
Abstract
Purpose
To investigate the main molecular resistance mechanisms to fluoroquinolones (FQs) in Pseudomonas aeruginosa and also to investigate the effect of time and concentration on mutations in resistance genes.
Materials and methods
The clinical isolates of P. aeruginosa which are sensitive to ciprofloxacin (CIP) or levofloxacin (LEV) were collected. The isolates were incubated with different concentrations of CIP or LEV for 5 days and the minimal inhibitory concentrations (MICs) of CIP, LEV and ofloxacin (OFX) were measured. The MIC of FQs to P. aeruginosa was measured by the agar dilution method. FQ resistance determining regions of gyrA, gyrB, parC and parE were amplified by PCR, and mutations in four genes were explored using sequence analysis with the Snapgene software. The relative expression levels of two efflux pumps genes (mexA and mexE) were measured by quantitative reverse transcription PCR.
Results
A total of eleven isolates were collected from the Second Hospital of Shanxi Medical University. Amino acid alterations in gyrA and gyrB were mainly detected in resistant mutants, and the percentage of strains with amino acid alterations in gyrB was significantly higher than that in gyrA (P<0.001). MICs of strains with mutations both in gyrA and gyrB were not significantly higher than those of strains with mutations only in gyrB (P>0.05). No amino acid alterations were detected in genes of parC and parE. In both gyrA and gyrB, the number of amino acid alterations increased with incubation time prolonged and increased with increasing incubation concentration.
Conclusion
CIP was more competent than LEV in making P. aeruginosa resistant to in vitro selection. Mutations occurring in gyrB played an important role in FQ resistance of P. aeruginosa in vitro selection.
Introduction
Pseudomonas aeruginosa is a clinically important opportunistic pathogen with high pathogenicity and high mortality. A recent report from the National Healthcare Safety Network (NHSN) reported P. aeruginosa to be one of the six most common nosocomial pathogens associated with healthcare-associated infections, and the resistance rate of P. aeruginosa to multiple drugs (aminoglycosides, extended-spectrum cephalosporin, carbapenems) exceeded 15%.Citation1 Infections caused by P. aeruginosa are difficult to treat due to multiple resistance mechanisms.Citation2,Citation3 The resistance mechanisms of P. aeruginosa include natural resistance, acquired resistance and adaptive resistance.Citation4,Citation5 These complex and various resistance mechanisms make the infections caused by P. aeruginosa life-threatening. Meanwhile, the resistance of P. aeruginosa to many antimicrobial drugs is emerging worldwide as a public threat.Citation6
Fluoroquinolones (FQs) are an important class of antimicrobial drugs used to treat infections caused by P. aeruginosa, among which ciprofloxacin (CIP) or levofloxacin (LEV) is used frequently.Citation7–Citation9 FOs act by inhibiting the intracellular targets, DNA gyrase and topoisomerase IV for inhibiting DNA replication. DNA gyrase tends to be the primary target in gram-negative bacteria, whereas topoisomerase IV is preferentially inhibited by FQs in gram-positive bacteria.Citation8,Citation10,Citation11 But the resistance of P. aeruginosa to CIP and LEV has emerged. Data from China Antimicrobial Surveillance Network showed that the resistance rate of P. aeruginosa to CIP had reached to 14.8% in 2017.Citation12 Also, a recent report from the NHSN showed that the resistance rate of P. aeruginosa to CIP and LEV exceeded 30% within American hospitals.Citation1 Studying the specific resistance mechanisms of FQs to P. aeruginosa is helpful in choosing the appropriate antibiotics, avoiding the emergence of resistant strains and developing new antimicrobial drugs.
In theory, the molecular resistance mechanisms of FQs to P. aeruginosa mainly involve gene mutations of gyrA and gyrB which encode DNA gyrase and parC and parE which encode topoisomerase IV. But many previous studies revealed that gyrA mutations played a crucial role in FQ resistance in P. aeruginosa and parC mutation was associated with the development of high-level resistance.Citation7,Citation9,Citation13–Citation17 A few studies reported gyrB mutations in FQ-resistant P. aeruginosa. Also, the relationship between gyrB mutations and drug resistance in the existing literatures is not explicit.Citation18–Citation20 This raises the question of whether gyrB mutations of P. aeruginosa play a role in resistance to FQs. To address this, in the current work, we incubated P. aeruginosa in vitro and detected the mutations in FQ resistance determining regions (FRDRs) of gyrA, gyrB, parC and parE. In addition, during the clinical treatment of infection caused by P. aeruginosa, the patients were initially sensitive to FQs, but the sensitivity was reduced after a period of use, resulting in failure of the treatment. Based on this problem, in this study, we set different incubation time periods and concentrations in order to find out the effect of time and concentration on mutations in resistance genes.
Materials and methods
Collection of clinical isolates
Clinical nonrepetitive isolates were collected from the Second Hospital of Shanxi Medical University. All isolates were identified by VITEK-2 Compact system (BioMerieux Italia S.p.A) and were stored in liquid medium of brain heart infusion at −86°C. These isolates and P. aeruginosa ATCC15692 were defined as the original strains. ATCC15692 was obtained from the First Hospital of Wuhan and was numbered as PA12 in this study.
P. aeruginosa was isolated from different patients for this research. This research was approved by the ethics committee of the Second Hospital of Shanxi Medical University. We had hidden the patients’ information when collecting the isolates. Also, the patients’ written informed consent was exempt, which was also approved by the ethics committee.
Measurement of minimal inhibitory concentration (MIC)
The susceptibility of original strains to CIP, LEV and ofloxacin (OFX) was determined by the agar dilution method, with the concentration of each antibacterial agent ranging from 0.0625 to 256 µg/mL. The results of MICs were interpreted according to the American Clinical and Laboratory Standards Institute 2017.Citation21 Each strain was separately incubated in Mueller–Hinton agars (MHAs; purchased from the British company Oxoid) containing four different concentrations of CIP or LEV. The four incubation concentrations referred to 0.5× MIC, 1× MIC, 2× MIC and 4× MIC, respectively, depending on the MICs of 12 original strains. Each isolate was incubated at each concentration for 5 days. Finally, the strains incubated for 1, 3 and 5 days were stored at −86°C. Similarly, the MICs of strains after incubation were measured by the agar dilution method.
Incubation experiment conducted in vitro
Each original strain was first inoculated in blood culture for recovery and then incubated in a thermotank at 37°C for 18–24 hours. Using cotton swab, a single bacterial colony was picked into a test tube containing normal saline and the bacterial suspensions were adjusted to 3.0 McFarland turbidity standard (MCF). The bacteria were separately incubated on MHAs containing CIP of four different concentrations. The incubation was conducted at 37°C for 1 day, and then the content transferred to the new MHAs containing the same concentration of CIP for the next day. This transfer continued until the incubation reached the fifth day.
The method of incubating strains with LEV was the same as incubating strains with CIP. The strains incubated with CIP were defined as the CIP group, and the strains incubated with LEV as the LEV group.
Conventional PCR amplification and DNA sequencing of FRDRs
The PCR amplification of gyrA, gyrB, parC and parE of each strain was performed in a final volume containing the following reaction mixture: Taq PCR Master Mix 25 µL, bacterial suspensions 1 µL (1.5 MCF), forward primer (10 µmol/L) 2 µL, reverse primer (10 µmol/L) 2 µL; finally, sterilized ddH2O was added and the mixture made up to 50 µL. The PCR primers of the four genes were shown in . Amplification reaction was performed on a thermal cycler (Lifepro thermal cycler PCR; Bioer, Hangzhou, People’s Republic of China) for 30 cycles. The cycle parameters included predegeneration at 95°C for 5 minutes, denaturation at 95°C for 30 seconds, annealing at 58°C for 30 seconds and elongation at 72°C for 40 seconds. PCR products were electrophoresed on a 1%–1.5% agarose gel, visualized by ethidium bromide staining and photographed under ultraviolet light. The PCR products were sent to Sangon Biotech Co., Ltd (Shanghai, People’s Republic of China) for sequencing. The sequencing primers were the same as the PCR primers. The flowchart describing the experimental design is shown in .
Table 1 Primer sequence used in PCR and RT-PCR
Figure 1 The flowchart describing the experimental design.
Abbreviations: MIC, minimum inhibitory concentration; RT-PCR, reverse transcription PCR.
The base sequences and amino acid sequences in the target regions of the four genes were compared with the PAO1 sequences in the GenBank using the Snapgene software. The DNA sequences were compared with the original nucleotide sequences of gyrA (accession number 882880), gyrB (accession number 879230), parC (accession number 879741) and parE (accession number 879897) genes in the GenBank data for PAO1.
Quantitative reverse transcription PCR (RT-PCR)
The expression of two efflux pumps genes (mexA and mexE) was measured by quantitative RT-PCR. The RT-PCR experiment of mexA and mexE of P. aeruginosa was performed in a final volume containing the following reaction mixture: 2× SG Fast qPCR Master Mix 10 µL, 10 µM forward primer 0.4 µL, 10 µM reverse primer 0.4 µL, a certain amount of template DNA, and finally, PCR-grade water added to make it up to 20 µL. The primers of the two genes are listed in . The amplification parameters included 40 cycles of predegeneration at 95°C for 3 minutes, denaturation at 95°Cfor 3 seconds, annealing at 60°C for 3 seconds and elongation at 60°C for 3 seconds.
The ΔΔCT method was used to calculate the relative expression of the two genes, and the results are shown as 2−ΔΔCT [ΔΔCT = (CT target gene − CTGAPDH)experimental group − (CT target gene − CT GAPDH) control group]. Measurement of the expression of the two genes in each strain was repeated twice and its mean was recorded in the final results. The GAPDH was used as a reference gene.
Statistical analysis
MICs were described as geometric mean±lg-1(SlgMIC).Citation22 All data were analyzed by SPSS16.0 software. Effect of the continuous incubation on the MICs to CIP, LEV and OFX was evaluated with the repeated measures ANOVA and least significant difference t-test. Measurement data were compared by independent samples t-test. The enumerated data were analyzed with chi-squared test. P<0.05 was considered statistically significant.
Results
Collection of clinical isolates
In this study, a total of 11 clinical isolates of P. aeruginosa were collected from the Second Hospital of Shanxi Medical University. These 11 isolates were from different specimens such as wound secretion (four isolates), urine (three isolates), blood (two isolates), pleural effusion (one isolate) and ascitic (one isolate). The MICs of the original strains to CIP, LEV and OFX are shown in .
Table 2 MICs of 12 original strains (µg/mL)
Incubation experiment conducted in vitro
MICs under different incubation concentrations and days
The 12 strains were separately incubated by four concentrations of CIP for 5 days. One hundred and thirty-five strains were successfully stored, and 9 strains were abandoned because their bacterial colonies were not seen on the culture medium after incubation for 48 hours. The nine strains abandoned included PA3d1, PA3d3, PA3d5, PA7c1, PA7c3, PA7c5, PA8d1, PA8d3 and PA8d5 (b and d represent the incubation concentrations of 2× MIC and 4× MIC, respectively; 1, 3 and 5 represent the incubation time of 1, 3 and 5 days, respectively). Similarly, 114 strains were finally stored after incubating with LEV. The 30 strains abandoned included PA1d1, PA1d3, PA1d5, PA3b1, PA3b3, PA3b5, PA3d1, PA3d3, PA3d5, PA4d1, PA4d3, PA4d5, PA5d1, PA5d3, PA5d5, PA6d1, PA6d3, PA6d5, PA7d1, PA7d3, PA7d5, PA12b1, PA12b3, PA12b5, PA12c1, PA12c3, PA12c5, PA12d1, PA12d3 and PA12d5 (a, b, c and d represent the incubation concentrations of 0.5× MIC, 1× MIC, 2× MIC and 4× MIC, respectively).
MICs of P. aeruginosa to CIP, LEV and OFX in the two groups are shown in and . According to the breakpoints given in the guidelines of the American Clinical and Laboratory Standards Institute 2017,Citation21 P. aeruginosa was resistant to CIP, LEV and OFX when the MIC was ≥4.00, 8.00 and 8.00 µg/mL, respectively. MICs of P. aeruginosa to CIP showed that in two groups, the MICs increased over time under any concentration and reached the maximum when the incubation reached the fifth day; meanwhile, the MICs increased with the incubation concentration and improved during incubation and reached the maximum when the incubation concentration reached 4× MIC. Also, the MICs to LEV and OFX increased over time and increased with incubation concentration. Unlike CIP, the MICs of P. aeruginosa to OFX reached the maximum on day 3 under any concentration.
Table 3 MICs of P. aeruginosa to CIP, LEV and OFX upon incubation in the CIP group
Table 4 MICs of P. aeruginosa to CIP, LEV and OFX upon incubation in the LEV group
For CIP resistance in P. aeruginosa, on day 1, the CIP group remained sensitive under any concentration, but the LEV group became resistant when the concentration in MHAs was increased to 4× MIC. On day 3, the CIP group became resistant under the concentration of 2× MIC, whereas the LEV group became resistant when the concentration in MHAs increased to 4× MIC. On day 5, the CIP group was resistant to CIP under any concentration, but the LEV group became resistant until the concentration in MHAs increased to 2× MIC. All the above results show that although the LEV group became resistant to CIP earlier than CIP group, the LEV group needed a higher concentration to become resistant to CIP than the CIP group over time, that is, the CIP group became resistant to CIP more easily than the LEV group over time. Similarly, the results of MICs to LEV and OFX showed that the CIP group became resistant to the two drugs more easily than the LEV group over time.
Statistical analysis of MICs to CIP, LEV and OFX
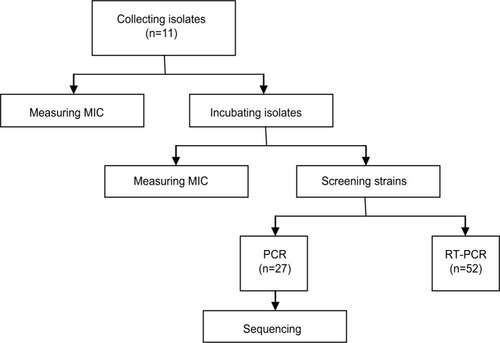
Repeated measures ANOVA showed that in MICs to CIP, the interaction between incubation time and group was statistically significant, that is, the trends in the two groups were significantly different over time (P<0.001). Further simple effect analysis found that, when the group differed, the MIC increments were different (P<0.05). The results above revealed that compared to LEV group, the CIP group’s MICs to CIP appeared to increase faster and had a higher increment in vitro selection (shown in ). Similarly, repeated measures ANOVA showed that in MICs to LEV and OFX, the interaction between incubation time and group was statistically significant, that is, the trends in the two groups were significantly different over time (P<0.001). Also, further simple effect analysis proved that compared to the LEV group, the CIP group’s MICs to LEV and OFX appeared to have a faster increase and higher increment over time (P<0.05).
Figure 2 Simple effect analysis of the relationship between groups and MICs to CIP, LEV and OFX.
Notes: (A) CIP. (B) LEV. (C) OFX. 3/0, 5/0: MICs on day 3 or 5 divided by the original MICs, which means the MIC incremental quantity on day 3 or 5. 3/1, 5/1: MICs on day 3 or 5 divided by the MICs on day 1, which means the MIC incremental speed on day 3 or 5. 1/0: MICs on day 1 divided by the original MICs, which means the MIC incremental quantity and speed on day 1. 5/3: MICs on day 5 divided by MICs on day 3, which means the MIC incremental speed on day 5. There is a statistical difference between the CIP group and the LEV group for the same MIC increment and the same drug (*P<0.05). There is a statistical difference between the CIP group and the LEV group for the same MIC increment and the same drug (**P<0.001).
Abbreviations: CIP, ciprofloxacin; LEV, levofloxacin; MIC, minimum inhibitory concentration; OFX, ofloxacin.
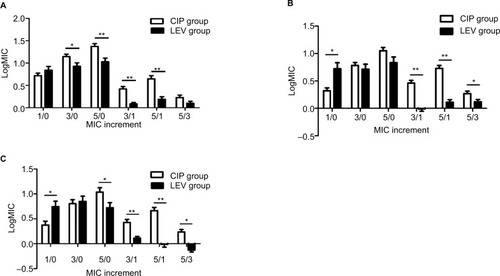
Resistance rate
The results in show that the resistance rate of P. aeruginosa to the three antibiotics increased over time. The CIP group’s resistance rates to the three antimicrobial drugs were gradually higher than those of the LEV group. On day 5, the resistance rates of the CIP group for the three drugs were significantly higher than those of the LEV group (P<0.05), that is, incubating with CIP made the strains resistant more easily than incubating with LEV.
Figure 3 The comparison of resistance rates of strains to CIP, LEV and OFX between the two groups.
Notes: (A) CIP. (B) LEV. (C) OFX. There was a statistical difference between the two groups (*P<0.05).
Abbreviations: CIP, ciprofloxacin; LEV, levofloxacin; MIC, minimum inhibitory concentration; OFX, ofloxacin.
In summary, in analysis of the three drugs in three aspects – the needed drug concentration to make the strains resistant, the MIC increment and the resistance rate – all the obtained results proved that CIP is more competent than LEV in making the strains resistant to in vitro selection. So, strains incubated with CIP were chosen for the next section.
Conventional PCR amplifying and DNA sequencing
After screening, 27 strains were used for PCR amplification and DNA sequencing, including 4 original strains (PA1, PA10, PA11 and PA12) and 23 strains incubated with CIP (). By the PCR assay, the presence of gyrA, gyrB, parC and parE was detected in 418, 510, 357 and 592 bp, respectively. By comparing the results with the corresponding nucleotide sequences of PAO1 in GenBank, amino acid alterations in gyrA and gyrB were mainly detected in these FQ-resistant strains. No amino acid alterations were detected in the genes parC and parE. According to the mutant sites of amino acid in gyrA and gyrB, the results of DNA sequencing are presented in and .
Table 5 MICs of 23 strains used for sequencing (µg/mL)
Table 6 Amino acid alterations of P. aeruginosa in gyrA
Table 7 Amino acid alterations of P. aeruginosa in gyrB
Our results showed mutations in gyrB were detected on day 1, whereas the mutations in gyrA were detected on day 3, that is, gyrB had an earlier mutation than gyrA. Besides, amino acid alterations in gyrA were detected in five strains (21.74%) and amino acid alterations in gyrB were detected in 23 strains (100%). The percentage of strains with mutations in gyrB was significantly higher than the percentage of strains with mutations in gyrA (P<0.001). Noteworthy, five strains had amino acid alterations in both gyrA and gyrB, but the geometric mean of MICs of these five strains were not significantly higher than that of strains with amino acid alterations only in gyrB (P>0.05). What is more, the number of amino acid alterations in gyrB (eight alterations) was more than that in gyrA (five alterations). All of the above results showed that mutations in gyrB played a more important role in FQ resistance to P. aeruginosa.
In both gyrA and gyrB, the number of amino acid alterations increased with time prolonged. Noteworthy, amino acid alteration in gyrA in one strain occurred on day 3 under 4× MIC, whereas in other strains it occurred on day 5 at concentrations below 4× MIC, that is, the strains showed gyrA mutations earlier when incubated with a higher concentration of CIP. From another perspective, in both gyrA and gyrB, the number of amino acid alterations increased with increasing incubation concentration. Noteworthy, no amino acid alteration in gyrA was found under the concentration of 1× MIC.
The amino acid alterations in gyrA mainly occurred at codon 83 (Thr83→Ile). The amino acid alterations in gyrB mainly occurred at codon 372 (Ala372→Val, Ala372→Leu), 424 (Ile424→Leu), 464(Leu→Ile) and 483(Glu483→Asp). Further analysis of the data revealed that a total of 23 strains had mutations in gyrB, of which 1 strain possessed a single amino acid alteration for Ala 372, 8 strains possessed two amino acid alterations for Ala 372 and Ile 424 and the remaining 14 strains possessed all the four above-mentioned amino acid alterations. In addition, other minor novel alterations were detected in gyrA and gyrB.
Efflux pumps expression
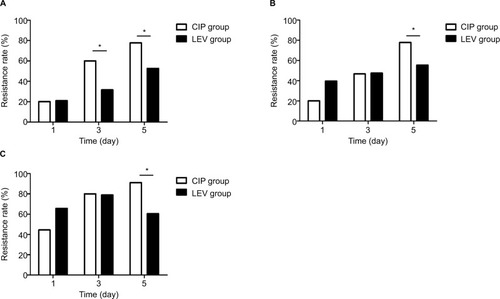
A total of 52 strains of P. aeruginosa were screened out, including 4 original isolates (PA1, PA10, PA11 and PA12) and 48 strains incubated with CIP. The results showed that the efflux pumps were not highly expressed (). Overexpression of efflux pumps was found to be present if the relative expression of mexA was higher than 3.00 and the relative expression of mexE was higher than 10.00.Citation23
Figure 4 The relative expression of two efflux pumps genes – MexA and MexE.
Notes: (A) MexA. (B) MexE. The efflux pump was highly expressed if the relative expression of MexA was higher than 3.00. The efflux pump was highly expressed if the relative expression of MexE was higher than 10.00.
Abbreviation: MIC, minimum inhibitory concentration.
Discussion
After incubating for 5 days under different drug concentrations, the effect of incubating with CIP on MICs and resistance was different from incubating with LEV. For the three drugs (CIP, LEV and OFX), the strains incubated with LEV needed a higher concentration to become resistant than the strains incubated with CIP over time. MICs of strains incubated with CIP showed faster increase over time and the final increment was higher. Also, the resistance rates of strains incubated with CIP became gradually higher than those of strains incubated with LEV over time. After comparisons in three aspects, CIP appeared to have stronger ability to make the strains resistant over time in vitro. This may indicate that LEV is superior to be recommended to treat infection caused by P. aeruginosa in terms of resistance patterns demonstrated in this analysis.
MIC refers to the lowest concentration of antimicrobial drugs that inhibit bacterial growth. The mutant prevention concentration (MPC) refers to the drug concentration at which the first-step resistant mutant has zero growth. If the drug concentration surpasses the MPC, the emergence of resistance is expected to be limited. The mutant selection window (MSW) refers to the range of drug concentration between the MIC and MPC, that is, within this window, resistant mutants will be selected under antimicrobial selective pressure.Citation24–Citation26 The MPCs of P. aeruginosa were 2.00 and 8.00 µg/mL for CIP and LEV, respectively.Citation27,Citation28 In this study, the incubation concentration ranged from 0.03125 to 2 µg/mL and from 0.0625 to 8 µg/mL for CIP and LEV, respectively. Except for the concentration of 0.5× MIC, other incubation concentrations were within the MSW. On one hand, our results showed the MICs increased over time, and resistant mutants had been selected on days 1, 3 and 5. On the other hand, our results showed the MICs increased with increase in the incubation concentration, and resistant mutants had been selected under the concentration of 1× MIC, 2× MIC and 4× MIC. It is worth noting that resistant mutants had also been selected under the concentration of 0.5× MIC, that is, sub-MIC can also be selected for resistant mutants. This has been reported in some literatures.Citation24,Citation26,Citation29 These results prove that the emergence of resistance was associated with time and the concentration of antimicrobial drugs. This suggests that we should pay attention to the dosage and course of drugs during clinical treatment of infection. In addition, after incubating with CIP or LEV, strains were not only resistant to CIP or LEV, but also resistant to OFX, that is, there was cross-resistance between FQs.Citation30,Citation31 This indicates that if P. aeruginosa is resistant to one drug of the FQs, it may also be resistant to other drugs of the FQs.
In this study, both mexA and mexE genes were not highly expressed, that is, the FQ resistance to P. aeruginosa in these resistant mutants cannot be contributed to efflux pumps. This may be because multidrug resistance was correlated with the overexpression of efflux pumps,Citation32–Citation34 while all resistant mutants in this study were not multidrug-resistant strains. So, in this study, another molecular resistance mechanism, mutations in target genes, mainly contributed to the FQ resistance to P. aeruginosa. Our results showed all mutations were detected in FQ-resistant strains and all point mutations were detected in gyrA and gyrB. In contrast, no mutations were found in the susceptible strains. These results indicated that mutations in gyrA and gyrB are associated with the resistance of P. aeruginosa to FQs. Meanwhile, our results found no mutations were detected in parC, which was not consistent with some studies.Citation7,Citation9,Citation35 This may be because P. aeruginosa is a gram-negative bacterium and the first target of FQ against gram-negative bacteria is DNA gyrase encoded by gyrA and gyrB.Citation8,Citation10,Citation11
Previous studies have reported that major amino acid alterations in gyrA frequently occur at codon 83 (Thr83→Ile), and gyrA mutations were closely correlated with FQ resistance to P. aeruginosa.Citation7,Citation9,Citation15,Citation16,Citation17,Citation36 A study had reported that amino acid alterations in gyrB occurred at codon 467 (Ser467→Phe) and 468 (Gln468→His), but the relationship between the alterations and resistance was not established.Citation15 These were not completely consistent with our results. Mutations in gyrA at codon 83(Thr83→Ile) were also found in our study, but our results showed four amino acid alterations in gyrB were mainly detected, including Ala372→Val, Ile424→Leu, Leu464→Ile and Glu483→Asp. Also, our results revealed that mutations in gyrB played an important role in FQ resistance to P. aeruginosa. But as the mutations in gyrB in FQ-resistant P. aeruginosa were reported rarely, we have no more data to compare. Also, the relationship between gyrB mutations and resistance needs further experiments to be confirmed.
In this study, resistant mutants with gyrA had the same amino acid alteration under the subconcentration of 0.5× MIC, whereas under the higher concentrations of 2× MIC and 4× MIC, they had other novel amino acid alterations. For resistant mutants with gyrB, the same change was found under lower and higher concentrations. These results indicated that resistant mutants may obtain stable drug resistance under the subconcentration, whereas higher incubation concentrations make mutations in genes of gyrA and gyrB changeable. This needs further experiments to be confirmed. Some studies reported that resistant strains can be cultured continuously in a medium without antimicrobial drugs to evaluate the stability of resistance.Citation37 In addition, these minor and rare mutations in gyrA and gyrB under higher concentrations may increase the likelihood of resistance or make the strains high-level resistant, which needs further verification.
Conclusion
In this study, CIP appeared to have stronger ability than LEV to make the strains resistant to in vitro selection. The molecular resistance mechanisms of FQs to P. aeruginosa were mainly the mutations in gyrA and gyrB, and the mutations in gyrB played a more important role in drug resistance.
Acknowledgments
We are grateful to the First Hospital of Wuhan for providing us with PAO1 of Pseudomonas aeruginosa. We would like to acknowledge to the Second Hospital of Shanxi Medical University for supporting this research. This work was supported by the Shanxi Province Natural Science Foundation (grant number 201601D11113). The sponsor had no involvement in any of the stages from the study design to submission of the paper for publication.
Disclosure
The authors report no conflicts of interest in this work.
References
- WeinerLMWebbAKLimbagoBAntimicrobial-resistant pathogens associated with healthcare-associated infections: summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2011–2014Infect Control Hosp Epidemiol201637111288130127573805
- BassettiMVenaACroxattoARighiEGueryBHow to manage infectionsDrugs Context2018721252729872449
- NguyenLGarciaJGruenbergKMacdougallCMultidrug-resistant Pseudomonas infections: hard to treat, but hope on the horizon?Curr Infect Dis Rep20182082329876674
- ShariatiAAzimiTArdebiliAInsertional inactivation of oprD in carbapenem-resistant Pseudomonas aeruginosa strains isolated from burn patients in Tehran, IranNew Microbes New Infect201821758029234497
- QiZDuanMLiAResearch status of drug resistance mechanism of Pseudomonas aeruginosaShandong Med J20145448386
- MoradaliMFGhodsSRehmBHPseudomonas aeruginosa lifestyle: a paradigm for adaptation, survival, and persistenceFront Cell Infect Microbiol201773928261568
- NguyenKNguyenTNguyenHLeDMutations in the gyrA, parC, and mexR genes provide functional insights into the fluoroquinolone-resistant isolated in VietnamInfect Drug Resist20181127528229535543
- JedreyHLilleyKSWelchMCiprofloxacin binding to GyrA causes global changes in the proteome of Pseudomonas aeruginosaFEMS Microbiol Lett201836513
- NouriRAhangarzadeh RezaeeMHasaniAAghazadehMAsgharzadehMThe role of gyrA and parC mutations in fluoroquinolones-resistant Pseudomonas aeruginosa isolates from IranBraz J Microbiol201647492593027522930
- GorganiNAhlbrandSPattersonAPourmandNDetection of point mutations associated with antibiotic resistance in Pseudomonas aeruginosaInt J Antimicrob Agents200934541441819656662
- DrlicaKMalikMKernsRJZhaoXQuinolone-mediated bacterial deathAntimicrob Agents Chemother200852238539217724149
- HuFGuoYZhuDSurveillance of CHINET bacterial resistance in ChinaChin J Infect Chemother2017175481491
- Ben NejmaMSioudOMastouriMQuinolone-resistant clinical strains of Pseudomonas aeruginosa isolated from University Hospital in Tunisia3 Biotech2018811
- VingopoulouEIDelisGABatziasGCPrevalence and mechanisms of resistance to fluoroquinolones in Pseudomonas aeruginosa and Escherichia coli isolates recovered from dogs suffering from otitis in GreeceVet Microbiol201821310210729291992
- YangXXingBLiangCYeZZhangYPrevalence and fluoroquinolone resistance of Pseudomonas aeruginosa in a hospital of South ChinaInt J Clin Exp Med2015811386139025785142
- SalmaRDabboussiFKassaaIKhudaryRHamzeMgyrA and parC mutations in quinolone-resistant clinical isolates of Pseudomonas aeruginosa from Nini Hospital in north LebanonJ Infect Chemother2013191778122821356
- MatsumotoMShigemuraKShirakawaTMutations in the gyrA and parC genes and in vitro activities of fluoroquinolones in 114 clinical isolates of Pseudomonas aeruginosa derived from urinary tract infections and their rapid detection by denaturing high-performance liquid chromatographyInt J Antimicrob Agents201240544044422884856
- FengYJonkerMJMoustakasIBrulSTer KuileBHDynamics of mutations during development of resistance by Pseudomonas aeruginosa against five antibioticsAntimicrob Agents Chemother20166074229423627139485
- AraisLRBarbosaAVCarvalhoCACerqueiraAMAntimicrobial resistance, integron carriage, and gyrA and gyrB mutations in Pseudomonas aeruginosa isolated from dogs with otitis externa and pyoderma in BrazilVet Dermatol2016272113e3126833540
- de La FuenteCMDaurosSPBelloTHMutations in gyrA and gyrB genes among strains of Gram-negative bacilli isolated from Chilean hospitals and their relation with resistance to fluoroquinolonesRev Med Chil200713591103111018064363
- CockerillFRPerformance Standards for Antimicrobial Susceptibility Testing: Twenty-First Informational SupplementWayne, PAClinical and Laboratory Standards Institute2017
- GaoYDuanJGengXDeficiency of quorum sensing system inhibits the resistance selection of Pseudomonas aeruginosa to ciprofloxacin and levofloxacin in vitroJ Glob Antimicrob Resist20171011311928729210
- BruchmannSDötschANouriBChabernyIFHäusslerSQuantitative contributions of target alteration and decreased drug accumulation to Pseudomonas aeruginosa fluoroquinolone resistanceAntimicrob Agents Chemother20135731361136823274661
- LiJXieSAhmedSAntimicrobial activity and resistance: influencing factorsFront Pharmacol2017836428659799
- AllenGPHarrisKAIn vitro resistance selection in Shigella flexneri by azithromycin, ceftriaxone, ciprofloxacin, levofloxacin, and moxifloxacinAntimicrob Agents Chemother2017617
- DayTHuijbenSReadAFIs selection relevant in the evolutionary emergence of drug resistance?Trends Microbiol201523312613325680587
- CantónRMorosiniMIEmergence and spread of antibiotic resistance following exposure to antibioticsFEMS Microbiol Rev201135597799121722146
- HansenGTZhaoXDrlicaKBlondeauJMMutant prevention concentration for ciprofloxacin and levofloxacin with Pseudomonas aeruginosaInt J Antimicrob Agents200627212012416426820
- Shun-MeiEZengJMYuanHLuYCaiRXChenCSub-inhibitory concentrations of fluoroquinolones increase conjugation frequencyMicrob Pathog2018114576229174700
- RenLHuangZLiwangYCDrug resistance and cross resistance of mutidrug resistant Mycobacterium tuberculosis to fluoroquinolones in vitroChina Pharm20151114881490
- XuchengShenXLiuMLiJDaiYIn vitro induction of resistance and cross resistance to five quinolones in Salmonella pullorumChina Poult20173992023
- LópezCATraversTPosKMZgurskayaHIGnanakaranSDynamics of intact MexAB–OprM efflux pump: focusing on the MexA–OprM interfaceSci Rep2017711652129184094
- HelmyOMKashefMTDifferent phenotypic and molecular mechanisms associated with multidrug resistance in Gram-negative clinical isolates from EgyptInfect Drug Resist20171047949829263684
- AdabiMTalebi-TaherMArbabiLSpread of efflux pump overexpressing-mediated fluoroquinolone resistance and multidrug resistance in Pseudomonas aeruginosa by using an efflux pump inhibitorInfect Chemother20154729810426157587
- YangWZhangMZhouJPangLWangGHouFThe molecular mechanisms of ciprofloxacin resistance in clinical Campylobacter jejuni and their genotyping characteristics in Beijing, ChinaFoodborne Pathog Dis201714738639228696791
- MuruganNMalathiJThereseKLMadhavanHNApplication of six multiplex PCR’s among 200 clinical isolates of Pseudomonas aeruginosa for the detection of 20 drug resistance encoding genesKaohsiung J Med Sci2018342798829413231
- HuMWangBFuLInduction and stability of Mycobacterium tuberculosis resistance to PA-824 in vitroChin J Antibiot2017422108112