
Abstract
Background
Colistin acts as the last line of defense against severe infections caused by carbapenem-resistant Enterobacteriaceae. Infections caused by extensively drug-resistant isolates coproducing MCR and carbapenemases have posed a serious public health concern.
Purpose
In this study, we reported the first clinical colistin and carbapenem-resistant Enterobacter hormaechei isolate SCNJ07 coharboring blaNDM-1 and mcr-9 from a patient with bloodstream infection in China.
Methods
Bacterial antimicrobial susceptibility testing was performed using the broth microdilution method. Conjugation assay was carried out to investigate the transferability of mcr-9 and blaNDM-1. Whole-genome sequencing of strain SCNJ07 was performed using an Illumina HiSeq system and the genetic characteristics of the mcr-9- and blaNDM-1-harboring plasmids were analyzed.
Results
Conjugation assays revealed that both blaNDM-1 and mcr-9 genes could successfully transfer their resistance phenotype to Escherichia coli strain J53. Whole genome sequencing showed that SCNJ07 possessed an FIB36:FIIY4 type self-transmissible plasmid bearing blaNDM-1, which possessed high similarity to previously reported pRJF866 in China. mcr-9 was located on a ~28-kb self-transmissible plasmid pMCR-SCNJ07 with both IncHI2 and IncR replicons. Two copies of intact IS903 that bracketed a ~8-kb region containing the mcr-9 gene were identified in pMCR-SCNJ07. BLASTn analysis revealed that a number of mcr-9-positive plasmids have been around for a while among Enterobacteriaceae worldwide.
Conclusion
This study reveals the likelihood of a wide dissemination of this newly identified colistin resistance gene mcr-9 among Enterobacteriaceae. Further surveillance is urgently needed to understand the prevalence and dissemination of mcr-9, thereby facilitating establishment of measures to control its spread.
Introduction
Carbapenem-resistant Enterobacteriaceae (CRE)-related infections are global public health issues.Citation1 The New Delhi metallo-β-lactamase (NDM) is one of the most common carbapenemases worldwide.Citation2 Polymyxins are among the last-line therapeutic options to treat serious infections caused by CRE.Citation3 However, concerns were raised regarding the increasing prevalence of the plasmid-borne colistin resistance gene, mcr-1, which has been detected from the environment, food, animals and humans around the world since its first discovery in China in late 2015.Citation4–Citation6 More worrisomely, cases of infection caused by extensively drug-resistant isolates that coproduce MCR-1 and NDM have been reported by groups from different regions.Citation7–Citation13
Following a recent study which identified an additional mcr homolog, mcr-9, in a Salmonella enterica serotype Typhimurium isolate,Citation14 we isolated an extensively drug-resistant Enterobacter hormaechei strain, which coproduces NDM-1 and MCR-9, from a patient with bloodstream infection. The genetic characteristics of the blaNDM-1 and mcr-9-harboring plasmids were analyzed. In addition, comparative genetic analysis of mcr-9 in pMCR-SCNJ07 and closely related plasmids were also carried out. To the best of our knowledge, this is the first report describing a clinical colistin and carbapenem-resistant E. hormaechei isolate coharboring blaNDM-1 and mcr-9 in China.
Materials And Methods
Bacterial Isolation And Identification
The E. hormaechei strain SCNJ07 was recovered from the blood sample of a 50-year-old male patient with obstructive jaundice in a hospital in Sichuan, China, in September 2018. It was initially identified as Enterobacter cloacae using the Vitek-2 compact system (bioMérieux, Marcy-l’Étoile, France) and by species identification established by sequencing of the 16S rRNA gene amplified with the universal primers 27F and 1492R.Citation15 The presence of the acquired carbapenemase genes (blaKPC, blaNDM, blaGES, blaIMP, blaOXA-48, and blaVIM) and mcr genes (mcr-1 to mcr-8) in SCNJ07 was determined by PCR amplification as described previously.Citation16–Citation21 The detection of mcr-9 was amplified using the primers MCR9-F (5ʹ- CTTTCCATAACAGCGAGACAC-3ʹ) and MCR9-R (5ʹ- GTATCCTTCCTGCCATCCTC-3ʹ).
Antimicrobial Susceptibility Testing
In vitro susceptibility tests of ceftazidime, ceftriaxone, cefepime, cefazolin, cefotetan, imipenem, aztreonam, ciprofloxacin, gentamycin, tobramycin, levofloxacin, amikacin, piperacillin-tazobactam and trimethoprim-sulfamethoxazole were performed using Vitek-2 system. The minimum inhibitory concentrations (MICs) of meropenem, colistin, polymyxin B, doxycycline, fosfomycin and tigecycline against the strain were determined using the microdilution broth method following recommendations of the Clinical Laboratory Standards Institute (CLSI) (CLSI, 2019). Breakpoints of colistin and tigecycline were interpreted according to the European Committee on Antimicrobial Susceptibility Testing (EUCAST) (http://www.eucast.org/); otherwise, we applied those defined by the CLSI.
Conjugation Assay
Conjugation experiments were carried out using broth-based methods with Escherichia coli J53 (azide-resistant) as the recipient strain. Transconjugants were selected on LB agar plates containing 150 μg/mL sodium azide plus 1 μg/mL meropenem or 4 μg/mL colistin. The presence of blaNDM-1 or mcr-9 in transconjugants was confirmed by PCR and Sanger sequencing.
Genome Sequencing And Analysis
Total genomic DNA of E. coli isolates was extracted using Rapid Bacterial Genomic DNA Isolation Kit (Sangon Biotech, Shanghai, China) according to the manufacturer’s protocol. Purified DNA was subjected to whole genomic sequencing on the Illumina HiSeq 2000 system with the 150-bp paired-end approach and 150× coverage. Reads were trimmed using TrimmomaticCitation22 and were then assembled using the SOAP de novo program.Citation23 Annotation was carried out using Prokka.Citation24 The species identification was performed by average nucleotide identity (ANI) analysis with JSpeciesWS (http://jspecies.ribohost.com/jspeciesws/#analyse). Antimicrobial resistance genes were identified using ResFinder v3.1 of the Center for genomic epidemiology (http://genomicepidemiology.org/).
Plasmid Sequencing And Analysis
Plasmid DNA from transconjugants containing blaNDM-1 or mcr-9 was extracted and sequenced using the Illumina HiSeq system. After filtering J53 chromosomal DNA data and assembling the remaining reads, the plasmid carrying blaNDM-1 or mcr-9 was completely circularized using PCR and Sanger sequencing to fill in gaps between contigs. The plasmid replicon type and MLST were determined using the PlasmidFinder (https://cge.cbs.dtu.dk/services/PlasmidFinder/) and pMLST tools (https://cge.cbs.dtu.dk/services/pMLST/). The annotations of the plasmid sequences were conducted using the RAST tools and edited manually.Citation25 Sequence alignment of mcr-9-carrying plasmids was performed using BLAST and visualized with Easyfig v 2.2.3.Citation26 Alignments with highly homologous complete plasmid sequences of pMCR-SCNJ07 available in NCBI were performed by using the BRIG tool.Citation27
Nucleotide Sequence Accession Numbers
Draft genome sequence of the strain SCNJ07 has been deposited into GenBank under the accession no. VBSC00000000. The complete sequences of pNDM-SCNJ07 and pMCR-SCNJ07 have been deposited into GenBank under accession no. MK933278 and MK933279, respectively.
Ethics Statement
The clinical isolate E. hormaechei SCNJ07 was generated as part of the routine hospital laboratory procedure. The Ethics Committee of the Southwest Medical University exempted this study from review because the present study only focused on bacteria and patient intervention was not required.
Results And Discussion
Antimicrobial Susceptibility Of The Strain E. hormaechei SCNJ07
Susceptibility testing by Vitek-2 system showed that the strain SCNJ07 was resistant to all tested drugs, including ceftazidime (MIC, ≥64 µg/mL), ceftriaxone (MIC, ≥64 µg/mL), cefepime (MIC, ≥64 µg/mL), cefazolin (MIC, ≥64 µg/mL), cefotetan (MIC, ≥64 µg/mL), imipenem (MIC, ≥16 µg/mL), aztreonam (MIC, ≥64 µg/mL), ciprofloxacin (MIC, ≥4 µg/mL), gentamycin (MIC, ≥16 μg/mL), tobramycin (MIC, ≥16 μg/mL), levofloxacin (MIC, ≥8 μg/mL), amikacin (MIC, ≥64 μg/mL), piperacillin-tazobactam (MIC, ≥128 µg/mL) and trimethoprim-sulfamethoxazole (MIC, ≥320 µg/mL). Additional broth microdilution testingCitation28 showed that it also exhibited resistance to meropenem (MIC, 256 µg/mL), doxycycline (MIC, 128 µg/mL), colistin (MIC, 16 µg/mL) and polymyxin B (MIC, 8 µg/mL) but was susceptible to fosfomycin (MIC, 64 µg/mL) and tigecycline (MIC, 2 µg/mL). PCR and sequencing analysis showed that the strain SCNJ07 harbored both a blaNDM-1 carbapenemase gene and the recently identified plasmid-borne colistin resistance gene mcr-9.
Location And Transferability Of mcr-9 And blaNDM-1
Conjugation assays showed that blaNDM-1 and mcr-9 were successfully transferred to E. coli J53 at the frequency of ~10−2 and 10−4 (transconjugant/recipient), respectively. Compared with the MICs for the recipient strain J53, transformants containing blaNDM-1 showed a 128-fold increase for meropenem (from 0.5 to 64 μg/mL) and those harboring mcr-9 16-fold for colistin (from 0.5 to 8 μg/mL). These findings revealed that blaNDM-1 and mcr-9 were functional and carried by two self-transmissible plasmids.
Genome Characteristics Of The Strain SCNJ07
Draft genome sequence of SCNJ07 was assembled into 120 contigs (112 were >1000 bp in length), which comprises 5,321,397 bp, with a 54.77% GC content. Species identification based on ANI analysis confirmed that the strain SCNJ07 actually belongs to E. hormaechei, as it only had an 86.32% identity (75.45% query coverage) to E. cloacae subsp. cloacae ATCC 13,047, but a 98.41% identity (80.30% coverage) to E. hormaechei subsp. steigerwaltii strain DSM 16691, obviously above the 95–96% cut-off for defining a bacterial species.Citation29
Resistance Profile Of The Strain SCNJ07
Analysis of the whole genome sequence of SCNJ07 allowed us to more confidently conclude that blaNDM-1 and mcr-9 co-existed in this strain. Besides, in consistence with its multidrug resistance phenotype, SCNJ07 also had multiple genes mediating resistance to β-lactams (blaCTX-M-3, blaTEM-1B, blaSHV-12 and blaACT-7), aminoglycosides (aac(6ʹ)-IIa, aadA16, aac(6ʹ)-Ib3 and rmtC), fluoroquinolones (qnrS1 and aac(6ʹ)-Ib-cr), macrolide (mph(A)), rifampicin (ARR-3), tetracycline (tet(D)), sulfonamide (sul1) and trimethoprim (dfrA16). E. hormaechei, a common nosocomial pathogen of clinical significance, was reported in several outbreaks of sepsis in neonatal intensive care units in the USA and in Brazil,Citation30 while it was only sporadically reported in China.Citation31 The identification of E. hormaechei SCNJ07 coproducing NDM-1 and MCR-9 in this study highlights the need to enhance the epidemiologic surveillance of this novel colistin resistance gene in CRE.
Analysis Of The blaNDM-1-Harbouring Plasmid pNDM-SCNJ07
Plasmid analysis revealed that the blaNDM-1 was carried by an FIB36:FIIY4 replication type plasmid, designated pNDM-SCNJ07, which was 110,786 bp in length and had an average GC content of 54.84%. pNDM-SCNJ07 is almost identical (99% identity and 99.98% coverage) to the plasmid pRJF866 (GenBank accession no. KF732966) from K. pneumoniae from Shanghai, China, in 2015.Citation32 Besides, similar FIB36:FIIY4-type plasmids carrying blaNDM-1 have been widely found among Enterobacteriaceae, including pKOX_NDM1 (Accession no. JQ314407) from Klebsiella michiganensis from Taiwan,Citation33 pNDM_20ES (Accession no. MF42356) from E. cloacae and pNDM_4TM (Accession no. MF42352) from Serratia marcescens from Romania.Citation34 On these plasmids, the blaNDM-1 gene was embedded in the same genetic environment and its flanking miniature inverted-repeat transposable elements (MITEs, positions 70834 to 71089 and 77035 to 77290 of pNDM-SCNJ07) were suggested to be responsible for the mobilization of blaNDM-1 onto the FIB36:FIIY4 plasmids.Citation33,Citation35
Genetic Characteristics Of The Plasmid-borne mcr-9
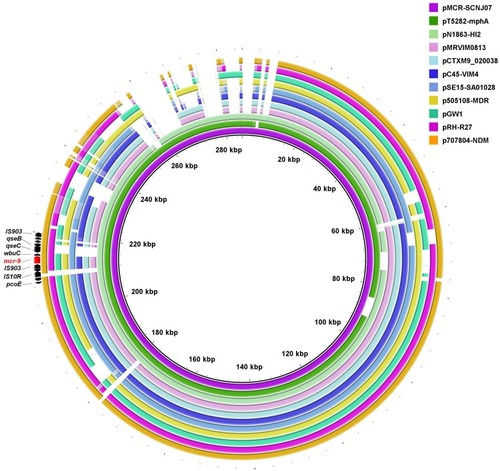
Complete sequence of the mcr-9-carrying plasmid, designated pMCR-SCNJ07, was circularized by PCR mapping using pT5282-mphA (Accession no. KY270852) as the template. pMCR-SCNJ07 was 285,587 bp in size with an average G+C content of 47.31%. The plasmid contained 327 predicted ORFs, and two replicons, IncHI2 (ST1) and IncR, and carried a number of additional resistance genes, including blaSHV-12, aadA16, aac(6ʹ)-IIa, mph(A), sul1 and tet(D). Sequence alignment showed that pMCR-SCNJ07 displayed 97% query coverage and 99.99% identity with the reference plasmid pT5282-mphA from an E. cloacae isolated in 2012 from a teaching hospital in Chongqing, China.Citation36 By BLAST, pMCR-SCNJ07 was also similar to several previously sequenced IncHI2 plasmids (), including pMRVIM0813 (86% query coverage and 99.97% identity, accession no. KP975077), pCTXM9_020038 (85% query coverage and 99.97% identity, accession no. CP031724), pC45-VIM4 (85% query coverage and 99.96% identity, accession no. LT991958) and pSE15-SA01028 (83% query coverage and 99.99% identity, accession no. CP026661), from a clinical E. cloacae isolate from USA in 2015, an E. hormaechei from China in 2016, an E. cloacae from France in 2018 and a Salmonella enterica subsp. Enterica isolate from Germany in 2015, respectively. It is noteworthy that all these plasmids were mcr-9-positive, highlighting an earlier presence of mcr-9 among Enterobacteriaceae around the world than previously known and raising the likelihood of ongoing undetected transmission. Close surveillance is urgently needed to determine mcr-9 prevalence and effective actions are required to control its further dissemination.
Figure 1 Circular comparison of mcr-9-carrying IncHI2 plasmids. The complete sequence of pMCR-SCNJ07 was used as the reference. The arrows indicate deduced ORFs and their orientations. The mcr-9 gene is indicated in red. The circular maps were generated using BRIGCitation27 and plasmids were included in the following order (inner to outer circles): pMCR-SCNJ07 (this study, accession no. MK933279), pT5282-mphA (KY270852), pN1863-HI2 (MF344583), pMRVIM0813 (KP975077), pCTXM9_020038 (CP031724), pC45-VIM4 (LT991958), pSE15-SA01028 (CP026661), p505108-MDR (KY978628), pGW1 (CP028975), pRH-R27 (LN555650) and p707804-NDM (MH909331).
The genetic environment of mcr-9 has not been described before. On pMCR-SCNJ07, mcr-9 was located in a ~8kb region bracketed by two copies of intact IS903, an insertion sequence originating from E. coli. In this region, wbuC (encoding a cupin fold metalloprotein) was located downstream of mcr-9, followed by a two-component system encoding genes qseC and qseB, a truncated insertion sequence ∆IS1R, and a remnant of ∆silR (encoding a transcriptional regulatory protein) (). The two copies of intact IS903 could form a composite transposon or a circular intermediate, which has the potential to mobilize the mcr-9 gene. However, no circular intermediate was detected despite repeated attempts via inverse PCR in our work. Whether mcr-9 could transfer through the formation of a circular intermediate requires further investigation. Of note, similar to the scenario in all known mcr-1 cassettes, in which one or both ISApl1 sequences are consistently absent in the mcr-1-bearing region,Citation37 the downstream IS903 was absent on some mcr-9-carrying plasmids ().
Figure 2 Colinear analyses for genetic environment of mcr-9. (A) Comparison of the mcr-9-containing regions from different plasmid reservoirs and the chromosome of E. hormaechei strain S5. (B) Comparison of the mcr-9-neighboring genetic contexts on IncHI2-type plasmids. The corresponding region on non-mcr-9-carrying plasmid R478 (top) is shown for comparison. Arrows indicate the positions and directions of the genes and Δ indicates the truncated gene. mcr-9 genes are indicated in dark blue and IS903 are highlighted in purple. Gray shades denote shared regions with a high degree of homology. Vertical green bars represent the direct repeats of IS10R. The accession numbers were: E. hormaechei strain S5 (Accession no. CP031571), pT5282-mphA (KY270852), pN1863-HI2 (MF344583), pGW1 (CP028975), R478 (NC_005211), pRH-R27 (LN555650), pN1863-HI2 (MF344583), and pMCR-SCNJ07 (this study, MK933279). The construction of sequence comparison was performed using BLAST (http://blast.ncbi.nlm.nih.gov) and Easyfig version 2.2.3.Citation26
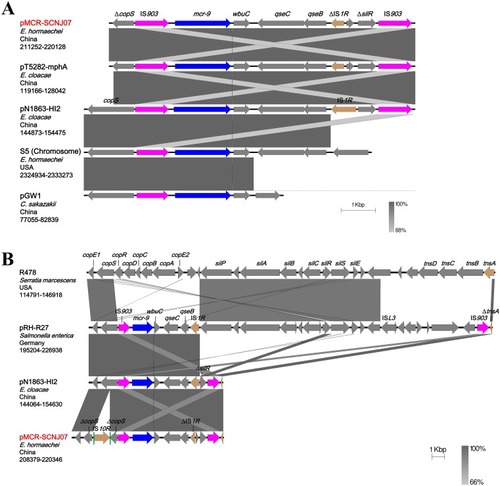
A pairwise comparison of the sil–cop region of IncHI2 plasmids R478 and pRH-R27 enabled us to learn that mcr-9 was likely to be transferred by the IS1R to IS903 region initially, and that insertion of this mcr-9-carrying region between sil and cop resulted in truncation of cop resistance gene clusters into ∆copS–copE1 ().Citation38 BLASTn analysis suggested that the mcr-9-carrying region of pMCR-SCNJ07 could be derived from pRH-R27 as a result of multiple genetic events (). No 9-bp target site duplication repeats, which are characteristic of the insertion of IS903, could be identified in the sequence adjacent to both IS903 elements. The detail of the evolutionary route of the mcr-9-carrying region, therefore, remains unclear.
To address the possible origin of mcr-9, a BLAST search against the NCBI’s non-redundant protein sequence database (nr) was conducted by using the amino acid sequence of mcr-9 as a probe. The result identified hits aligned to phosphoethanolamine transferases from multiple genera of Enterobacteriaceae (100% query coverage and 94–100% identity), followed by that from Buttiauxella brennerae (WP_064558897, 100% query coverage and 86.83% identity). It should be noted that of the flanking genes of mcr-9 within the ~8kb region, none but the wbuC gene showed homologous to that from Buttiauxella, with 98% query coverage and 85.9% identity between their amino acid sequences. These findings suggested that the disseminated mcr-9 gene in Enterobacteriaceae might have originated from Buttiauxella species and that the wbuC gene had been likely comobilized with mcr-9 from its original genetic context. Additional studies are needed to fully characterize the origin of mcr-9.
Conclusion
In summary, we here report the discovery of a clinical E. hormaechei strain carrying both blaNDM-1 and mcr-9. Despite few reports characterizing mcr-9-harboring plasmids, this newly identified mobile colistin resistance gene is likely to have already been widely disseminated. Yet more worryingly, two copies of IS903 encompassing the mcr-9 gene have the potential to mobilize mcr-9 by hijacking more plasmids as the vehicle to disseminate this gene. Therefore, screening for the mcr-9 gene should be urgently included in the surveillance of colistin-resistant Enterobacteriaceae from humans, animals, and the environment.
Author Contributions
LZ designed the experiments. YY, GW, and CL performed the experiments. LX, JS and YY analyzed the data. YL and LZ analyzed the data and wrote the manuscript. FZ edited the original draft. All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Acknowledgments
This work was supported by the Project of Education Department in Sichuan, China (18ZB0633), Natural Science Foundation of Southwest Medical University (No. 2017-ZRZD-022 and 2018-ZRZD-011), and National Undergraduate Innovation and Entrepreneurship Project (No.201816032021). The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Disclosure
The authors report no conflicts of interest in this work.
References
- Logan LK, Weinstein RA. The epidemiology of carbapenem-resistant Enterobacteriaceae: the impact and evolution of a global menace. J Infect Dis. 2017;215(suppl_1):S28–S36. doi:10.1093/infdis/jiw28228375512
- Wu WJ, Feng Y, Tang GM, Qiao F, McNally A, Zong ZY. NDM metallo-β-lactamases and their bacterial producers in health care settings. Clin Microbiol Rev. 2019;32(2):e00115–00118. doi:10.1128/CMR.00115-1830700432
- Kaye KS, Pogue JM, Tran TB, Nation RL, Li J. Agents of last resort: polymyxin resistance. Infect Dis Clin North Am. 2016;30(2):391–414. doi:10.1016/j.idc.2016.02.00527208765
- Liu YY, Wang Y, Walsh TR, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16(2):161–168. doi:10.1016/S1473-3099(15)00424-726603172
- Sun J, Zhang HM, Liu YH, Feng YJ. Towards understanding MCR-like colistin resistance. Trends Microbiol. 2018;26(9):794–808. doi:10.1016/j.tim.2018.02.00629525421
- Wang Q, Sun J, Li J, et al. Expanding landscapes of the diversified mcr-1-bearing plasmid reservoirs. Microbiome. 2017;5(1):70. doi:10.1186/s40168-017-0288-028683827
- Delgado-Blas JF, Ovejero CM, Abadia-Patino L, Gonzalez-Zorn B. Coexistence of mcr-1 and blaNDM-1 in Escherichia coli from Venezuela. Antimicrob Agents Chemother. 2016;60(10):6356–6358. doi:10.1128/AAC.01319-1627431212
- Liu BT, Song FJ, Zou M, Hao ZH, Shan H. Emergence of colistin resistance gene mcr-1 in Cronobacter sakazakii producing NDM-9 and in Escherichia coli from the same animal. Antimicrob Agents Chemother. 2017;61(2):e01444–16.27855074
- Feng S, Shen C, Chen H, et al. Co-production of MCR-1 and NDM-5 in Escherichia coli isolated from a colonization case of inpatient. Infect Drug Resist. 2018;11:1157–1161. doi:10.2147/IDR.S17116430147343
- Li X, Mu X, Zhang P, et al. Detection and characterization of a clinical Escherichia coli ST3204 strain coproducing NDM-16 and MCR-1. Infect Drug Resist. 2018;11:1189–1195. doi:10.2147/IDR.S17504130147347
- Xu L, Wang P, Cheng J, Qin S, Xie W. Characterization of a novel bla NDM-5-harboring IncFII plasmid and an mcr-1-bearing IncI2 plasmid in a single Escherichia coli ST167 clinical isolate. Infect Drug Resist. 2019;12:511–519. doi:10.2147/IDR.S19299830881056
- Lin YC, Kuroda M, Suzuki S, Mu JJ. Emergence of an Escherichia coli strain co-harbouring mcr-1 and blaNDM-9 from a urinary tract infection in Taiwan. J Glob Antimicrob Resist. 2019;16:286–290. doi:10.1016/j.jgar.2018.10.00330312830
- Mediavilla JR, Patrawalla A, Chen L, et al. Colistin- and carbapenem-resistant Escherichia coli harboring mcr-1 and blaNDM-5, causing a complicated urinary tract infection in a patient from the United States. MBio. 2016;7(4). doi:10.1128/mBio.01191-16.
- Carroll LM, Gaballa A, Guldimann C, Sullivan G, Henderson LO, Wiedmann M. Identification of novel mobilized colistin resistance gene mcr-9 in a multidrug-resistant, colistin-susceptible Salmonella enterica serotype Typhimurium isolate using a combination of high-throughput, in silico screening and functional analysis. mBio. 2019;10(3): e00853-19. doi:10.1128/mBio.00853-19
- Lane DJ. 16S/23S rRNA sequencing. Nucleic Acid Tech Bacterial Sys. 1991;115–175.
- Szekely E, Damjanova I, Janvari L, et al. First description of bla(NDM-1), bla(OXA-48), bla(OXA-181) producing Enterobacteriaceae strains in Romania. Int J Med Microbiol. 2013;303(8):697–700. doi:10.1016/j.ijmm.2013.10.00124183483
- Dallenne C, Da Costa A, Decre D, Favier C, Arlet G. Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in Enterobacteriaceae. J Antimicrob Chemother. 2010;65(3):490–495. doi:10.1093/jac/dkp49820071363
- Ellington MJ, Kistler J, Livermore DM, Woodford N. Multiplex PCR for rapid detection of genes encoding acquired metallo-beta-lactamases. J Antimicrob Chemother. 2007;59(2):321–322. doi:10.1093/jac/dkl48117185300
- Poirel L, Le Thomas I, Naas T, Karim A, Nordmann P. Biochemical sequence analyses of GES-1, a novel class A extended-spectrum beta-lactamase, and the class 1 integron In52 from Klebsiella Pneumoniae. Antimicrob Agents Chemother. 2000;44(3):622–632. doi:10.1128/AAC.44.3.622-632.200010681329
- Wang X, Wang Y, Zhou Y, et al. Emergence of a novel mobile colistin resistance gene, mcr-8, in NDM-producing Klebsiella pneumoniae. Emerg Microbes Infect. 2018;7(1):122. doi:10.1038/s41426-018-0124-z29970891
- Yang YQ, Li YX, Lei CW, Zhang AY, Wang HN. Novel plasmid-mediated colistin resistance gene mcr-7.1 in Klebsiella pneumoniae. J Antimicrob Chemother. 2018;73:1791–1795. doi:10.1093/jac/dky11129912417
- Bolger AM, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics. 2014;30(15):2114–2120. doi:10.1093/bioinformatics/btu17024695404
- Li R, Li Y, Kristiansen K, Wang J. SOAP: short oligonucleotide alignment program. Bioinformatics. 2008;24(5):713–714. doi:10.1093/bioinformatics/btn02518227114
- Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014;30(14):2068–2069. doi:10.1093/bioinformatics/btu15324642063
- Aziz RK, Bartels D, Best AA, et al. The RAST server: rapid annotations using subsystems technology. BMC Genomics. 2008;9:75. doi:10.1186/1471-2164-9-7518261238
- Sullivan MJ, Petty NK, Beatson SA. Easyfig: a genome comparison visualizer. Bioinformatics. 2011;27(7):1009–1010. doi:10.1093/bioinformatics/btr03921278367
- Alikhan NF, Petty NK, Ben Zakour NL, Beatson SA. BLAST ring image generator (BRIG): simple prokaryote genome comparisons. BMC Genomics. 2011;12:402. doi:10.1186/1471-2164-12-40221824423
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing. 29th CLSI supplement M100 Wayne, PA: Clinical and Laboratory Standards Institute; 2019
- Richter M, Rossello-Mora R. Shifting the genomic gold standard for the prokaryotic species definition. Proc Natl Acad Sci U S A. 2009;106(45):19126–19131. doi:10.1073/pnas.090641210619855009
- Mezzatesta ML, Gona F, Stefani S. Enterobacter cloacae complex: clinical impact and emerging antibiotic resistance. Future Microbiol. 2012;7(7):887–902. doi:10.2217/fmb.12.6122827309
- Zheng F, Sun J, Cheng C, Rui Y. Molecular characteristics of carbapenem-resistant gram-negative bacteria in southern China. Microb Drug Resist. 2015;21(2):178–185. doi:10.1089/mdr.2014.008525469995
- Qu H, Wang X, Ni Y, et al. NDM-1-producing Enterobacteriaceae in a teaching hospital in Shanghai, China: incX3-type plasmids may contribute to the dissemination of blaNDM-1. Int J Infect Dis. 2015;34:8–13. doi:10.1016/j.ijid.2015.02.02025743762
- Huang TW, Wang JT, Lauderdale TL, et al. Complete sequences of two plasmids in a blaNDM-1-positive Klebsiella oxytoca isolate from Taiwan. Antimicrob Agents Chemother. 2013;57(8):4072–4076. doi:10.1128/AAC.02266-1223752513
- Phan HTT, Stoesser N, Maciuca IE, et al. Illumina short-read and MinION long-read WGS to characterize the molecular epidemiology of an NDM-1 Serratia marcescens outbreak in Romania. J Antimicrob Chemother. 2018;73(3):672–679. doi:10.1093/jac/dkx45629237003
- Wailan AM, Sidjabat HE, Yam WK, et al. Mechanisms involved in acquisition of blaNDM genes by IncA/C2 and IncFIIY plasmids. Antimicrob Agents Chemother. 2016;60(7):4082–4088. doi:10.1128/AAC.00368-1627114281
- Liang Q, Yin Z, Zhao Y, et al. Sequencing and comparative genomics analysis of the IncHI2 plasmids pT5282-mphA and p112298-catA and the IncHI5 plasmid pYNKP001-dfrA. Int J Antimicrob Agents. 2017;49(6):709–718. doi:10.1016/j.ijantimicag.2017.01.02128390961
- Li R, Xie M, Zhang J, et al. Genetic characterization of mcr-1-bearing plasmids to depict molecular mechanisms underlying dissemination of the colistin resistance determinant. J Antimicrob Chemother. 2017;72(2):393–401. doi:10.1093/jac/dkw41128073961
- Dolejska M, Villa L, Poirel L, Nordmann P, Carattoli A. Complete sequencing of an IncHI1 plasmid encoding the carbapenemase NDM-1, the ArmA 16S RNA methylase and a resistance-nodulation-cell division/multidrug efflux pump. J Antimicrob Chemother. 2013;68(1):34–39. doi:10.1093/jac/dks35722969080