deposition in the putamen of T2DM patients was further confirmed,Citation34 which may affect cortico-striato-thalamocortical neural pathways and impede normal neurotransmission, manifesting as dysfunction of voluntary movement, cognition, language, and sensory.Citation34 Similarly, iron deposition in the striatum and frontal lobe of T2DM patients was elevated as assessed by quantitative imaging QSM and may contribute to the decline in executive function in T2DM.Citation35 Iron in the hypothalamus of T1DM rats is excess and imbalanced, showing increased cytoplasmic iron stores and decreased mitochondrial iron stores. This dysregulation can lead to neuronal degeneration and imbalances in energy homeostasis.Citation36 The above studies confirmed the iron deposition in the basal ganglia and tightly connected areas (cortex and thalamus) induced by DM, and the abnormalities in multiple cognitive functions caused by this should also be taken seriously.
It has been confirmed that in the T2DM mouse model cerebellum contains higher levels of monoamine oxidase (MAO)-A and -B, an enzyme that catalyzes the oxidative deamination of biogenic amines, which promotes the loss of neurotransmitters such as dopamine and 5-hydroxytryptamine. live.Citation37 The multifunctional iron-chelating drug M30 inhibited the enhanced MAO-A activity and decreased the MAO-B activity.Citation37 Although the literature is limited, this may indirectly reflect the importance of iron homeostasis in the cerebellum of diabetic models on dopaminergic metabolism, which deserves further exploration and confirmation.
Mechanisms of Brain Iron Deposition in DCI
Cellular Iron Metabolism and Regulation in the Brain
Brain iron uptake first crosses the blood-brain barrier (BBB)- composed mainly of brain microvascular endothelial cells (BMVEC) and astrocytes.Citation38 Specifically, ferric transferrin (Tf) enters BMVEC by binding to the transferrin receptor (TfR), and forms endosomes. Due to the change of microenvironment pH, Fe dissociates from Tf and reduces from ferric ion to ferrous ion, and then releases the reduced iron into the cytoplasm through divalent metal-ion transporter-1 (DMT1). Finally, it leaves BMVEC through ferroportin (FPN) and is exported into the brain interstitial fluid, accompanied by the oxidation of ferrous ions by ceruloplasmin (mostly synthesized and released by the choroid plexus) to iron ions and loaded onto TfCitation38,Citation39 ().
Figure 1 Iron enters the brain and is transported to neurons and glial cells. Iron-binding Tf binds to TfR and is internalized in endosomes. Acidification of endosomes dissociates Fe from Tf and reduces it to ferrous ions, which are subsequently released into the cytoplasm by DMT1. Finally, ferrous ions leave BMVECs through FPN1 and are exported into the interstitial fluid of the brain, where they are oxidized to ferric by ceruloplasmin. Neurons and microglia mainly take up Fe through the Tf-Tfr1 system, while astrocytes can take up iron directly from BMVECs through DMT1. Created with Biorender.com.
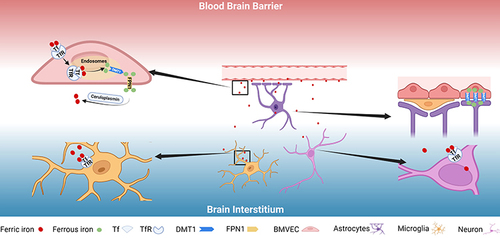
After crossing the BBB, ferric transferrin in the interstitial fluid is taken up by neurons and glial cells through TfR-mediated endocytosis to meet their metabolic needs,Citation40 and neurons absorb ferric iron.Citation41 In addition, DMT1 is strongly expressed in the peduncles of astrocytes that directly contact BMVEC, suggesting the ability to uptake amounts of non-Tf-bound iron directly from the BBB.Citation42–44 Although it is speculated that neuronal DMT1 has the role of non-transferrin binding uptake, this has not been fully confirmed in vivo.Citation45 As the only known iron exporter, FPN1 widely exists in BMVEC, neurons and various glial cells, which dominates the release of iron in nerve cells in the brainCitation46 (). Thus, CNS iron uptake is tightly regulated, and iron homeostasis in any neural cell requires a balance of uptake and export.
Iron Overload Mechanism of DCI
As mentioned before, the iron absorption process begins at the BBB. Although increased iron exposure may lead to iron accumulation in multiple areas, the BBB may be the first area of the brain attacked by excess iron. The study found that the integrity of the BBB permeability in the cortex and striatum of DM rats was damaged after stroke. While the application of the iron chelator effectively protected the BBB, conversely confirming the contribution of iron overload in the destruction of the BBB induced by DM.Citation47 In addition, iron promoted the death of primary BMVEC isolated from DM animals, while iron chelator could effectively increase the viability of BMVEC.Citation47 Recent reports indicate that iron chelator can protect the integrity of glial vessels and improve the survival of endothelial cells in the brain of female DM mice.Citation48 Although the protection of the BBB is unquestionable, the targets and regulatory mechanisms of iron chelator in both sexes seem to be inconsistent, prompting researchers to pay attention to the differences in sensitivity to iron accumulation caused by different sexes and the impact on the BBB.
Cellular iron homeostasis is carefully regulated by multiple proteins, including TfR1 as an iron uptake protein and FPN1 as an export protein.Citation49 The study found that TfR1 was upregulated and FPN1 was downregulated in hippocampus of T2DM model, which may be an important factor causing neuronal iron overload. Similarly, downregulation of the FPN1 gene expression in the hippocampus was also found in T1DM rats.Citation19,Citation25 A recent report showed increased co-expression of DMT1 with reactive astrocytes in the hippocampus of T2DM mice.Citation29 Considering that upregulation of DMT1 expression leads to iron accumulation, which further promotes astrocyte activation,Citation50 this may partly explain the structural and metabolic abnormalities of astrocytes induced by DM.Citation51,Citation52 This seems to be a good understanding of why iron overload occurs. Simply put, more iron enters the cells and less exits.
Changes in ferritin expression should also be of concern. Ferritin is a hollow iron storage protein, consisting of a heavy chain (FTH) and a light chain (FTL), which can store a large amount of free iron entering cells until the cavity is saturated, thus protecting cells from iron toxicity damage.Citation53,Citation54 In general, the composition of ferritin varies greatly in different cells. Most of the ferritin in tissues with iron storage function is FTL, which has a more stable structure and stores more iron; while tissues with high iron oxidation activity mainly contain FTH, which has significant antioxidant activity.Citation55,Citation56 In the brain, neurons mainly express FTH, while glial cells mainly express FTL.Citation57–59 It has been confirmed that FTH expression is significantly decreased in the hippocampus of T2DM model,Citation19 which may represent the presence of a large amount of free iron in cells. This severely affects neuronal survival as free iron (especially ferrous iron) is a key factor in promoting cellular oxidative damage and lipid peroxidation,Citation60 and low expression of FTH is insufficient to counteract them. In fact, iron buffer and ferritin are mainly found in glial cells rather than neurons.Citation61 FTL did not show significant differential expression in T2DM brains,Citation19,Citation29 which may represent a saturated state of iron stores and an inability to accommodate excess iron. Interestingly, the level of ferritin was significantly increased in the hippocampus of T1DM rats, without distinguishing between FTL and TFH.Citation62 This seems to indicate differential regulation of ferritin in different types of DM brains, which requires further investigation of the specific mechanisms.
In addition to the classical mechanisms of iron regulation, it is also important to focus on specific proteins or channels that regulate iron metabolism. The study found upregulation of lipocalin-2 (LCN2) in the hippocampus and lipopolysaccharide/high glucose-treated neurons of DM mice, which may promote DCI.Citation63 Although the researchers did not mention the significance of DM-induced LCN2 elevation on iron overload, it was confirmed that the expression of LCN2 in the brain was positively correlated with iron overload in various brain regions.Citation64 In fact, LCN2 is a transferrin-independent iron transporter that associates with siderophore and participates in cellular iron transport.Citation65 It has also been reported that L-type calcium channels are the main way for cells to take up ferrous iron when iron content increases.Citation66 Whether LCN2 and L-type calcium channels are involved in DM-induced brain iron deposition remains to be further confirmed.
Factors of Diabetic Brain Iron Deposition
As mentioned above, complex regulation of iron-related proteins maintains cellular iron acquisition, utilization and elimination, but they are often intervened differently. It has been confirmed that hepcidin is the main factor regulating iron metabolism.Citation67 As an antimicrobial peptide, hepcidin is mainly secreted in the liver,Citation67 also expressed in the brain.Citation68 It has been reported that hepcidin deficiency in T2DM may result in iron deposition in the hippocampus and consequent cognitive impairment.Citation69 The researchers did not explore the specific consequences of underexpression of hepcidin, but hepcidin has been shown to regulate the expression of iron transporters such as TfR1, DMT1, and FPN1 in BMVEC.Citation70 Interestingly, many studies have demonstrated the downregulation of FPN1 in the DM hippocampus,Citation19,Citation25,Citation29 which seems to contradict the conclusion of low expression of hepcidin, because hepcidin binds to FPN1 and functionally downregulates FPN1 to affect iron availability.Citation71 In addition, the expression rate of hepcidin in the brain is low and a considerable part actually comes from the liver,Citation68 but the expression of hepcidin in DM patients has a completely opposite conclusion.Citation72,Citation73 This may be due to the characteristics of different diabetic patients and the complex regulatory factors of hepcidin, but may the same be true of hepcidin levels in the brain?Citation62 More experimental and clinical studies are needed to confirm or refute the specific role of hepcidin in DM.
In any case, the contribution of hyperglycemia should not be ignored, as it is the most important pathological outcome of DM. Neurons have a continuously high demand for glucose, and neuronal uptake of glucose depends on the concentration of extracellular glucose. Therefore, persistent hyperglycemia induced by DM can lead to a multiplied increase in neuronal glucose levels, and excessive glucose metabolism can lead to neuronal damage, known as glucose neurotoxicity.Citation74 Studies have found that diabetic hyperglycemia aggravates neuronal death by inhibiting the transcription and secretion of lactoferrin in neutrophils and accumulating iron in neurons.Citation75 In addition, earlier reports suggested a positive correlation between serum ferritin and serum glucose levels.Citation76 In the periphery, excess iron interferes with glucose metabolism and causes DM by disrupting insulin signaling,Citation77,Citation78 whereas iron reduction has been shown to improve pathological hyperglycemia.Citation79,Citation80 Thus, a high-glucose environment promotes iron overload and vice versa. However, due to the lack of reports, the specific mechanisms leading to hyperglycemia and iron metabolism disorders still need to be further explored.
As the main pathological factor of T2DM, insulin resistance (IR) should also be mentioned. In fact, compared with T1DM, T2DM has a higher incidence of cognitive impairment, and T2DM induces neurological damage and cognitive impairment earlier than T1DM,Citation81 so more attention should be paid to the independent role of IR. IR triggers neuronal insulin signaling dysregulation, which may cause DCI.Citation82–84 Insulin resistance has been shown to induce redistribution of TfR on the cell surface, promoting iron overload.Citation85,Citation86 In turn, iron overload and elevated ferritin levels inhibit insulin synthesis and secretion and induce IR.Citation87,Citation88 It has been reported that abnormal iron metabolism aggravates memory impairment and neuronal survival in DM, mainly by exacerbating IR in the CNS.Citation89 Also be concerned about the crosstalk between IR and hepcidin: IR may have interfered with hepcidin expression and secretion, whereas treatment with hepcidin gene silencing improved IR.Citation90,Citation91 Anyway, the role of IR on brain iron overload and the specific mechanism deserve further investigation.
Attention should be paid to the promoting effect of inflammation on brain iron deposition in DM, as DM has been proved to be systemic chronic inflammation.Citation92 A number of inflammatory factors were confirmed to be significantly increased in DM model brains, such as interleukin (IL)-1, IL-6 and tumor necrosis factor (TNF)-α.Citation93–95 Note that these cytokines can enhance the strength of iron transport. It has been demonstrated that inflammation (especially the secretion of TNF-α and IL-6) increases iron accumulation in neurons due to the resulting upregulation of DMT1.Citation96 More studies have confirmed the consistency of DMT1 and TNF-α overexpression in CNS inflammation models.Citation50,Citation97 Another interesting conclusion was the significant downregulation of FPN protein levels in neurons following inflammatory stimulation of TNF-α and IL-6, regardless of the effect of hepcidin.Citation96 In addition, increased DMT1 and TfR1 levels and decreased FPN1 were also observed in IL-1β or TNF-α-treated neurons.Citation98 In T1DM, iron deposition in the brain is mainly caused by inflammation-induced BBB and damage,Citation99 and neuronal iron deposition is also strongly associated with inflammation.Citation62 In fact, inflammation and iron overload are like a chicken-and-egg relationship. In other words, neuroinflammation in DM does not necessarily occur earlier than brain iron overload, because it has been reported that deferoxamine can target neuroinflammation to alleviate DCI.Citation100
Consequences of Brain Iron Deposition in DCI
As a factor that triggers or exacerbates DCI, brain iron accumulation has been confirmed to occur in multiple regions of the brain and act on a variety of cell damage. Considering the brain’s high iron requirement and sensitivity to reactive oxygen species (ROS), brain iron accumulation and concomitant oxidative damage were confirmed to be associated with early DCI.Citation101,Citation102 The biological function of iron is based on its redox ability, that is, the interconversion between the ferrous and ferric states.Citation103 At the same time, excess iron can be toxic due to iron’s ability to exist in various oxidation states. Specifically, when an excess of iron is present, the formation of highly reactive hydroxyl radicals, an excess of hydrogen peroxide, and a deficiency of antioxidants can be catalyzed by the Fenton reactionCitation104 (). A large number of studies have confirmed the increase of ROS in the brain of DM models, which is often accompanied by the weakening of antioxidant systems, such as superoxide dismutase (SOD), glutathione/glutathione disulfide (GSH/GSSG) and catalase (CAT).Citation105–108 It also showed elevation of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) 2 and NOX4, the main endogenous biological systems for ROS formation.Citation29 It has been demonstrated that direct glucotoxicity in neurons is mainly due to increased intracellular glucose oxidation, leading to enhanced ROS production.Citation109 High glucose can also consume NADPH through the sorbitol pathway to disrupt the GSH cycle, reduce the ability of glutathione peroxidase to metabolize H2O2 into water, and then increase the channel for H2O2 to enter the Fenton reaction, thereby generating super hydroxyl radicals.Citation74 This oxidative stress may be the initiator of multiple pathological effects and interact through different pathways, eventually inducing the progression of brain damage in DM.Citation110
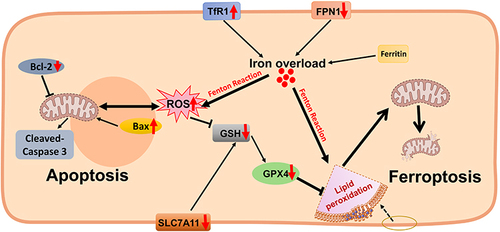
Figure 2 Excess iron-induced cell death in the diabetic brain. The expression of TfR1 and DMT1 is up-regulated in the diabetic brain (especially in hippocampal neurons), and the expression of FPN1 is down-regulated, which may lead to iron overload. As a direct consequence, iron overload can increase ROS and lipid oxide levels and reduce antioxidant activity such as GSH through the Fenton reaction. On the one hand, this leads to the damage of mitochondrial structure and function, the down-regulation of GPX4 and SLC7A11 protein expression and triggers neuronal ferroptosis; on the other hand, it may lead to the up-regulation of Bax protein and the down-regulation of Bcl-2 protein, triggering apoptosis. Some of the materials in are created with Biorender.com.
Among the many targets of oxidative stress, lipids are the most implicated class of biomarkers. A large number of oxidants attack cell membranes, lipoproteins, and other lipid-containing structures, and lipid peroxidation occurs. Since the brain is rich in polyunsaturated fatty acids and relatively deficient in the oxidation system, lipid peroxidation easily occurs when the redox imbalance in the brain occurs.Citation111 As mentioned earlier, high levels of lipid peroxidation are an important cause of ferroptosis, so it is necessary to pay attention to changes in lipid peroxidation. Malondialdehyde is a common lipid peroxidation product, while 4-hydroxynonenal, the most biologically active lipid peroxidation product, has various cytotoxic effects of lipid peroxidation, such as depletion of GSH, dysfunction of structural proteins, reduction of enzyme activity, and induction of cell death,Citation112 which have been shown to affect the severity of diabetic complications.Citation19,Citation29,Citation105–108 Note that iron is also an important component of lipid peroxidase subunits. High glucose and palmitic acid treatment increased Fe2+ in mouse hippocampal primary neurons, which in turn increased ROS and lipid peroxidation levels and decreased GSH concentration and GSH/GSSG ratio.Citation26 Similarly, under high glucose concentration environment, iron overload can aggravate the oxidative stress injury of PC12 neurons, possibly through the NF-E2-related factor 2 (Nrf2) /ARE signaling pathway.Citation113 Interestingly, administration of deferoxamine did effectively reverse excessive protein carbonylation in the prefrontal cortex and hippocampus of the DM model, but had no effect on the activity of antioxidant enzymes.Citation108 This is understandable since oxidative stress does not occur solely due to iron overload, as confirmed by other reports.Citation114,Citation115
Ferroptosis has been shown to be involved in the development of various diabetic complications,Citation116,Citation117 also with neurodegeneration.Citation89 Ferroptosis, first proposed in 2012, is a regulated non-apoptotic form of cell death mainly due to inactivation of glutathione peroxidase 4 (GPX4) and iron-dependent lipid peroxidation.Citation118,Citation119 Excess iron accumulates in cells and generates a large number of ROS, which provide a powerful oxidant for lipid peroxidation and lead to the biogenesis of polyunsaturated fatty acids and the occurrence of ferroptosis.Citation120,Citation121 Iron overload is a key factor of ferroptosis, and the use of iron chelators can effectively reduce the occurrence of ferroptosis in various experimental models.Citation118,Citation122 The presence of ferroptosis in DCI has been widely documented. Downregulation of GPX4 and solute carrier family 7 member 11 (SLC7A11) in the hippocampus of T2DM models triggered ferroptosis, manifested by abnormalities in mitochondrial membranes and collapse of mitochondrial cristae,Citation19,Citation26,Citation28,Citation123 especially in neuronsCitation26,Citation28 (). Similarly, T1DM has also been focused on the presence of ferroptosis in the hippocampus, including damage to neuronal mitochondria.Citation25 It has also been reported that epidermal growth factor/Nrf2/heme oxygenase-1 signaling and the microbiota-gut-brain axis regulate ferroptosis in T2DM hippocampal neurons.Citation124 Ferroptosis of BMVECs in T2DM should also be taken seriously, as this may exacerbate BBB disruption and cognitive impairment after diabetic stroke,Citation47 possibly related to the p53/GPX4 axis.Citation125 Overall, iron overload plays an important role as well as ferroptosis, especially in the context of DM-induced cognitive decline,Citation89 and onset of iron overload may be the main driver of ferroptosis.
Apoptosis, also known as programmed cell death, occurs during normal physiological processes (development and aging) and acts as a homeostatic mechanism to maintain cell populations.Citation126 Inappropriate apoptosis can induce a variety of diseases, including neurodegenerative diseases, ischemic injury, autoimmune diseases and many types of cancer. A number of evidences have shown that hyperglycemia and its metabolic disorders can cause necrosis and apoptosis of nerve cells, and there is a significant correlation with DCI.Citation127–129 Iron damages membrane lipids, proteins, and nucleic acids through ROS toxicity, thereby increasing neuronal apoptosis and affecting its function.Citation130 A new pathway for the induction of apoptosis under glucolipotoxic conditions has been identified, involving increased iron import, triggered by depletion of mitochondrial membrane potential and generation of hydroxyl radicals from hydrogen peroxide by the Fenton reaction.Citation131 Susceptibility to apoptosis has been shown to be controlled by the proapoptotic B-cell lymphoma-2 (BCL2) homologue family. Briefly, upregulation of pro-apoptotic Bcl-2-associated X protein (Bax) and downregulation of the homologous anti-apoptotic Bcl-2 protein, as well as activated cleavage of Caspase-3, are important mechanisms of apoptosis.Citation126 Up-regulated Bax and down-regulated Bcl-2 in the hippocampus of iron-overloaded rats confirmed the role of iron accumulation in promoting neuronal apoptosisCitation132 (). Although no direct evidence was found to confirm the role of iron overload in DM brain on apoptosis, follow-up studies should be considered.
Treatment Strategies for DCI and Brain Iron Deposition
It is well known that good glycemic control significantly reduces the risk of DM complications and improves diabetic complications. However, studies have found that simple control of blood glucose does not completely prevent the occurrence and progression of complications, especially CNS lesions.Citation133,Citation134 In other words, blood glucose levels had a limited effect on DCI.Citation133,Citation134 Therefore, it makes sense to test antidiabetic drugs to treat cognitive impairment or to focus on other drugs that improve DCI.
Liraglutide, a glucagon-like peptide-1 receptor agonist, is clinically approved for obesity and T2DM.Citation135 The protective effects of liraglutide against neurodegeneration and cerebral ischemic events have been extensively demonstrated,Citation136 also involving DCI,Citation137,Citation138 beyond its hypoglycemic effect. Our previous studies demonstrated the broad effects of liraglutide on hippocampal iron overload in T2DM models, including neurons and astrocytes, which in turn improved DCI.Citation19,Citation29 In fact, liraglutide also had a significant effect on iron deposition in the DM liver, which is the first major metabolic organ of iron.Citation139 These findings seem to encourage the use of liraglutide against DCI and brain iron overload, but further confirmation by clinical trials is needed.
As mentioned earlier, numerous studies have confirmed the contribution of iron overload to DCI, which makes it possible to intervene by means of iron chelation. Studies have confirmed the favorable effects of deferoxamine in alleviating DCI by inhibiting oxidative stress, neuroinflammation and regulating iron homeostasis.Citation100 In addition, the application of deferoxamine can regulate the immune content of c-Jun N-terminal kinase, mitogen-activated protein kinase-38, brain-derived neurotrophic factor, and protein kinases A and C in the prefrontal cortex, hippocampus, amygdala, and nucleus accumbens of DM rats, confirming its regulatory effect on intracellular pathways in brain regions.Citation140 In a post-stroke memory impairment model in DM, deferoxamine prevented DM-mediated glial vascular remodeling and compromised BBB integrity, while improving diabetic memory function, suggesting a new therapeutic strategy for iron chelation to maintain glial vessel integrity and improve endothelial cell survival.Citation48 Similarly, deferoxamine treatment prevented vascular regression and microglial activation in DM rats and improved aquaporin-4 polarity and BBB permeability, while reducing iron overload-induced BMVEC ferroptosis and lipid peroxidation, and improving post-stroke cognitive impairment in DM.Citation47 Although iron chelation has been recognized as a means of improving various diabetic complications,Citation141,Citation142 it should be used with caution in clinical practice, because iron deficiency caused by inappropriate iron chelation is also harmful.
Traditional Chinese medicine (TCM) seems to have received some attention and recognition in the prevention and treatment of DCI. TCM is characterized by multi-component, multi-target interactions with few adverse reactions.Citation143 In addition to compounds, the main active ingredient of a certain drug may also prove to have a protective effect against DCI. Danggui-Shaoyao-San has been shown to balance oxidant/antioxidant levels, improve the expression of neurotrophic factors and reduce neuronal apoptosis, thereby alleviating DM-induced cognitive dysfunction.Citation144 Furthermore, in addition to anti-neuron injury, Danggui-Shaoyao-San also has a good effect on physiological regulation such as iron metabolism.Citation145 Berberine, the main component of Coptis chinensis Franch., has been shown to play a protective role against DCI by reducing hippocampal neuron damage, improving tau protein hyperphosphorylation and repairing axonal damage.Citation146,Citation147 Interestingly, berberine treatment can reduce cellular iron overload through proteins that regulate iron uptake.Citation148 Although TCMs may have multiple effects including improving DCI and regulating iron metabolism, valid pharmacological and toxicological evaluations are required.
Lifestyle changes and the need for ongoing self-regulation are major components of DM care. Studies have found that DM and its complications can be largely prevented and alleviated through interventions that promote lifestyle changes, such as a healthy diet and regular physical exercise.Citation149 Although lifestyle interventions do not appear to significantly change the prevalence of DCI,Citation150,Citation151 the beneficial effects of various lifestyle interventions on neurodegeneration have been demonstrated,Citation152 so more specific forms of lifestyle intervention may Targeted and beneficial changes to DCI.Citation153 Note that in animal models, low-iron diets significantly improved DM (mainly showing significant increases in insulin sensitivity and beta-cell function).Citation78 Could this benefit apply to DCI? More clinical and experimental evidence is still needed.
Conclusions
Due to the link between iron overload and DCI, the “iron” factor of brain injury in DM has been emphasized. Here, we review the distribution, mechanisms, causes, and consequences of iron overload in the DM brain, and try to discover drugs that can effectively intervene. In fact, the specific mechanism of the relationship between iron metabolism and DCI is not fully understood, and the pathological consequences of iron overload also need to be further explored. Based on the above reports, we believe that controlling iron metabolism in the central nervous system may be a promising therapeutic approach for DCI, but the vast majority of reports still use basic research, and these still need to be translated into evidence for clinical trials.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no conflicts of interest in this work.
Additional information
Funding
References
- Cukierman T, Gerstein HC, Williamson JD. Cognitive decline and dementia in diabetes--systematic overview of prospective observational studies. Diabetologia. 2005;48(12):2460–2469. doi:10.1007/s00125-005-0023-4
- Koekkoek PS, Kappelle LJ, van den Berg E, Rutten GE, Biessels GJ. Cognitive function in patients with diabetes mellitus: guidance for daily care. Lancet Neurol. 2015;14(3):329–340. doi:10.1016/S1474-4422(14)70249-2
- McCrimmon RJ, Ryan CM, Frier BM. Diabetes and cognitive dysfunction. Lancet. 2012;379(9833):2291–2299. doi:10.1016/S0140-6736(12)60360-2
- Rawlings AM, Sharrett AR, Schneider AL, et al. Diabetes in midlife and cognitive change over 20 years: a cohort study. Ann Intern Med. 2014;161(11):785–793. doi:10.7326/M14-0737
- Schernhammer E, Hansen J, Rugbjerg K, Wermuth L, Ritz B. Diabetes and the risk of developing Parkinson’s disease in Denmark. Diabetes Care. 2011;34(5):1102–1108. doi:10.2337/dc10-1333
- Ashraghi MR, Pagano G, Polychronis S, Niccolini F, Politis M. Parkinson’s disease, diabetes and cognitive impairment. Recent Pat Endocr Metab Immune Drug Discov. 2016;10(1):11–21. doi:10.2174/1872214810999160628105549
- Barnes DE, Yaffe K. The projected effect of risk factor reduction on Alzheimer’s disease prevalence. Lancet Neurol. 2011;10(9):819–828. doi:10.1016/S1474-4422(11)70072-2
- Sheftel AD, Mason AB, Ponka P. The long history of iron in the Universe and in health and disease. Biochim Biophys Acta. 2012;1820(3):161–187. doi:10.1016/j.bbagen.2011.08.002
- Milto IV, Suhodolo IV, Prokopieva VD, Klimenteva TK. Molecular and cellular bases of iron metabolism in humans. Biochemistry. 2016;81(6):549–564. doi:10.1134/S0006297916060018
- Gao H, Yang J, Pan W, Yang M. Iron overload and the risk of diabetes in the general population: results of the Chinese health and nutrition survey cohort study. Diabetes Metab J. 2022;46(2):307–318. doi:10.4093/dmj.2020.0287
- Harrison AV, Lorenzo FR, McClain DA. Iron and the Pathophysiology of Diabetes. Annu Rev Physiol. 2023;85:339–362. doi:10.1146/annurev-physiol-022522-102832
- Swaminathan S, Fonseca VA, Alam MG, Shah SV. The role of iron in diabetes and its complications. Diabetes Care. 2007;30(7):1926–1933. doi:10.2337/dc06-2625
- Rajpathak SN, Crandall JP, Wylie-Rosett J, Kabat GC, Rohan TE, Hu FB. The role of iron in type 2 diabetes in humans. Biochim Biophys Acta. 2009;1790(7):671–681. doi:10.1016/j.bbagen.2008.04.005
- Sampaio AF, Silva M, Dornas WC, et al. Iron toxicity mediated by oxidative stress enhances tissue damage in an animal model of diabetes. Biometals. 2014;27(2):349–361. doi:10.1007/s10534-014-9717-8
- Minamiyama Y, Takemura S, Kodai S, et al. Iron restriction improves type 2 diabetes mellitus in otsuka long-Evans Tokushima fatty rats. Am J Physiol Endocrinol Metab. 2010;298(6):E1140–9. doi:10.1152/ajpendo.00620.2009
- Geng W, Pan L, Shen L, et al. Evaluating renal iron overload in diabetes mellitus by blood oxygen level-dependent magnetic resonance imaging: a longitudinal experimental study. BMC Med Imaging. 2022;22(1):200. doi:10.1186/s12880-022-00939-7
- Wang B, Zhang Y, Sun N, et al. MRI-measured myocardial iron load in patients with severe diabetic heart failure. Clin Radiol. 2018;73(3):324 e1–324 e7. doi:10.1016/j.crad.2017.10.012
- Zhang WL, Meng HZ, Yang MW. Regulation of DMT1 on bone microstructure in type 2 diabetes. Int J Med Sci. 2015;12(5):441–449. doi:10.7150/ijms.11986
- An JR, Su JN, Sun GY, et al. Liraglutide alleviates cognitive deficit in db/db Mice: involvement in oxidative stress, iron overload, and ferroptosis. Neurochem Res. 2022;47(2):279–294. doi:10.1007/s11064-021-03442-7
- Gozzelino R, Arosio P. Iron homeostasis in health and disease. Int J Mol Sci. 2016;17(1):130. doi:10.3390/ijms17010130
- Hofer T, Perry G. Nucleic acid oxidative damage in Alzheimer’s disease-explained by the hepcidin-ferroportin neuronal iron overload hypothesis? J Trace Elem Med Biol. 2016;38:1–9. doi:10.1016/j.jtemb.2016.06.005
- Knierim JJ. The hippocampus. Curr Biol. 2015;25(23):R1116–21. doi:10.1016/j.cub.2015.10.049
- Milne NT, Bucks RS, Davis WA, et al. Hippocampal atrophy, asymmetry, and cognition in type 2 diabetes mellitus. Brain Behav. 2018;8(1):e00741. doi:10.1002/brb3.741
- Grillo CA, Piroli GG, Wood GE, Reznikov LR, McEwen BS, Reagan LP. Immunocytochemical analysis of synaptic proteins provides new insights into diabetes-mediated plasticity in the rat hippocampus. Neuroscience. 2005;136(2):477–486. doi:10.1016/j.neuroscience.2005.08.019
- Hao L, Mi J, Song L, et al. SLC40A1 mediates ferroptosis and cognitive dysfunction in type 1 diabetes. Neuroscience. 2021;463:216–226. doi:10.1016/j.neuroscience.2021.03.009
- Tang W, Li Y, He S, et al. Caveolin-1 alleviates diabetes-associated cognitive dysfunction through modulating neuronal ferroptosis-mediated mitochondrial homeostasis. Antioxid Redox Signal. 2022;37(13–15):867–886. doi:10.1089/ars.2021.0233
- Soltesz I, Losonczy A. CA1 pyramidal cell diversity enabling parallel information processing in the hippocampus. Nat Neurosci. 2018;21(4):484–493. doi:10.1038/s41593-018-0118-0
- Xie Z, Wang X, Luo X, et al. Activated AMPK mitigates diabetes-related cognitive dysfunction by inhibiting hippocampal ferroptosis. Biochem Pharmacol. 2023;207:115374. doi:10.1016/j.bcp.2022.115374
- An JR, Liu JT, Gao XM, et al. Effects of liraglutide on astrocyte polarization and neuroinflammation in db/db mice: focus on iron overload and oxidative stress. Front Cell Neurosci. 2023;17:1136070. doi:10.3389/fncel.2023.1136070
- Dringen R, Bishop GM, Koeppe M, Dang TN, Robinson SR. The pivotal role of astrocytes in the metabolism of iron in the brain. Neurochem Res. 2007;32(11):1884–1890. doi:10.1007/s11064-007-9375-0
- Gholamhosseinian A, Abbasalipourkabir R, Ziamajidi N, Sayadi M, Sayadi K. The anti-inflammatory effect of omega-3 polyunsaturated fatty acids dramatically decreases by iron in the hippocampus of diabetic rats. Life Sci. 2020;245:117393. doi:10.1016/j.lfs.2020.117393
- Hedden T, Gabrieli JD. Shared and selective neural correlates of inhibition, facilitation, and shifting processes during executive control. Neuroimage. 2010;51(1):421–431. doi:10.1016/j.neuroimage.2010.01.089
- Yang Q, Zhou L, Liu C, et al. Brain iron deposition in type 2 diabetes mellitus with and without mild cognitive impairment-an in vivo susceptibility mapping study. Brain Imaging Behav. 2018;12(5):1479–1487. doi:10.1007/s11682-017-9815-7
- Li J, Zhang Q, Zhang N, Guo L. Increased brain iron deposition in the putamen in patients with type 2 diabetes mellitus detected by quantitative susceptibility mapping. J Diabetes Res. 2020;2020:7242530. doi:10.1155/2020/7242530
- Li J, Zhang Q, Zhang N, Guo L. Increased brain iron detection by voxel-based quantitative susceptibility mapping in type 2 diabetes mellitus patients with an executive function decline. Front Neurosci. 2020;14:606182. doi:10.3389/fnins.2020.606182
- Pandur E, Szabo I, Hormay E, et al. Alterations of the expression levels of glucose, inflammation, and iron metabolism related miRNAs and their target genes in the hypothalamus of STZ-induced rat diabetes model. Diabetol Metab Syndr. 2022;14(1):147. doi:10.1186/s13098-022-00919-5
- Mechlovich D, Amit T, Bar-Am O, Weinreb O, Youdim MB. Molecular targets of the multifunctional iron-chelating drug, M30, in the brains of mouse models of type 2 diabetes mellitus. Br J Pharmacol. 2014;171(24):5636–5649. doi:10.1111/bph.12862
- McCarthy RC, Kosman DJ. Mechanisms and regulation of iron trafficking across the capillary endothelial cells of the blood-brain barrier. Front Mol Neurosci. 2015;8:31. doi:10.3389/fnmol.2015.00031
- McCarthy RC, Kosman DJ. Iron transport across the blood-brain barrier: development, neurovascular regulation and cerebral amyloid angiopathy. Cell Mol Life Sci. 2015;72(4):709–727. doi:10.1007/s00018-014-1771-4
- Dusek P, Hofer T, Alexander J, Roos PM, Aaseth JO. cerebral iron deposition in neurodegeneration. Biomolecules. 2022;12(5):714. doi:10.3390/biom12050714
- Brock JH. Lactoferrin--50 years on. Biochem Cell Biol. 2012;90(3):245–251. doi:10.1139/o2012-018
- Burdo JR, Menzies SL, Simpson IA, et al. Distribution of divalent metal transporter 1 and metal transport protein 1 in the normal and Belgrade rat. J Neurosci Res. 2001;66(6):1198–1207. doi:10.1002/jnr.1256
- Cheli VT, Correale J, Paez PM, Pasquini JM. Iron metabolism in oligodendrocytes and astrocytes, implications for myelination and remyelination. ASN Neuro. 2020;12:1759091420962681. doi:10.1177/1759091420962681
- Wang XS, Ong WY, Connor JR. A light and electron microscopic study of the iron transporter protein DMT-1 in the monkey cerebral neocortex and hippocampus. J Neurocytol. 2001;30(4):353–360. doi:10.1023/a:1014464514793
- Skjorringe T, Burkhart A, Johnsen KB, Moos T. Divalent metal transporter 1 (DMT1) in the brain: implications for a role in iron transport at the blood-brain barrier, and neuronal and glial pathology. Front Mol Neurosci. 2015;8:19. doi:10.3389/fnmol.2015.00019
- Batista-Nascimento L, Pimentel C, Menezes RA, Rodrigues-Pousada C. Iron and neurodegeneration: from cellular homeostasis to disease. Oxid Med Cell Longev. 2012;2012:128647. doi:10.1155/2012/128647
- Abdul Y, Li W, Ward R, et al. Deferoxamine treatment prevents post-stroke vasoregression and neurovascular unit remodeling leading to improved functional outcomes in type 2 male diabetic rats: role of endothelial ferroptosis. Transl Stroke Res. 2021;12(4):615–630. doi:10.1007/s12975-020-00844-7
- Li W, Abdul Y, Chandran R, et al. Deferoxamine prevents poststroke memory impairment in female diabetic rats: potential links to hemorrhagic transformation and ferroptosis. Am J Physiol Heart Circ Physiol. 2023;324(2):H212–H225. doi:10.1152/ajpheart.00490.2022
- Rouault TA. Iron metabolism in the CNS: implications for neurodegenerative diseases. Nat Rev Neurosci. 2013;14(8):551–564. doi:10.1038/nrn3453
- Pelizzoni I, Zacchetti D, Campanella A, Grohovaz F, Codazzi F. Iron uptake in quiescent and inflammation-activated astrocytes: a potentially neuroprotective control of iron burden. Biochim Biophys Acta. 2013;1832(8):1326–1333. doi:10.1016/j.bbadis.2013.04.007
- Moghaddam HK, Baluchnejadmojarad T, Roghani M, et al. Berberine ameliorate oxidative stress and astrogliosis in the hippocampus of STZ-induced diabetic rats. Mol Neurobiol. 2014;49(2):820–826. doi:10.1007/s12035-013-8559-7
- Shen Z, Li ZY, Yu MT, Tan KL, Chen S. Metabolic perspective of astrocyte dysfunction in Alzheimer’s disease and type 2 diabetes brains. Bio Pharmacoth. 2023;158:114206. doi:10.1016/j.biopha.2022.114206
- Lal A. Iron in health and disease: an update. Indian J Pediatr. 2020;87(1):58–65. doi:10.1007/s12098-019-03054-8
- Zhang N, Yu X, Xie J, Xu H. New insights into the role of ferritin in iron homeostasis and neurodegenerative diseases. Mol Neurobiol. 2021;58(6):2812–2823. doi:10.1007/s12035-020-02277-7
- Worwood M. Ferritin in human tissues and serum. Clin Haematol. 1982;11(2):275–307. doi:10.1016/S0308-2261(21)00338-6
- Powell LW, Alpert E, Isselbacher KJ, Drysdale JW. Human isoferritins: organ specific iron and apoferritin distribution. Br J Haematol. 1975;30(1):47–55. doi:10.1111/j.1365-2141.1975.tb00516.x
- Connor JR, Menzies SL. Cellular management of iron in the brain. J Neurol Sci. 1995;134:33–44. doi:10.1016/0022-510x(95)00206-h
- Qi Y, Dawson G. Hypoxia specifically and reversibly induces the synthesis of ferritin in oligodendrocytes and human oligodendrogliomas. J Neurochem. 1994;63(4):1485–1490. doi:10.1046/j.1471-4159.1994.63041485.x
- Connor JR. Iron acquisition and expression of iron regulatory proteins in the developing brain: manipulation by ethanol exposure, iron deprivation and cellular dysfunction. Dev Neurosci. 1994;16(5–6):233–247. doi:10.1159/000112115
- Guo G, Sun M, Li Y, et al. Serum ferritin has limited prognostic value on mortality risk in patients with decompensated cirrhosis: a propensity score matching analysis. Lab Med. 2023;54(1):47–55. doi:10.1093/labmed/lmac064
- Zecca L, Youdim MB, Riederer P, Connor JR, Crichton RR. Iron, brain ageing and neurodegenerative disorders. Nat Rev Neurosci. 2004;5(11):863–873. doi:10.1038/nrn1537
- Zeinivand M, Nahavandi A, Baluchnejadmojarad T, Roghani M, Golab F. Dalteparin as a novel therapeutic agent to prevent diabetic encephalopathy by targeting oxidative stress and inflammation. Basic Clin Neurosci. 2020;11(6):795–804. doi:10.32598/bcn.11.6.1775.1
- Jeong EA, Lee J, Shin HJ, et al. Tonicity-responsive enhancer-binding protein promotes diabetic neuroinflammation and cognitive impairment via upregulation of lipocalin-2. J Neuroinflammation. 2021;18(1):278. doi:10.1186/s12974-021-02331-8
- Kim BW, Jeong KH, Kim JH, et al. Pathogenic upregulation of glial lipocalin-2 in the parkinsonian dopaminergic system. J Neurosci. 2016;36(20):5608–5622. doi:10.1523/JNEUROSCI.4261-15.2016
- Xiao X, Yeoh BS, Vijay-Kumar M. Lipocalin 2: an emerging player in iron homeostasis and inflammation. Annu Rev Nutr. 2017;37:103–130. doi:10.1146/annurev-nutr-071816-064559
- Ma Z, Zhou Y, Xie J. Nifedipine prevents iron accumulation and reverses iron-overload-induced dopamine neuron degeneration in the substantia nigra of rats. Neurotox Res. 2012;22(4):274–279. doi:10.1007/s12640-012-9309-8
- Park CH, Valore EV, Waring AJ, Ganz T. Hepcidin, a urinary antimicrobial peptide synthesized in the liver. J Biol Chem. 2001;276(11):7806–7810. doi:10.1074/jbc.M008922200
- Raha-Chowdhury R, Raha AA, Forostyak S, Zhao JW, Stott SR, Bomford A. Expression and cellular localization of hepcidin mRNA and protein in normal rat brain. BMC Neurosci. 2015;16:24. doi:10.1186/s12868-015-0161-7
- Liu J, Hu X, Xue Y, et al. Targeting hepcidin improves cognitive impairment and reduces iron deposition in a diabetic rat model. Am J Transl Res. 2020;12(8):4830–4839.
- Du F, Qian ZM, Luo Q, Yung WH, Ke Y. Hepcidin suppresses brain iron accumulation by downregulating iron transport proteins in iron-overloaded rats. Mol Neurobiol. 2015;52(1):101–114. doi:10.1007/s12035-014-8847-x
- Nemeth E, Tuttle MS, Powelson J, et al. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science. 2004;306(5704):2090–2093. doi:10.1126/science.1104742
- Jiang F, Sun ZZ, Tang YT, Xu C, Jiao XY. Hepcidin expression and iron parameters change in Type 2 diabetic patients. Diabetes Res Clin Pract. 2011;93(1):43–48. doi:10.1016/j.diabres.2011.03.028
- Sam AH, Busbridge M, Amin A, et al. Hepcidin levels in diabetes mellitus and polycystic ovary syndrome. Diabet Med. 2013;30(12):1495–1499. doi:10.1111/dme.12262
- Tomlinson DR, Gardiner NJ. Glucose neurotoxicity. Nat Rev Neurosci. 2008;9(1):36–45. doi:10.1038/nrn2294
- Xiao Z, Shen D, Lan T, et al. Reduction of lactoferrin aggravates neuronal ferroptosis after intracerebral hemorrhagic stroke in hyperglycemic mice. Redox Biol. 2022;50:102256. doi:10.1016/j.redox.2022.102256
- Fernandez-Real JM, Ricart-Engel W, Arroyo E, et al. Serum ferritin as a component of the insulin resistance syndrome. Diabetes Care. 1998;21(1):62–68. doi:10.2337/diacare.21.1.62
- Ferrannini E. Insulin resistance, iron, and the liver. Lancet. 2000;355(9222):2181–2182. doi:10.1016/S0140-6736(00)02397-7
- Cooksey RC, Jones D, Gabrielsen S, et al. Dietary iron restriction or iron chelation protects from diabetes and loss of beta-cell function in the obese (ob/ob lep-/-) mouse. Am J Physiol Endocrinol Metab. 2010;298(6):E1236–43. doi:10.1152/ajpendo.00022.2010
- Fernandez-Real JM, Penarroja G, Castro A, Garcia-Bragado F, Hernandez-Aguado I, Ricart W. Blood letting in high-ferritin type 2 diabetes: effects on insulin sensitivity and beta-cell function. Diabetes. 2002;51(4):1000–1004. doi:10.2337/diabetes.51.4.1000
- Houschyar KS, Ludtke R, Dobos GJ, et al. Effects of phlebotomy-induced reduction of body iron stores on metabolic syndrome: results from a randomized clinical trial. BMC Med. 2012;10:54. doi:10.1186/1741-7015-10-54
- Zilliox LA, Chadrasekaran K, Kwan JY, Russell JW. Diabetes and cognitive impairment. Curr Diab Rep. 2016;16(9):87. doi:10.1007/s11892-016-0775-x
- Nuzzo D, Picone P, Baldassano S, et al. Insulin resistance as common molecular denominator linking obesity to alzheimer’s disease. Curr Alzheimer Res. 2015;12(8):723–735. doi:10.2174/1567205012666150710115506
- De Felice FG, Benedict C. A key role of insulin receptors in memory. Diabetes. 2015;64(11):3653–3655. doi:10.2337/dbi15-0011
- Takechi R, Lam V, Brook E, et al. Blood-brain barrier dysfunction precedes cognitive decline and neurodegeneration in diabetic insulin resistant mouse model: an implication for causal link. Front Aging Neurosci. 2017;9:399. doi:10.3389/fnagi.2017.00399
- Chung JY, Kim HS, Song J. Iron metabolism in diabetes-induced Alzheimer’s disease: a focus on insulin resistance in the brain. Biometals. 2018;31(5):705–714. doi:10.1007/s10534-018-0134-2
- Noetzli LJ, Mittelman SD, Watanabe RM, Coates TD, Wood JC. Pancreatic iron and glucose dysregulation in thalassemia major. Am J Hematol. 2012;87(2):155–160. doi:10.1002/ajh.22223
- Chen L, Li Y, Zhang F, Zhang S, Zhou X, Ji L. Association of serum ferritin levels with metabolic syndrome and insulin resistance in a Chinese population. J Diabetes Complications. 2017;31(2):364–368. doi:10.1016/j.jdiacomp.2016.06.018
- Fernandez-Real JM, Manco M. Effects of iron overload on chronic metabolic diseases. Lancet Diabetes Endocrinol. 2014;2(6):513–526. doi:10.1016/S2213-8587(13)70174-8
- Cerasuolo M, Di Meo I, Auriemma MC, et al. Iron and ferroptosis more than a suspect: beyond the most common mechanisms of neurodegeneration for new therapeutic approaches to cognitive decline and dementia. Int J Mol Sci. 2023;24(11):9637. doi:10.3390/ijms24119637
- Zhang X, Zhang L, Tan YM, et al. Hepcidin gene silencing ameliorated inflammation and insulin resistance in adipose tissue of db/db mice via inhibiting METs formation. Mol Immunol. 2021;133:110–121. doi:10.1016/j.molimm.2021.02.015
- Wang H, Li H, Jiang X, Shi W, Shen Z, Li M. Hepcidin is directly regulated by insulin and plays an important role in iron overload in streptozotocin-induced diabetic rats. Diabetes. 2014;63(5):1506–1518. doi:10.2337/db13-1195
- Esser N, Legrand-Poels S, Piette J, Scheen AJ, Paquot N. Inflammation as a link between obesity, metabolic syndrome and type 2 diabetes. Diabetes Res Clin Pract. 2014;105(2):141–150. doi:10.1016/j.diabres.2014.04.006
- Rom S, Zuluaga-Ramirez V, Gajghate S, et al. Hyperglycemia-driven neuroinflammation compromises BBB leading to memory loss in both diabetes mellitus (DM) type 1 and type 2 mouse models. Mol Neurobiol. 2019;56(3):1883–1896. doi:10.1007/s12035-018-1195-5
- Yun JH, Lee DH, Jeong HS, Kim HS, Ye SK, Cho CH. STAT3 activation in microglia exacerbates hippocampal neuronal apoptosis in diabetic brains. J Cell Physiol. 2021;236(10):7058–7070. doi:10.1002/jcp.30373
- Lee HJ, Yang SJ. Supplementation with nicotinamide riboside reduces brain inflammation and improves cognitive function in diabetic mice. Int J Mol Sci. 2019;20:17.
- Urrutia P, Aguirre P, Esparza A, et al. Inflammation alters the expression of DMT1, FPN1 and hepcidin, and it causes iron accumulation in central nervous system cells. J Neurochem. 2013;126(4):541–549. doi:10.1111/jnc.12244
- Rathore KI, Redensek A, David S. Iron homeostasis in astrocytes and microglia is differentially regulated by TNF-alpha and TGF-beta1. Glia. 2012;60(5):738–750. doi:10.1002/glia.22303
- Wang J, Song N, Jiang H, Wang J, Xie J. Pro-inflammatory cytokines modulate iron regulatory protein 1 expression and iron transportation through reactive oxygen/nitrogen species production in ventral mesencephalic neurons. Biochim Biophys Acta. 2013;1832(5):618–625. doi:10.1016/j.bbadis.2013.01.021
- Vela D. The dual role of hepcidin in brain iron load and inflammation. Front Neurosci. 2018;12:740. doi:10.3389/fnins.2018.00740
- Zeinivand M, Nahavandi A, Zare M. Deferoxamine regulates neuroinflammation and oxidative stress in rats with diabetes-induced cognitive dysfunction. Inflammopharmacology. 2020;28(2):575–583. doi:10.1007/s10787-019-00665-7
- Altamura S, Muckenthaler MU. Iron toxicity in diseases of aging: alzheimer’s disease, Parkinson’s disease and atherosclerosis. J Alzheimers Dis. 2009;16(4):879–895. doi:10.3233/JAD-2009-1010
- Schroder N, Figueiredo LS, De lima MN. Role of brain iron accumulation in cognitive dysfunction: evidence from animal models and human studies. J Alzheimers Dis. 2013;34(4):797–812. doi:10.3233/JAD-121996
- Papanikolaou G, Pantopoulos K. Iron metabolism and toxicity. Toxicol Appl Pharmacol. 2005;202(2):199–211. doi:10.1016/j.taap.2004.06.021
- Stoyanovsky DA, Tyurina YY, Shrivastava I, et al. Iron catalysis of lipid peroxidation in ferroptosis: regulated enzymatic or random free radical reaction? Free Radic Biol Med. 2019;133:153–161. doi:10.1016/j.freeradbiomed.2018.09.008
- Hoyos CM, Stephen C, Turner A, Ireland C, Naismith SL, Duffy SL. Brain oxidative stress and cognitive function in older adults with diabetes and pre-diabetes who are at risk for dementia. Diabetes Res Clin Pract. 2022;184:109178. doi:10.1016/j.diabres.2021.109178
- Clark GJ, Pandya K, Lau-Cam CA. The effect of metformin and taurine, alone and in combination, on the oxidative stress caused by diabetes in the rat brain. Adv Exp Med Biol. 2017;975(1):353–369. doi:10.1007/978-94-024-1079-2_31
- Marefati N, Abdi T, Beheshti F, Vafaee F, Mahmoudabady M, Hosseini M. Zingiber officinale (Ginger) hydroalcoholic extract improved avoidance memory in rat model of streptozotocin-induced diabetes by regulating brain oxidative stress. Horm Mol Biol Clin Investig. 2021;43(1):15–26. doi:10.1515/hmbci-2021-0033
- Reus GZ, Dos Santos MA, Abelaira HM, et al. Antioxidant treatment ameliorates experimental diabetes-induced depressive-like behaviour and reduces oxidative stress in brain and pancreas. Diabetes Metab Res Rev. 2016;32(3):278–288. doi:10.1002/dmrr.2732
- Bonnefont-Rousselot D. Glucose and reactive oxygen species. Curr Opin Clin Nutr Metab Care. 2002;5(5):561–568. doi:10.1097/00075197-200209000-00016
- Butterfield DA, Halliwell B. Oxidative stress, dysfunctional glucose metabolism and Alzheimer disease. Nat Rev Neurosci. 2019;20(3):148–160. doi:10.1038/s41583-019-0132-6
- Pratico D, Sung S. Lipid peroxidation and oxidative imbalance: early functional events in Alzheimer’s disease. J Alzheimers Dis. 2004;6(2):171–175. doi:10.3233/jad-2004-6209
- Park MW, Cha HW, Kim J, et al. NOX4 promotes ferroptosis of astrocytes by oxidative stress-induced lipid peroxidation via the impairment of mitochondrial metabolism in Alzheimer’s diseases. Redox Biol. 2021;41:101947. doi:10.1016/j.redox.2021.101947
- Zhao S, Zhang L, Xu Z, Chen W. Neurotoxic effects of iron overload under high glucose concentration. Neural Regen Res. 2013;8(36):3423–3433. doi:10.3969/j.issn.1673-5374.2013.36.008
- Zhao M, Li XW, Chen Z, et al. Neuro-protective role of metformin in patients with acute stroke and type 2 diabetes mellitus via AMPK/mammalian target of rapamycin (mTOR) signaling pathway and oxidative stress. Med Sci Monit. 2019;25:2186–2194. doi:10.12659/MSM.911250
- Duzova H, Naziroglu M, Cig B, Gurbuz P, Akatli AN. Noopept attenuates diabetes-mediated neuropathic pain and oxidative hippocampal neurotoxicity via inhibition of TRPV1 channel in rats. Mol Neurobiol. 2021;58(10):5031–5051. doi:10.1007/s12035-021-02478-8
- Yang XD, Yang YY. Ferroptosis as a novel therapeutic target for diabetes and its complications. Front Endocrinol (Lausanne). 2022;13:853822. doi:10.3389/fendo.2022.853822
- Sha W, Hu F, Xi Y, Chu Y, Bu S. Mechanism of ferroptosis and its role in type 2 diabetes mellitus. J Diabetes Res. 2021;2021:9999612. doi:10.1155/2021/9999612
- Dixon SJ, Lemberg KM, Lamprecht MR, et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell. 2012;149(5):1060–1072. doi:10.1016/j.cell.2012.03.042
- Sui X, Zhang R, Liu S, et al. RSL3 drives ferroptosis through GPX4 inactivation and ROS production in colorectal cancer. Front Pharmacol. 2018;9:1371. doi:10.3389/fphar.2018.01371
- Sumneang N, Siri-Angkul N, Kumfu S, Chattipakorn SC, Chattipakorn N. The effects of iron overload on mitochondrial function, mitochondrial dynamics, and ferroptosis in cardiomyocytes. Arch Biochem Biophys. 2020;680:108241. doi:10.1016/j.abb.2019.108241
- Gao M, Monian P, Pan Q, Zhang W, Xiang J, Jiang X. Ferroptosis is an autophagic cell death process. Cell Res. 2016;26(9):1021–1032. doi:10.1038/cr.2016.95
- Lei P, Bai T, Sun Y. Mechanisms of ferroptosis and relations with regulated cell death: a review. Front Physiol. 2019;10:139. doi:10.3389/fphys.2019.00139
- Guo T, Yu Y, Yan W, et al. Erythropoietin ameliorates cognitive dysfunction in mice with type 2 diabetes mellitus via inhibiting iron overload and ferroptosis. Exp Neurol. 2023;365:114414. doi:10.1016/j.expneurol.2023.114414
- Chen J, Guo P, Han M, Chen K, Qin J, Yang F. Cognitive protection of sinomenine in type 2 diabetes mellitus through regulating the EGF/Nrf2/HO-1 signaling, the microbiota-gut-brain axis, and hippocampal neuron ferroptosis. Phytother Res. 2023;37:3323–3341. doi:10.1002/ptr.7807
- Chen C, Huang Y, Xia P, et al. Long noncoding RNA Meg3 mediates ferroptosis induced by oxygen and glucose deprivation combined with hyperglycemia in rat brain microvascular endothelial cells, through modulating the p53/GPX4 axis. Eur J Histochem. 2021;65(3). doi:10.4081/ejh.2021.3224
- Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol. 2007;35(4):495–516. doi:10.1080/01926230701320337
- Hamed SA. Brain injury with diabetes mellitus: evidence, mechanisms and treatment implications. Expert Rev Clin Pharmacol. 2017;10(4):409–428. doi:10.1080/17512433.2017.1293521
- Kong FJ, Ma LL, Guo JJ, Xu LH, Li Y, Qu S. Endoplasmic reticulum stress/autophagy pathway is involved in diabetes-induced neuronal apoptosis and cognitive decline in mice. Clin Sci (Lond). 2018;132(1):111–125. doi:10.1042/CS20171432
- Sadeghi A, Hami J, Razavi S, Esfandiary E, Hejazi Z. The effect of diabetes mellitus on apoptosis in hippocampus: cellular and molecular aspects. Int J Prev Med. 2016;7:57. doi:10.4103/2008-7802.178531
- de Lima MN, Polydoro M, Laranja DC, et al. Recognition memory impairment and brain oxidative stress induced by postnatal iron administration. Eur J Neurosci. 2005;21(9):2521–2528. doi:10.1111/j.1460-9568.2005.04083.x
- Hansen JB, Dos Santos LRB, Liu Y, et al. Glucolipotoxic conditions induce beta-cell iron import, cytosolic ROS formation and apoptosis. J Mol Endocrinol. 2018;61(2):69–77. doi:10.1530/JME-17-0262
- Yun S, He X, Zhang W, Chu D, Feng C. Alleviation effect of grape seed proanthocyanidins on neuronal apoptosis in rats with iron overload. Biol Trace Elem Res. 2020;194(1):210–220. doi:10.1007/s12011-019-01766-8
- Launer LJ, Miller ME, Williamson JD, et al. Effects of intensive glucose lowering on brain structure and function in people with type 2 diabetes (ACCORD MIND): a randomised open-label substudy. Lancet Neurol. 2011;10(11):969–977. doi:10.1016/S1474-4422(11)70188-0
- Erus G, Battapady H, Zhang T, et al. Spatial patterns of structural brain changes in type 2 diabetic patients and their longitudinal progression with intensive control of blood glucose. Diabetes Care. 2015;38(1):97–104. doi:10.2337/dc14-1196
- Tella SH, Rendell MS. Glucagon-like polypeptide agonists in type 2 diabetes mellitus: efficacy and tolerability, a balance. Ther Adv Endocrinol Metab. 2015;6(3):109–134. doi:10.1177/2042018815580257
- Wicinski M, Socha M, Malinowski B, et al. Liraglutide and its neuroprotective properties-focus on possible biochemical mechanisms in alzheimer’s disease and cerebral ischemic events. Int J Mol Sci. 2019;20:5.
- Zhang M, Yan W, Yu Y, et al. Liraglutide ameliorates diabetes-associated cognitive dysfunction via rescuing autophagic flux. J Pharmacol Sci. 2021;147(3):234–244. doi:10.1016/j.jphs.2021.07.004
- Yang Y, Fang H, Xu G, et al. Liraglutide improves cognitive impairment via the AMPK and PI3K/Akt signaling pathways in type 2 diabetic rats. Mol Med Rep. 2018;18(2):2449–2457. doi:10.3892/mmr.2018.9180
- Song JX, An JR, Chen Q, et al. Liraglutide attenuates hepatic iron levels and ferroptosis in db/db mice. Bioengineered. 2022;13(4):8334–8348. doi:10.1080/21655979.2022.2051858
- Reus GZ, Bernardini Dos Santos MA, Abelaira HM, et al. Antioxidant therapy alters brain MAPK-JNK and BDNF signaling path-ways in experimental diabetes mellitus. Curr Neurovasc Res. 2016;13(2):107–114. doi:10.2174/1567202613666160219115832
- Zeinivand M, Sharifi M, Hassanshahi G, Nedaei SE. Deferoxamine has the potential to improve the COVID-19-related inflammatory response in diabetic patients. Int J Pept Res Ther. 2023;29(4):63. doi:10.1007/s10989-023-10516-3
- Hattori N, Schnell O, Bengel FM, et al. Deferoxamine improves coronary vascular responses to sympathetic stimulation in patients with type 1 diabetes mellitus. Eur J Nucl Med Mol Imaging. 2002;29(7):891–898. doi:10.1007/s00259-002-0799-0
- Meng J, Zhu Y, Ma H, Wang X, Zhao Q. The role of traditional Chinese medicine in the treatment of cognitive dysfunction in type 2 diabetes. J Ethnopharmacol. 2021;280:114464. doi:10.1016/j.jep.2021.114464
- Shi JJ, Liu HF, Hu T, et al. Danggui-Shaoyao-San improves cognitive impairment through inhibiting O-GlcNAc-modification of estrogen alpha receptor in female db/db mice. J Ethnopharmacol. 2021;281:114562. doi:10.1016/j.jep.2021.114562
- Fu X, Liu Q, Sun X, Chang H, Liu Y, Han J. Research advances in the treatment of alzheimer’s disease with polysaccharides of danggui-shaoyao-san. J Alzheimers Dis. 2022;85(1):7–19. doi:10.3233/JAD-210656
- Wang S, He B, Hang W, et al. Berberine alleviates tau hyperphosphorylation and axonopathy-associated with diabetic encephalopathy via Restoring PI3K/Akt/GSK3beta Pathway. J Alzheimers Dis. 2018;65(4):1385–1400. doi:10.3233/JAD-180497
- Zhang JH, Zhang JF, Song J, et al. Effects of berberine on diabetes and cognitive impairment in an animal model: the mechanisms of action. Am J Chin Med. 2021;49(6):1399–1415. doi:10.1142/S0192415X21500658
- Wang Y, Yue S, Cai F, et al. Treatment of berberine alleviates diabetic nephropathy by reducing iron overload and inhibiting oxidative stress. Histol Histopathol. 2023:18599. doi:10.14670/HH-18-599
- American Diabetes Association. 9. Professional practice committee: standards of medical care in diabetes-2021. Diabetes Care. 2021;44(Suppl 1):S3. doi:10.2337/dc21-Sppc
- Wing RR, Look ARG. Does lifestyle intervention improve health of adults with overweight/obesity and type 2 diabetes? Findings from the Look AHEAD Randomized Trial. Obesity. 2021;29(8):1246–1258. doi:10.1002/oby.23158
- Espeland MA, Luchsinger JA, Baker LD, et al. Effect of a long-term intensive lifestyle intervention on prevalence of cognitive impairment. Neurology. 2017;88(21):2026–2035. doi:10.1212/WNL.0000000000003955
- Kivipelto M, Mangialasche F, Ngandu T. Lifestyle interventions to prevent cognitive impairment, dementia and Alzheimer disease. Nat Rev Neurol. 2018;14(11):653–666. doi:10.1038/s41582-018-0070-3
- Palta P, Carlson MC, Crum RM, et al. Diabetes and cognitive decline in older adults: the ginkgo evaluation of memory study. J Gerontol a Biol Sci Med Sci. 2017;73(1):123–130. doi:10.1093/gerona/glx076