 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Background
Curcumin, the active ingredient from turmeric rhizomes, has been shown to have a wide range of pharmacological properties including antioxidant and anti-inflammatory effects. Curcumin has been reviewed for its multiple molecular action on inhibiting tumor angiogenesis via its mechanisms of cyclooxygenase (COX)-2, and vascular endothelial growth factor (VEGF) inhibition. In this present study, we aimed to assess the effects of curcumin on preventing diabetes-induced vascular dysfunction in association with COX-2, nuclear factor-κB (NF-κB) expression, and prostanoid production.
Methods
Twelve-week-old male Wistar rats were separated into five groups: 1) diabetes with 0.9% normal saline (DM-NSS; n =10), 2) diabetes treated with curcumin 30 mg/kg (n =10), 3) diabetes treated with curcumin 300 mg/kg (n =10), 4) the control with 0.9% normal saline (n =10), and 5) the control treated with 300 mg/kg (n =10). Daily oral feeding of curcumin was started at 6 weeks after the streptozotocin injection. Levels of 6-keto prostaglandin (PG) F1αand thromboxane (TX) B2 were determined from mesenteric perfusates using enzyme immunoassay kits. Protein kinase C (PKC)-β II and COX-2 with NF-κB levels were analyzed in the mesenteric arteries by immunofluorescent staining and immunohistochemistry, respectively.
Results
The ratio of 6-keto-PGF1αand TXB2 was significantly decreased in DM-NSS compared with the control (P < 0.05). Double-immunofluorescent staining with specific antibodies for PKC-βII and α-smooth muscle actins showed that the diabetic mesenteric arteries contained increased of PKC-βII within the vascular wall. Also, COX-2 expression and activated NF-κB in the small mesenteric artery of diabetes mellitus rats were markedly increased when compared with the control. Interestingly, curcumin could inhibit the upregulation of all of these biomarkers.
Conclusion
These findings show that curcumin can attenuate diabetes-induced vascular dysfunction in association with its potential for COX-2 and NF-κB suppression, PKC inhibition, and improving the ratio of prostanoid products PGI2/TXA2.
Introduction
Diabetes mellitus (DM) is a common metabolic disease with a high and growing prevalence affecting 4% of the population worldwide: 171 million people in the year 2000 and an expected 366 million in 2030.Citation1 Diabetic vascular diseases represent a major cause of mortality and morbidity in diabetic patients. Both micro- and macrovascular complications are the burden of the disease, not only in terms of individual health and wellbeing, but also in terms of the impact on the economic status of a patient’s family and their country.Citation2 It is suggested that hyperglycemia induces an intracellular elevation of reactive oxygen species (ROS). The cumulative ROS can consequently cause long-term changes in the structure and functions of macromolecules, including protein, lipids, and DNA. With this rationale, the dysfunction of the endothelial cells has been documented as a common finding in diabetic patients with its underlining causes of oxidative stress. The potential contribution of increased ROS to the development of endothelial dysfunction in diabetes has received considerable interest, since it interferes with the production of nitric oxide (NO), a key factor in multiple processes of vascular functional homeostasis. It has been pointed out that one of the major pathways that increases ROS production in endothelial cells is the diacylglycerol (DAG)–protein-kinase C (PKC) pathway. Moreover, high-glucose activated PKC upregulation has been reported for its significant role in inducing diabetic endothelial dysfunction.Citation3–Citation6 It has been demonstrated that oxidative stress will be produced more and more via this PKC upregulation, which leads to the activations of nuclear factor-κB (NF-κB) and cyclo-oxygenase (COX)-2 expressions.Citation7–Citation11 Previous studies have shown that in vitro incubation of rabbit arteries with a high glucose concentration increases vasoconstrictor prostanoids. These effects were prevented by both COX inhibitors and prostaglandin (PG)H2/ thromboxane (TX)A2 receptor antagonist, thereby restoring endothelium-dependent relaxation.Citation8 Recent biochemical studies have proposed a possible role for enhanced COX-2 expression in high glucose-induced alterations in vasoconstrictor prostanoid production in cultured endothelial cells.Citation9 Also, it has been demonstrated that upregulation of COX isoforms is associated with a significant elevation of vascular prostaglandin synthesis.Citation10 However, there have been only a limited number of studies investigating the consequences of alterations in microvascular prostanoid synthesis.Citation11
Curcumin, which has been shown to have a wide range of pharmacological properties including antioxidant and anti-inflammatory effects, has been reviewed for its multiple molecular targets on inhibiting PKC, COX-2, and NF-κB expressions.Citation12–Citation14 The antioxidant effect of curcumin has been found to be at least 10 times greater than that of vitamin E. It has been demonstrated that the antioxidant activity of curcumin could be mediated through antioxidant enzymatic systems including superoxide dismutase (SOD), catalase, and glutathione peroxidase.Citation15,Citation16
Even though the beneficial effect of curcumin treatment on hypoglycemia in streptozotocin (STZ)-induced diabetes rats has been reported,Citation17 its molecular mechanisms have not been clarified yet. The mechanism by which curcumin improves this situation is probably due to its hypocholesterolemic influence, antioxidant nature, and increase in plasma insulin levels.Citation18–Citation20
The anti-inflammatory effects of curcumin have been shown to mediate through the suppression of both COX-2 and lipoxygenase proteins as well as the downregulation of NF-κB. The anti-inflammatory pathways of curcumin have been discovered mostly from the tumor cell study, however, the effects of curcumin on a diabetic animal model remain unknown.Citation20–Citation22
Therefore, in the present study, we aimed to assess the effects of plant-derived antioxidant, curcumin, on diabetesinduced endothelial dysfunction in association with its mechanism on NF-κB, COX-2, and prostanoid ratio.
Method
Twelve-week-old male Wistar rats were housed in a temperature- and light-controlled environment, and were fed standard feed and drank tap water ad libitum. The present study was conducted in accordance with the guidelines for animal experimentation established by the National Research Council of Thailand and approved by the Institutional Animal Care and Use Committee of Chulalongkorn University.
Induction of experimental diabetes
The rats were randomly divided into nondiabetic and diabetic groups. Diabetic rats were induced by a single intravenous injection of STZ (55 mg/kg) (Sigma-Aldrich, St. Louis, MO). STZ was freshly prepared by dissolving in citrate buffer pH 4.5 (Sigma-Aldrich) and immediately injected into the tail vein after 8 hours of fasting. The control rats received instead a citrate buffer of the same volume. STZ-induced diabetic rats were included and retained for the experiments if their blood glucose was greater than 200 mg/dL. Blood glucose was measured using a glucometer (ACCU-CHEK, ADVANTAGE; Roche Diagnostics, Mannheim, Germany). Animals were separated into five groups: 1) diabetes treated with 0.9% normal saline group (DM-NSS; n =10), 2) diabetes treated with curcumin (Cayman Chemical, Ann Arbor, MI) 30 mg/kg dissolved in corn oil (DM-CUR30; n =10), 3) diabetes treated with curcumin 300 mg/kg (DM-CUR300; n =10), 4) control treated with 0.9% normal saline group (CON-NSS; n =10), and 5) control treated with 300 mg/kg bodyweight (CONCUR300; n =10). It is noted that the daily oral feeding of curcumin was started at 6 weeks after the STZ injection since it was shown in our previous study that endothelial dysfunction in STZ-rats occurred 6 weeks after STZ injection.Citation23 The feeding of curcumin was continued for 8 weeks.
Measurement of metabolic parameters
At the end of each experiment, a blood sample from each rat was collected for further plasma glucose and glycosylated hemoglobin (HbA1c) determination, using the enzymatic method and the turbidimetric immunoinhibition method, respectively (Bangkok RIA Laboratory Co. Ltd, Bangkok, Thailand).
Determination of prostanoid levels
PGI2 is rapidly hydrolyzed nonenzymatically from 6-keto- PGF1α. Therefore, in most studies, 6-keto-PGF1αis widely used as an indicator for determining PGI2 production. TXB2, which is the hydrolysis product of TXA2, is commonly used as an indicator of TXA2 production as well.
Under pentobarbital sodium anesthesia, the abdominal cavity was opened via the midline position. The mesentery was exteriorized. The ileocecal portion of the mesentery was carefully spread on a plexiglass chamber and continuously perfused by 1 mL/min Kreb-Ringer buffer solution (37°C, pH 7.4 composition in mmol/L; 135.7 NaCl, 4.7 KCl, 2.52 CaCl2, 1.18 KH2PO4, 1.64 MgSO4.7H2O, and 7.14 NaHCO3) for 15-minute equilibration. After 15 minutes, 1 mL of the mesenteric perfusate was collected immediately for the measurements of existing 6-keto-PGF1αand TXB2 (basal level). The levels of 6-keto-PGF1αand TXB2 were determined by using enzyme immunoassay kits. According to the manufacturer’s protocols (Cayman Chemical), the quantitative levels of 6-keto-PGF1αand TXB2 could be assessed by the standard calibration curves of both substances.
Immunofluorescent staining for PKC-βII
Under pentobarbital sodium anesthesia, the rat mesentery was exposed. A single unbranched small mesenteric artery with a diameter of approximately 100 μm was selected for study. The selected microvessel was then dissected and cleared of connective tissue and briefly rinsed in icecold phosphate-buffered saline (PBS). After collection, the mesenteric arteries were then immediately fixed in 4% paraformaldehyde for 24 hours and were embedded in paraffin. These specimens were then deparaffinized in xylene, rehydrated in graded ethanol and distilled water, and the antigen unmasked with sodium citrate (10 mmol/L, pH 6.0) (Dako, Glostrup, Denmark), followed by a microwave heat source on high power for 3 minutes and then with low power for 10 minutes. After the PBS wash, the nonspecific background was blocked with 3% normal horse serum at room temperature for 20 minutes. Incubation with anti-PKC-βII (1:100 dilutions) (SC-210; Santa Cruz Biotechnology, Santa Cruz, CA) was performed at room temperature for 60 minutes. Anti-α-smooth muscle actin (1:200 dilutions) (Dako) was also used to colocalize microvascular smooth muscle. Sections were then washed in PBS and incubated with the secondary antibody swine anti-rabbit IgG-TRITC (1:50 dilution) (R0156; Dako) for PKC-βII and rabbit anti-mouse IgG-FITC (1:50 dilution) (R0261; Dako) for smooth muscle actin at room temperature for 30 minutes. The arteries received two 3-minute washes in PBS and were covered with mounting medium containing DAPI (4,6-diamidino-2-phenylindole) (Vector Laboratories, Burlingame, CA) for fluorescent counterstaining of nuclei. Labeling of the arteries with secondary antibody alone was used as negative controls. Images were obtained using laser scanning confocal microscopy (E800; Nikon, Tokyo, Japan) to establish the localization of PKC-βII and α-smooth muscle actin in the small mesenteric arteries.
Immunohistochemistry for COX-2
After the collection, the mesenteric arteries were fixed by the same protocol. Paraffin-embedded sections of the mesenteric arteries were sequentially exposed to the solutions as described previously. COX-2 was immunohistochemically detected by incubation with rabbit anti-COX-2 (1:500 dilutions) (RP111; Diagnostic Biosystems, Pleasanton, CA) followed by the appropriate secondary antibody, anti-rabbit IgG horseradish peroxidase.
Immunohistochemistry for NF-κB
Activated NF-κB was detected by incubation with anti- NF-κB p65 rabbit polyclonal antibody (1:150 dilution) (SC-109; Santa Cruz Biotechnology), which was recognized as an epitope accessible only when NF-κB is bound to DNA. This was followed by incubation with a horseradish peroxidase- conjugated secondary antibody. The data were expressed as a positive signal in the vasculature. The stained sections were examined under a microscopy system (Optiphot 2; Nikon) equipped with Nikon Digital Sight DS-Fi1, DS-L2, and 20×objective lens (CF Plan Fluor; Nikon).
Quantitative analysis for the amount of COX-2 and NF-κB p65 was measured using Global Lab Image/2 software (Data Translation, Marlborough, MA). Using the histogram tool of the software, both the lowest and highest intensity values in the brown immunoreactive area were determined, and then the total number of pixels which had the intensity within those threshold values was obtained for small regions of interest (ROI) (45 ×45 pixels). Means ±standard error of mean (SEM) of the total number of pixels of immunoreactions of COX-2 or NF-κB p65 were calculated by averaging the small ROI of each video frame.
where, xi is the total number of brown-precipitate pixels in each ROI, and n is the total number of ROI in each video frame.
Statistical analysis
All data were expressed as mean ±SEM. For comparison among groups of animals, one-way analysis of variance (ANOVA) was used. Also Tukey’s post-hoc test was used when comparing the difference of the means between the diabetic rats and the controls, and between the diabeticuntreated and diabetic-treated animals. The statistical probabilities, P < 0.001, P <, 0.01, and P <, 0.05, were considered statistically significant. The data were analyzed with the SPSS program for Windows (version 16.0; SPSS Inc., Chicago, IL).
Results
In , bodyweight, mean arterial blood pressure (mABP), plasma glucose, and HbA1c values are shown for each group. The blood glucose and percentage of HbA1c of all diabetic groups (DM-NSS, DM-CUR30, and DMCUR300) were significantly higher than those of the control (P < 0.01). Interestingly, among DM-NSS, DM-CUR30, and DM-CUR300, there was a significant difference in percentage of HbA1c (P <0.05).
Table 1 Bodyweight and mean arterial blood pressure, blood glucose, and percentage HbA1c
Only the DM-NSS group had a significantly increased mABP when compared with the CON-NSS group (P <0.01). Conversely, the levels of mABP of DM-CUR30 and DMCUR300 were significantly lower than those of age-matched untreated diabetic rats (P <0.05).
Supplementation of curcumin 30 and 300 mg/kg per day could lower blood glucose in the diabetic group down to 18.73% and 30.26%, respectively. This antidiabetic action of curcumin is confirmed by previous reports, which show that curcumin can mediate its hypoglycemic effect through the stimulation of the pancreas to produce and secrete insulin.Citation24–Citation27 Treatment of streptozotocin in our diabetes model causing beta cell destruction leads to hyperglycemia with hypoinsulinemia. However, limited B-cells regeneration may occur, allowing cellular responses to curcumin activation. However, the curcumin supplementation could not reduce blood glucose back to its normal state. The blood glucose and HbA1c of both DMCUR30 and DM-CUR300 still significantly increased when compared with CON-NSS. In other words, the antidiabetic effect of curcumin is not great enough to be used alone.
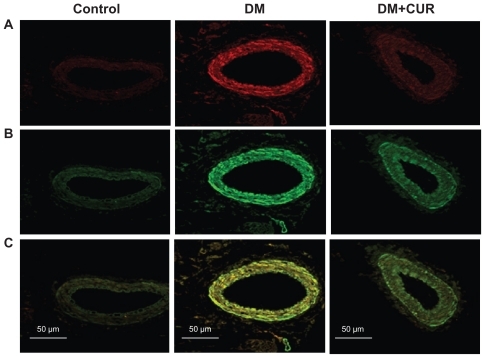
In , immunofluorescent staining of small mesenteric arteries displays a strong signal for PKC-βII in the diabetic rats (). Double-immunofluorescent staining with specific antibodies for PKC-βII and α-smooth muscle actin shows that the diabetic mesenteric arteries contained PKC- βII within the vascular smooth muscle cells (). Interestingly, supplementation with curcumin (300 mg/kg) could reduce PKC-II expression in DM-CUR 300, with results looking similar to the control staining results. It is noted that the negative control displays a minimal detectable fluorescence when the secondary antibodies were used alone. These results suggest that mesenteric vessels in the DM-NSS have higher levels of PKC-βII both in the endothelium and the smooth muscle cells.
Figure 1 Co-immunofluorescent staining of protein kinase C (PKC)-βII and vascular smooth muscle cells in mesenteric arteries (diameter =100–120 μm) taken from control, diabetes mellitus (DM), and DM curcumin 300 (DM-CUR 300) groups. Double-immunofluorescent staining was performed to demonstrate PKC-βII in the mesenteric arteries (red fluorescence; A), α-smooth muscle actin (green fluorescence; B), and merged image (yellow; C). Magnification: ×400.
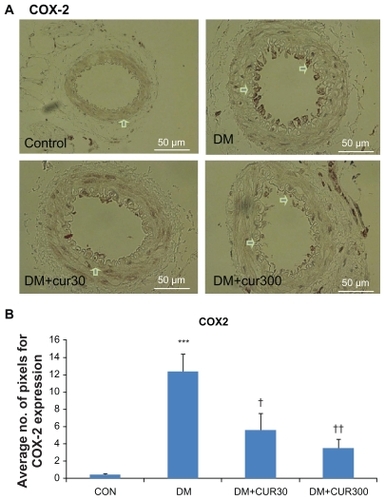
demonstrate that COX-2 expression in the small mesenteric artery of the DM rats was markedly increased when compared with the control. Curcumin supplementation reduced COX-2 expression in the mesenteric microvessels significantly when using low and high doses of curcumin (DM-CUR30, P < 0.05; DM-CUR300, P < 0.01); although these reduced COX-2 expressions do not seem to completely eliminate the COX-2 expression in diabetic arteries.
Figure 2 A) Immunostaining for cyclo-oxygenase (COX)-2 in small mesenteric arteries. Sections of arterial segment show the endothelial cell layer inwards. Positive immunoreactions are observed as a brown precipitate.
Abbreviations: CON-NSS, control with 0.9% normal saline; DM-CUR30, diabetes-treated with curcumin 30 mg/kg; DM-CUR300, diabetes-treated with curcumin 300 mg/kg; DM-NSS, diabetes treated with 0.9% normal saline; SEM, standard error of the mean.
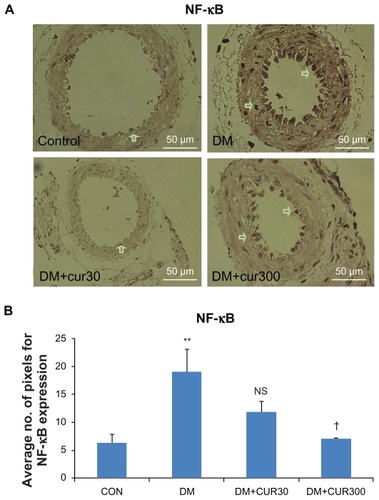
demonstrate that NF-κB p65 was expressed more in the diabetic small mesenteric arteries when compared with the control. DM-CUR300 seemed to attenuate this diabetes-induced NF-κB p65 upregulation significantly (P <0.05).
Figure 3 A) The expression of nuclear factor-κB p65 (NF-κB p65) in small mesenteric arteries in the control (left upper panel), DM (right upper panel), DM-CUR30 (left lower panel) and DM-CUR300 (right lower panel) rats. Positive immunoreactions are observed as a brown precipitate. Magnification: ×400. B) Expression of NF-κB in the endothelium layer in small mesenteric arteries using image analysis (Global Lab Image/2 software) measurement in the DM-NSS, DM-CUR30, and DM-CUR300 groups.
Abbreviations: DM-CUR30, diabetes-treated with curcumin 30 mg/kg; DM-CUR300, diabetes-treated with curcumin 300 mg/kg; DM-NSS, diabetes treated with 0.9% normal saline; NF-κB, nuclear factor-κB; SEM, standard error of the mean.
indicates that 6-keto-PGF1αrelease was significantly decreased in the DM group (407.6 ±37 pg/mL) when compared with the control (536.9 ±53.3 pg/mL) (P < 0.05). There was a signif icantly increased TXB2 production in the DM group (58.4 ±5.8 pg/mL) when compared with their respective controls (29.4 ±4.3 pg/mL) (P <0.05). In addition, there was a significantly decreased 6-keto- PGF1α/TXB2 ratio in the DM group (7.0) when compared with their respective controls (18.4) (P < 0.05). It is to be noted that in the prostanoids ratio study, only the high-dose supplementation of curcumin (DM-CUR300) was conducted. This high dose could be a more effective dose of curcumin supplementation to reduce the diabetes-induced vascular NF-κB p65 and COX-2 upregulation.
Table 2 Means ±SEM of basal levels of 6-keto-PGF1α and TXB2 together with the 6-keto-PGF1α/TXB2 ratios in the mesenteric arteriolar bed of CON-NSS, DM-NSS, and DM-CUR300 groups
Discussion
In the present study, we have shown that the effects of curcumin supplementation on attenuating diabetes-induced endothelial dysfunction are closely associated with its antioxidant and anti-inflammatory properties. Two doses of curcumin were used in this study. The rationale of these low and high doses was based on the literature reviews that each antioxidant and anti-inflammatory action was indicated by different optimal doses.Citation20 A high dose of curcumin supplementation seems to prevent the dysregulation for the multiple signaling pathways induced by diabetes. As such, the idea that curcumin may help prevent diabetic vascular complications was confirmed by the present finding that supplementation with a high dose of curcumin, 300 mg/kg per day, appears to improve the diabetic endothelial functions which was shown by the decrease in mABP and the suppression of COX-2 and NF-κB at the mesenteric arterial wall.
At present, the mechanisms of increased oxidative stress in diabetes are demonstrated on a multifactor basis.Citation6–Citation9 The pathway of DAG-PKC is well documented as one mechanism of increasing oxidative stress since such activation leads to altered NADPH (nicotinamide adenine dinucleotide phosphate) oxidase. From our previous report, it is shown that curcumin can decrease superoxide at the mesenteric vascular wall as demonstrated by the oxygen radical-sensitive fluorescent probe, hydroethidine.Citation28 It was shown that supplementation with a high dose of curcumin could decrease malondialdehyde, the common indicator of oxidative stress, by almost 50% in the STZ-rat model.Citation29,Citation30
The interaction between NO and O2 •−occurred at an extremely rapid rate, three times faster than the reaction rate between O2 •−and SOD.Citation31 Therefore, this hyperglycemiainduced O2 •−may quench NO and directly contribute to the dysfunction of several consequently endothelium-dependent processes. In addition, it is well known that diabetes-induced ROS play a key role in enhancing inflammation through the activation of stress kinases and redox sensitive transcription factors such as NF-κB. NF-κB, a redox-sensitive factor and a key regulator of antioxidant enzymes, can initiate transcription of many genes involved in inflammatory and immune response. In unstimulated cells, NF-κB is a heterodimeric complex that is sequestered in the cytoplasm by interacting with the inhibitory I-κB family. When these cells are stimulated, I-κB is phosphorylated with subsequent release of NF-κB resulting in the translocation of NF-κB from the cytoplasm to the nucleus where it can initiate the expression of various target genes.Citation31,Citation32
In this study, the results showed that the expression of NF-kB p65 in the small mesenteric arterial wall was increased significantly in the DM group. The results of quantitative comparison using image analysis (Global Lab II software) indicated that only the high doses of curcumin supplementation (300 mg/kg bodyweight) could decrease this upregulated NF-κB p65 expression significantly (P < 0.05) ().
It was reported that NF-κB was suppressed by curcumin through inhibiting the activity of I-κB kinase (IKK).Citation33 Oxidative stress activates NF-κB-mediated transcription of proinflammatory mediators either through the activation of its activating inhibitor, IKK, or the enhanced recruitment or activation of transcriptional co-activators. In cancer-related inflammation, although numerous different pathways are activated during the inflammatory response, NF-κB is thought to be of the most importance.Citation34
Activation of peroxisome proliferator-activated receptors gamma (PPARγ) by curcumin resulted in inhibition of NF-κB trans-activating activity and increased expression of PPARγ at both the transcriptional and translational levels in activated hepatic stellate cells.Citation35 In principle, curcumin has been widely demonstrated to have potent antioxidant activities. It was reported that curcumin could increase antioxidant glutathione levels by induction of glutamate cysteine ligase and act as an anti-inflammatory agent through inhibition of NF-κB signaling.Citation36
This is the first report showing that curcumin, a polyphenol, has beneficial effects on significantly decreased COX-2 expression in the diabetic endothelial layer. Although in diabetes, the changes in vascular prostanoid production are clearly indicated in particular in association with diabetic vasculopathy, the role of COX-2 in diabetes is not very clear.Citation37,Citation38 an in vivo relevance to diabetic complications was suggested by the observation of high levels of COX-2 mRNA presented in monocytes from type 1 or type 2 diabetic patients, but not from normal volunteers.Citation39 Moreover, it has been shown that this high glucose-induced COX-2 mRNA expression was implicated by the involvement of multiple pathways, including the p38 MAPK, PKC, and NF-κB. As shown in the present findings, high glucose enhanced the increase of PKC, COX-2, and NF-κB, particularly at the endothelial lining of mesenteric arteries. Moreover, the results of quantitative comparison using image analysis (Global Lab II software) showed that the high doses of curcumin supplementation could significantly decrease COX-2 (P < 0.05) ().
In our study, the unstimulated (basal) level of 6-keto- PGF1α (stable metabolite of PGI2) in the diabetic rats was markedly reduced compared with those of the control rats. While curcumin enhances 6-keto-PGF1α production up to almost basal level of control rats (), the basal levels of TXB2 (stable metabolite of TXA2) in diabetic rats were significantly elevated as compared with those of the control rats. These findings may have implications for the existing role of diabetes-induced COX-2 expression, since the prostaglandin which may be responsiblefor an enhanced production of TXA2 while reducing the production of PGI2. In addition, we also found a marked COX-2 immunostaining in the mesenteric arteries of diabetic animals, which was localized in the endothelial layer of the arterial wall (). These results are in accordance with other studies obtained from both diabetic animal modelsCitation40 and human diabetes.Citation41
Interestingly, supplementation with curcumin could improve the ratio of prostanoid change to higher PGI2 than TXA2 level (). According to prostanoid levels, slight immunostaining for COX-2 was detected in the arteries of diabetic rats supplemented with curcumin (). Thus, curcumin has been found to be effective in inhibiting TXA2 synthesis in inflammation by modulating the COX-2-pathway.
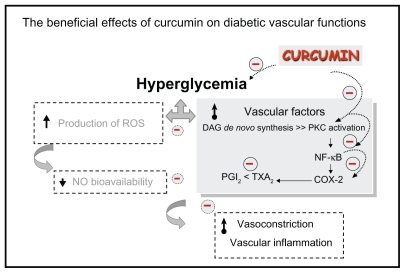
In conclusion, our findings provide in vivo evidence that curcumin supplementation (300 mg/kg) significantly improved diabetes-induced endothelial dysfunction related to its potential for COX-2 and NF-κB suppression and PKC inhibition, and resulted in an improved ratio of prostanoid products (as summarized in ).
Figure 4 The beneficial effects of curcumin on diabetic vascular functions. Curcumin supplementation (300 mg/kg bodyweight) improved diabetes-induced vascular dysfunction associated with its potential to reduce blood sugar, COX-2 and NF-κB suppression, PKC inhibition, and improve the ratio of prostanoid products PGI2 and TXA2.
Indeed, it is important to note that ruboxistaurin (LY333531) mesylate, a PKC-isozyme-selective inhibitor, is in Phase III clinical trials in patients with type 1 and type 2 DM to determine its efficacy in preventing the development of diabetic microvascular complications.Citation42 In addition, clinical trials using up to 8000 mg curcumin per day for 3 months have shown no toxicity.Citation43 Therefore, it is possible that curcumin supplementation might be beneficial for diabetic patients by improving microvascular functions and preventing the consequences of cardiovascular complications. Therefore, curcumin might be considered as a phytoceutical agent to be used for the treatment of diabetic vascular complications in diabetes patients in the future.
Authors’ contributions
SR participated together with co-authors in the design of this study. The experiments were carried out by SR, SP, and NT. PR was the immunotechnique assistant. SR interpreted and analyzed the data. SR prepared the initial draft of the manuscript, and SP read and revised the manuscript. All authors read and approved the manuscript.
Acknowledgements
This study was supported by Ratchadaphiseksomphot Fund, Faculty of Medicine, the 90th Anniversary of Chulalongkorn University Fund, Graduate School, Chulalongkorn University.
Disclosure
The authors declare that they have no competing interests related to this study.
References
- Wild S Roglic G Green A Sicree R King H Global prevalence of diabetes: estimates for the year 2000 and projections for 2030 Diabetes Care 2004 27 1047 1053 15111519
- World Health Organization Mortality and country Fact Sheet 2006 Available from: http://www.who.int/entity/healthinfo/statistic Accessed Apr 2006
- VanderJagt DJ Harrison JM Ratliff DM Hunsaker LA Vander Jagt DL Oxidative stress indices IDDM subjects with and without long-term diabetic complications Clin Biochem 2001 34 265 270 11440725
- Bunnag SC Implications of microcirculation-research based information on prevention and treatment of diabetes mellitus type 2: a perspective Clin Hemorheol Microcirc 2006 34 43 50 16543616
- Chakravarthy U Hayes RG Stitt AW McAuley E Archer DB Constitutive nitric oxide synthase expression in retinal vascular endothelial cells is suppressed by high glucose and advanced glycation end products Diabetes 1998 47 945 952 9604873
- Muniyappa R Srinivas PR Ram JL Walsh MF Sowers JR Calcium and protein kinase C mediate high-glucose-induced inhibition of inducible nitric oxide synthase in vascular smooth muscle cells Hypertension 1998 31 289 295 9453318
- Sharpe PC Liu WH Yue KK Glucose-induced oxidative stress in vascular contractile cells: comparison of aortic smooth muscle cells and retinal pericytes Diabetes 1998 47 801 809 9588453
- Tesfamariam B Brown ML Deykin D Cohen RA Elevated glucose promotes generation of endothelium-derived vasoconstrictor prostanoids in rabbit aorta J Clin Invest 1990 85 929 932 2312734
- Cosentino F Eto M de Paolis P van der Loo B Bachschmid M Ullrich V High glucose causes upregulation of cyclooxygenase-2 and alters prostanoid profile in human endothelial cells: role of protein kinase C and reactive oxygen species Circulation 2003 107 1017 1023 12600916
- Quilley J Chen YJ Role of COX-2 in the enhanced vasoconstrictor effect of arachidonic acid in the diabetic rat kidney Hypertension 2003 42 837 843 12900435
- Davidge ST Prostaglandin H synthase and vascular function Circ Res 2001 89 650 660 11597987
- Chattopadhyay I Biswas K Bandyopadhyay U Banerjee RK Turmeric and curcumin: biological actions and medicinal applications Curr Sci 2004 87 44 53
- Sharma RA Steward WP Gescher AJ Pharmacokinetics and pharmacodynamics of curcumin Aggarwal BB Young-Joon S Shishodia S Targets and Therapeutic Uses of Curcumin in Health and Disease New York, NY Springer 2007
- Farhangkhoee H Khan ZA Chen S Chakrabarti S Differential effects of curcumin on vasoactive factors in the diabetic rat heart Nutr Metab (Lond) 2006 3 27 16848894
- Adam BK Cai J Armstrong J EF24, a novel synthetic curcumin analog, induces apoptosis in cancer cells via a redox-dependent mechanism Anticancer Drugs 2005 16 263 275 15711178
- Khopde SM Priyadarsini KI Venkatesan N Rao MNA Free radicail scavenging ability and antioxidant efficiency of curcumin and its substituted analogue Biophys Chem 1999 80 85 91 17030320
- Mahesh T Sri Balasubashini MM Menon VP Photo-irradiated curcumin supplementation in streptozotocin-induced diabetic rats: effect on lipid peroxidation Therapie 2004 59 639 644 15789828
- Meghana K Sanjeev G Ramesh B Curcumin prevents streptozotocin-induced islet damage by scavenging free radicals: a prophylactic and protective role Eur J Pharmacol 2007 577 183 191 17900558
- Murugan P Pari L Effect of tetrahydrocurcumin on lipid peroxidation and lipids in streptozotocin-nicotinamide-induced diabetic rats Basic Clin Pharmacol Toxicol 2006 99 122 127 16918712
- Aggarwal BB Harikumar KB Potential therapeutic effects of curcumin, the anti-inflammatory agent, against neurodegenerative, cardiovascular, pulmonary, metabolic, autoimmune and neoplastic diseases Int J Biochem Cell Biol 2009 41 1 40 59 18662800
- Plummer SM Hollooway KA Manson MM Inhibition of cyclooxygenase 2 expression in colon cells by the chemopreventive agent curcumin involves inhibition of NF-kappaB activation via the NIK/IKK signalling complex Oncogene 1999 18 6013 6020 10557090
- Chun KS Keum YS Han SS Song YS Kim SH Surh YJ Curcumin inhibits phorbol ester-induced expression of cyclooxygenase-2 in mouse skin through suppression of extracellular signal-regulated kinase activity and NF-kappaB activation Carcinogenesis 2003 24 1515 1534 12844482
- Sridulyakul P Chakraphan D Patumraj S Vitamin C supplementation could reverse diabetes-induced endothelial cell dysfunction in mesenteric microcirculation in STZ-rats Clin Hemorheol Microcirc 2006 34 315 321 16543652
- Halim E Hussain MA Hypoglycemic, hypolipidemic and antioxidant properties of combination of curcumin from Curcuma Longa, Linn, and partially purified product from Abroba Augusta, Linn, in streptozotocin induced diabetes Indian J Clinical Biochem 2002 17 33 43
- Sharma S Kulkarni SK Chopra K Curcumin, the active principle of turmeric (Curcuma longa), ameliorates diabetic nephropathy in rats Clin Exp Pharmacol Physiol 2006 33 940 945 17002671
- Patumraj S Wongeakin N Jariyapongskul A Futrakul N Bunnag S Combined effects of curcumin and vitamin C to protect endothelial dysfunction in the iris tissue of STZ-induced diabetic rats Clin Hemorheol Microcirc 2006 35 481 489 17148847
- Menon VP Sudheer AR Antioxidant and anti-inflammatory properties of curcumin Adv Exp Med Bio 2007 595 105 125 17569207
- Rungseesantivanon S Thengchaisri N Ruangvejvorachai P Patumraj S Curcumin supplementation could improve diabetes-induced endothelial dysfunction associated with decreased vascular superoxide production and PKC inhibition BMC Complement Altern Med 2010 10 57 20946622
- Wongeakin N Sridulyakul P Jariyapongskul A Suksamrarn A Patumraj S Effects of curcumin and tetrahydrocurcumin on diabetes induced endothelial dysfunction Afr J Biochem Res 2009 3 259 265
- Wolin MS Interactions of oxidants with vascular signaling systems Arterioscler Thromb Vasc Biol 2000 20 6 1430 1442 10845855
- Yamamoto Y Gaynor RB Therapeutic potential of inhibition of the NF-κB pathway in the treatment of inflammation and cancer J Clin Invest 2001 107 2 135 142 11160126
- Gilmore TD The Rel/NF-κB signal transduction pathway: introduction Oncogene 1999 18 49 6842 6844 10602459
- Pan MH Lin-Shiau SY Lin JK Comparative studies on the suppression of nitric oxide synthase by curcumin and its hydrogenated metabolites through down-regulation of IκB kinase and NFκB activation in macrophages Biochem Pharmacol 2000 60 1665 1676 11077049
- Philip M Rowley DA Schreiber H Inflammation as a tumor promoter in cancer induction Semin Cancer Biol 2004 14 433 439 15489136
- Xu J Fu Y Chen A Activation of peroxisome proliferator-activated receptor-γ contributes to the inhibitory effects of curcumin on rat hepatic stellate cell growth Am J Physiol Gastrointest Liver Physiol 2003 285 1 G20 G30 12660143
- Biswas SK McClure D Jimenez LA Megson IL Rahman I Curcumin induces glutathione biosynthesis and inhibits NF-κB activation and interleukin-8 release in alveolar epithelial cells: mechanism of free radical scavenging activity Antioxid Redox Signal 2005 7 32 41 15650394
- Cohen RA The role of nitric oxide and other endothelium-derived vasoactive substances in vascular disease Prog Cardiovasc Dis 1995 38 105 108 7568902
- Shanmugam N Kim YS Lanting L Natarajan R Regulation of cyclooxygenase- 2 expression in monocytes by ligation of the receptor for advanced glycation end products J Biol Chem 2003 278 34834 34844 12837757
- Shanmugam N Gaw Gonzalo IT Natarajan R Molecular mechanisms of high glucose-induced cyclooxygenase-2 expression in monocytes Diabetes 2004 53 795 802 14988266
- Bagi Z Erdei N Toth A Type 2 diabetic mice have increased arteriolar tone and blood pressure: enhanced release of COX-2- derived constrictor prostaglandins Arter Thromb Vasc Biol 2005 25 1610 1616
- Szerafin T Erdei N Fulop T Increased cyclooxygenase-2 expression and prostaglandin-mediated dilation in coronary arterioles of patients with diabetes mellitus Circ Res 2006 99 e12 e17 16917094
- Wei Z Xiao-Li W Kathryn GL Hon-Chi L Inhibition of protein kinase C protects against diabetes-induced impairment in arachidonic acid dilation of small coronary arteries J Pharmacol Exp Ther 2006 319 199 207 16861398
- Chainani-Wu N Safety and anti-inflammatory activity of curcumin: a component of tumeric (Curcuma longa) J Altern Complement Med 2003 9 161 168 12676044