
Abstract
Purpose
This study aimed to evaluate the molecular characteristics and prevalence of clinical carbapenem-resistant Klebsiella aerogenes (CRKA), collected during an outbreak in a Chinese tertiary hospital.
Methods
Antimicrobial susceptibility test, using 17 antibiotics, was performed on 14 CRKA isolates. The strains were examined for the presence of β-lactamase genes by PCR, and efflux pump phenotype was determined by efflux pump inhibition test. Presence of outer-membrane porins was examined. Clonal relatedness among the isolates was investigated by pulsed-field gel electrophoresis (PFGE). S1 nuclease-PFGE and plasmid incompatibility group analysis were performed to determine plasmids, and the genetic environment of blaKPC-2 was analyzed. Epidemiological data were collected via chart review.
Results
The 14 CRKA isolates were all resistant to carbapenems; five distinct groups (PFGE types A–E) were observed. All 14 isolates carried the blaKPC-2 gene. S1 nuclease-PFGE indicated the size of blaKPC-2-carrying plasmids to range from 20 kb to 200 kb, and the 14 plasmids belonged to various incompatibility groups. The most frequent genetic environment of blaKPC-2 was Tn1721- blaKPC-2-ΔTn3-IS26. PFGE type A group, including 11 KPC-2-producing clinical isolates, was primarily responsible for dissemination.
Conclusion
Our findings suggest both transposons and vertical transmission to contribute to the transformation of blaKPC-2. The results strongly suggest strict implementation of infection control of CRKA, in healthcare facilities.
Video abstract

Point your SmartPhone at the code above. If you have a QR code reader the video abstract will appear. Or use:
Introduction
Carbapenem is a broad-spectrum β-lactam antibiotic with high stability to β-lactamases mediated by plasmids and chromosomes. However, carbapenem-resistant Enterobacteriaceae (CRE) strains have recently been reported frequently worldwide. CRE isolates show multiple-drug resistance, and even pan-drug resistance, to existing antimicrobial agents, resulting in a serious clinical challenge, treatment failure, and high mortality rates.Citation1–Citation3 Klebsiella aerogenes, formerly known as Enterobacter aerogenes, is one of the major bacterial species responsible for nosocomial infections. The prevalence of carbapenem-resistant K. aerogenes (CRKA) has reached 21% in a Chinese hospital in ShanghaiCitation4 and 12.5% in the USA,Citation5 thus undoubtedly becoming a major public health concern.Citation6,Citation7 Reported mechanisms of resistance to carbapenemsCitation8 include production of carbapenemases, overproduction of β-lactamases, efflux pumps, porin deficiency, and a change in penicillin-binding proteins. The most common carbapenemases that are overproduced by these strains include Klebsiella pneumoniae carbapenemase (KPC) and metalloenzymes.Citation9–Citation11 KPC-type carbapenemases remain the major determinant of carbapenem resistance in K. pneumoniae, and in many other Enterobacteriaceae.Citation12 Plasmids harboring the blaKPC gene, in association with the mobile element Tn1721, are largely responsible for the effective spread of these genes among K. pneumoniae in People’s Republic of China, especially among the ST11 isolates, which is in contrast to their spread via Tn4401 transposons reported in other countries. However, little is known about the genetic environment of Enterobacteriaceae, besides K. pneumoniae, in People’s Republic of China.
In the present study, we evaluated the molecular characteristics and prevalence of clinical CRKA isolates in a tertiary teaching hospital in Shanghai, People’s Republic of China. These data should provide valuable insight into the development of strategies for treatment and control of the spread of CRKA isolates.
Materials And Methods
Clinical Isolates
All the 61 clinical K. aerogenes strains, which were isolated from January 2013 to December 2013 at Huashan Hospital (Fudan University, Shanghai, People’s Republic of China), were screened to identify carbapenem-resistant isolates based on disc diffusion method results (imipenem or meropenem inhibition zone diameters ≤19 mm, or ertapenem inhibition zone diameters ≤18 mm). All the isolates in this manuscript were part of the routine hospital laboratory procedure. They are obtained from our hospital bacteria bank. They were not specifically isolated for this research. Based on this screen, a total of 14 non-duplicate carbapenem-resistant isolates were detected, and species confirmation was performed by 16S RNA amplification.Citation13
Antimicrobial Susceptibility
Antimicrobial susceptibility test was performed with all 14 CRKA isolates using disc diffusion and broth microdilution methods based on Clinical and Laboratory Standards Institute (CLSI), 2019 guidelines.Citation14 The minimum inhibitory concentrations (MICs) of imipenem, meropenem, ertapenem, piperacillin-tazobactam, cefepime, cefotaxime, cefoxitin, ceftazidime, amikacin, ciprofloxacin, aztreonam, cefuroxime, levofloxacin, gentamicin, and fosfomycin were determined according to CLSI standards, whereas those of tigecycline and colistin were determined according to European Committee on Antimicrobial Susceptibility Testing guidelines. In addition, the MICs of imipenem were determined in the presence of 25 g mL−1 of the efflux pump inhibitor Phe-Arg β-naphthylamide dihydrochloride (PAβN) to investigate the role of efflux pump in the carbapenem resistance mechanism.Citation15
Pulsed-Field Gel Electrophoresis (PFGE)
PFGE of XbaI (Takara, Dalian, People’s Republic of China)-digested genomic DNA samples of K. aerogenes isolates was performed according to the manufacturer’s instructions. The DNA fragments were separated with a CHEF-Mapper XA PFGE system (Bio-Rad, USA). Electrophoresis was performed for 20 hrs at 14°C with pulse times ranging from 3 to 40 s at 6 V cm−1. The DNA fingerprints generated were analyzed according to the criteria proposed by Tenover et alCitation16.
Detection Of β-Lactamase Genes
All K. aerogenes isolates were screened for carbapenemase-encoding genes (blaSME, blaKPC, blaNDM, blaIMP, blaVIM,and blaOXA-48-like), plasmid-mediated AmpC β-lactamase-encoding genes (blaAAC, blaFOX, blaMOX, blaDHA, blaCIT, and blaEBC) and β-lactamase-encoding genes (blaTEM, blaSHV, blaCTX-M, and blaOXA-1) by PCR using previously reported primers.Citation17 To identify all genes detected in the PCR assays, PCR amplicons were sequenced, and the sequences were compared with those available in the National Center for Biotechnology Information GenBank database using BLAST searches (http://www.ncbi.nlm.nih.gov/BLAST).
Analysis Of Outer-Membrane Proteins
Outer-membrane proteins were extracted and quantified as described previously.Citation18 After purification, most proteins, except for outer-membrane porins, were removed, and subsequently loaded onto a 12% sodium dodecyl sulfate-polyacrylamide gel. After 150-min electrophoresis at 80 V in a Mini-PROTEAN Tetra set-up (Bio-Rad, People’s Republic of China), the gel was stained with 0.1% Coomassie brilliant blue (Sangon, Shanghai, People’s Republic of China). Based on the molecular weight of Omp35 and Omp36, porins could be easily identified on the gel. The coding sequences of omp35 and omp36, along with their respective promoter regions, were amplified as previously described,Citation19 and aligned with the respective sequences of E. aerogenes strain NCTC 10336 (GenBank accession no. KT894105.1 and KT894106.1, respectively).
Transformation And S1 Nuclease-PFGE
The transfer experiment was carried out in mixed lysogeny broth cultures. The 14 CRKA strains producing blaKPC-2 served as the donors while Escherichia coli DH5α served as the recipient strain. After a short incubation in ice, a mixture of chemically competent bacteria and plasmids, extracted from donor, was placed at 42°C for 45 s (heat shock) and then placed back in ice. SOC medium was added and the transformed cells were incubated at 37°C for 30 mins with shaking. Transformed bacteria were then plated and incubated at 37°C for 20 hrs.Citation20 Transformants were selected on lysogeny broth agar plates containing 50 mg L−1 ampicillin. S1 nuclease-PFGE was then performed to determine the location of blaKPC-2 gene, as described previously.Citation21
Incompatibility Group Analysis Of blaKPC-2-Carrying Plasmids
Next-generation sequencing, using the Illumina Miseq system, was performed to sequence the 14 blaKPC-2-carrying plasmids.Citation22 Sequencing reads were assembled de novo into contigs using SPAdes,Citation23 and manually inspected thereafter using Geneious 9. Gene predictions were conducted using the most popular ab-initio methods, Glimmer (v3.02) and Prodigal (v2.6.2). The consensus genes sets (with the same start or stop positions in parent scaffolds), predicted by both tools, were chosen as candidate gene models. The potential plasmids were identified by PlasmidFinder, and plasmid typing was performed using pMSI software.Citation24
Genetic Environment Of The blaKPC-2 Gene
A PCR mapping approach was adopted to compare the genetic context of blaKPC-2 gene in the 14 CRKA isolates with that of carbapenem-resistant K. pneumoniae strains reported in our hospital previously, using the same primers.Citation25 The obtained amplification products were sequenced and compared with those available in the National Center for Biotechnology Information GenBank database using BLAST searches (http://www.ncbi.nlm.nih.gov/BLAST).
Outbreak Characteristics
Outbreak data were collected via chart review for each patient from the hospital’s uniform electronic database. The following parameters were assessed: 1) general demographics, such as age, sex, and other background information; 2) the ward to which the patient was assigned after admission; and 3) previous use of antibiotics, particularly carbapenems.
Results
Prevalence, Antibiotic Susceptibility, And Resistance Mechanism Of CRKA Isolates
A total of 14 isolates were screened out as CRKA based on the disc diffusion results and confirmed by broth microdilution method. This accounts for a rate of 23.0% of the 61 non-duplicated clinical K. aerogenes strains isolated during 2013.
MICs of the 14 isolates to the 17 antimicrobial agents tested are listed in . All the isolates were non-susceptible to ertapenem, imipenem, meropenem, cefotaxime, ceftazidime, cefoxitin, cefepime, piperacillin/tazobactam, cefuroxime, cefoxitin, aztreonam, and fosfomycin. Treatment with the efflux pump inhibitor PAβN did not decrease the MICs of carbapenems in any of the CRKA isolates tested ().
Table 1 In-Vitro Activities Of Antimicrobial Agents Against CRKA Isolates
Table 2 Carbapenem MICs, β-Lactamase Detection, And blaKPC-2-Carrying Plasmid Analysis In The Isolates Studied
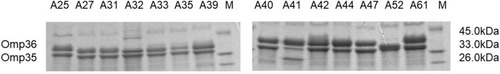
Moreover, SDS-PAGE analysis revealed the loss of Omp36 porin from isolate A52, whereas no such loss was found in the other 13 CRKA isolates (); omp36 in isolate A52 failed to be amplified after several trials with different primers. No mutation in omp35 or omp36 was found in the other isolates.
Figure 1 Outer-membrane protein profiles of 14 clinical K. aerogenes isolates on a 12% sodium dodecyl sulfate-polyacrylamide gel.
Abbreviations: K. aerogenes, Klebsiella aerogenes; M, marker.
β-Lactamase Characterization And Genetic Diversity
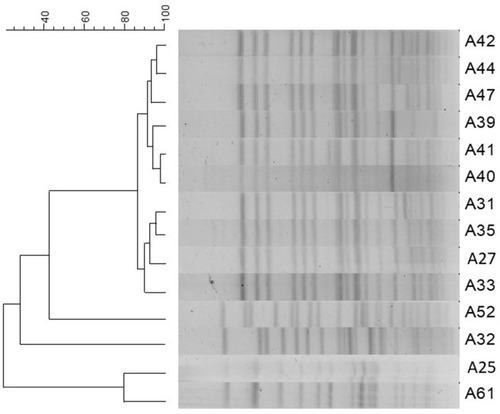
The PFGE results indicated that the 14 CRKA isolates belonged to five distinct PFGE groups (types A–E), with type A being the most dominant, including 71.4% (10/14) of the K. aerogenes isolates ().
Figure 2 PFGE patterns for CRKA. Isolates with 85% similarity were considered the same type.
Abbreviations: PFGE, pulsed-field gel electrophoresis; CRKA, carbapenem-resistant Klebsiella aerogenes.
All 14 isolates carried the blaKPC-2 gene. Six isolates (42.9%) carried the blaCTX-M-15 gene, two isolates (14.3%) carried the blaCTX-M-14 gene, and three isolates (21.4%) carried the blaSHV-1 gene. No PCR product was obtained for any of the other genes investigated (). The 14 blaKPC-2-carrying plasmids, obtained in the present study, belonged to various incompatibility groups, with IncFII and IncN being the most dominant.
Carbapenem resistance was successfully transferred from the 14 CRKA strains to E. coli DH5α by transformation. S1 nuclease-PFGE indicated that the size of the blaKPC-2-carrying plasmid ranged from approximately 20 kb to 200 kb (Figure S1). Notably, the 14 transformants simultaneously harbored the blaKPC-2 and blaCTX-M genes while blaSHV was not detected in any of the transformants ().
Genetic Environment Of The blaKPC-2 Gene
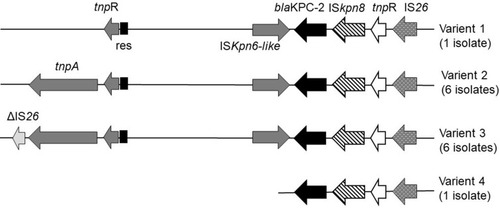
The blaKPC-2-bearing genetic structure of the 12 KPC-2-producing CRKA isolates was determined to be Tn1721- blaKPC-2-ΔTn3-IS26 with (6 isolates) or without (6 isolates) a truncated IS26 element fused at the left-most end of Tn1721. The other two strains only showed a partial version of this structure, which indicated variation in the Tn1721 region ().
Figure 3 Genetic environment of the blaKPC-2 gene from clinical isolates. Genes are depicted as arrows according to the direction of transcription. Variant 1 included isolate A25. Variant 2 included isolates A27, A31, A33, A35, A44, and A47. Variant 3 included isolates A32, A39, A40, A41, A42, and A61. Variant 4 included isolate A52.
Outbreak Characteristics
The epidemiological data of three patients (A25, A31, and A32) were not available since they had been hospitalized in a branch hospital. The remaining 11 patients (eight males and three females, aged 14–67 years) were all hospitalized in the neurosurgery ward of Huashan Hospital, except for one patient (A61) that was hospitalized in the intensive care unit. The patients’ clinical characteristics are listed in . Patient 1, from whom the CRKA isolate A27 was identified (from a sputum sample), had undergone surgery in February 2013 and developed post-operative pneumonia. Nearly 3 weeks after this, the first CRKA isolate was identified. Patient 2, who was in the same ward, was diagnosed with an invasive infection with CRKA (A33, PFGE type A). After 3 months, five additional patients successively tested positive for CRKA isolates within 12 weeks (PFGE type A). All these patients experienced actual pneumonia caused by CRKA. Overall, 63.6% of the patients (7/11) had undergone surgical procedures and 81.8% (9/11) had received mechanical ventilation.
Table 3 Clinical Characteristics Of Patients Infected With CRKA
Discussion
Although CRKA generally represents a smaller proportion of CRE, compared to carbapenem-resistant K. pneumoniae and E. coli, our findings demonstrated a quite relevant proportion of such strains, thus demanding more attention in this regard. Resistance rates against amikacin and gentamicin were higher than 75%. One isolate was found to be resistant to colistin; there were, indeed, some reports of colistin resistance in K. aerogenes during this time.Citation26,Citation27 Since colistin is generally considered a “last-line” antibiotic, this emerging resistance is highly concerning and worth highlighting.
Study of the resistance mechanism indicated that efflux pumps are not involved in the resistance, despite a previous report of their important role in carbapenem resistance.Citation28 Moreover, porin dysfunction is probably not the main mechanism of resistance in these isolates.
A study conducted in Italy demonstrated blaKPC as the major mechanism (97%) of carbapenem resistance in K. pneumoniae.Citation29 In contrast, in Thailand and Hong Kong, the rate of KPC-producing isolates was reported to be very low among CRE isolates.Citation30,Citation31 Therefore, although KPC-producing isolates have clearly spread worldwide, there are still regional differences in their prevalence. KPC was first discovered in the USA in 1996Citation32 and has spread worldwide since. Till date, KPC has been divided into 22 types, with KPC-2 dominating in People’s Republic of China.Citation33,Citation34
Transposable element Tn1721 is a proven transposon that has often played an important role in the dissemination of blaKPC-2 gene in People’s Republic of China. On the contrary, Tn4401 is the dominant transposon in Western countries. Most KPC-2 carrying Tn1721 transposons were reported in ST11 K. pneumoniae. Since studies of Tn1721 transposon acting in other species are rare,Citation35 in this study, we investigated Tn1721- blaKPC-2-ΔTn3-IS26 transposon in CRKA that shared the same genetic background with KPC-2-producing K. pneumoniae isolates, collected from the same hospital during the same period, as determined previously,Citation25 thus suggesting a common genetic structure for Enterobacteriaceae. Results indicated that Tn1721 transposons could not only transfer horizontally through K. pneumoniae, but could also transfer across different species, in this case through K. pneumoniae to K. aerogenes. Since IncFII and IncN are two dominant plasmid incompatibility groups detected in 14 isolates, the possibility of contribution of transposon as well as vertical transmission is stressed for the transformation of blaKPC-2.
The 11 CRKA strains in the dominant subgroup A that produced KPC-2 were likely to be primarily responsible for the first isolation and subsequent dissemination in the neurosurgery ward. The outbreak characteristic data showed mechanical ventilation, and previous use of broad-spectrum antibiotics could be potential risk factors for the acquisition of CRKA across the hospitalized patients.
Thus, the increased prevalence of carbapenem-resistant isolates in our hospital may have been caused by a failure to control the spread of these resistant strains, pointing at the importance of raising awareness of this situation in People’s Republic of China, as well as worldwide.
Conclusion
In this study, we identified KPC-2 production as the major carbapenem-resistance mechanism of CRKA. Moreover, we confirmed that Tn1721 transposon could transfer through the same species or via different ones. KPC-producing isolates can readily cause clonal spread and transposon transformation/vertical transmission across different species in a hospital environment. However, the proportion of transposon transformation and plasmid vertical transmission in KPC-2 dissemination would require further study. Our current study investigated KPC-2 dissemination in K. aerogenes and shed light on transposable element and plasmid transmission for antibiotic resistance.
Author Contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (grant no. 81401699 to XQ).
Disclosure
The authors report no conflicts of interest in this work.
References
- Munoz-Price LS, Poirel L, Bonomo RA, et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect Dis. 2013;13(9):785–796. doi:10.1016/S1473-3099(13)70190-723969216
- Nordmann P, Naas T, Poirel L. Global spread of carbapenemase producing Enterobacteriaceae. Emerg Infect Dis. 2011;17(10):1791–1798. doi:10.3201/eid1710.11065522000347
- Bornet C, Davin-Regli A, Bosi C, Pages JM, Bollet C. Imipenem resistance of Enterobacter aerogenes mediated by outer membrane permeability. J Clin Microbiol. 2000;38(3):1048–1052.10698994
- Qin X, Yang Y, Hu F, Zhu D. Hospital clonal dissemination of enterobacter aerogenes producing carbapenemase KPC-2 in a Chinese teaching hospital. J Med Microbiol. 2014;63(2):222–228.24273320
- Guh AY, Bulens SN, Mu Y, et al. Epidemiology of carbapenem-resistant enterobacteriaceae in 7 US communities, 2012–2013. J Am Med Assoc. 2015;314(14):1479–1487.
- Voulgari E, Poulou A, Koumaki V, Tsakris A. Carbapenemase-producing Enterobacteriaceae: now that the storm is finally here, how will timely detection help us fight back? Future Microbiol. 2013;8(1):27–39.23252491
- Lee CR, Lee JH, Park KS, Kim YB, Jeong BC, Lee SH. Global dissemination of carbapenemase-producing Klebsiella pneumoniae: epidemiology, genetic context, treatment options, and detection methods. Front Microbiol. 2016;13(7):895.
- Papp-Wallace KM, Endimiani A, Taracila MA, Bonomo RA. Carbapenems: past, present, and future. Antimicrob Agents Chemother. 2011;55(11):4943–4960.21859938
- Ahn C, Syed A, Hu F, O’Hara JA, Rivera JI, Doi Y. Microbiological features of KPC-producing Enterobacter isolates identified in a U.S. hospital system. Diagn Microbiol Infect Dis. 2014;80(2):154–158.25053203
- Pollett S, Miller S, Hindler J, Uslan D, Carvalho M, Humphries RM. Phenotypic and molecular characteristics of carbapenem-resistant Enterobacteriaceae in a health care system in Los Angeles, California, from 2011 to 2013. J Clin Microbiol. 2014;52(11):4003–4009. doi:10.1128/JCM.01397-1425210072
- Wang X, Chen G, Wu X, et al. Increased prevalence of carbapenem resistant Enterobacteriaceae in hospital setting due to cross-species transmission of the blaNDM-1 element and clonal spread of progenitor resistant strains. Front Microbiol. 2015;16(6):695.
- Tzouvelekis LS, Markogiannakis A, Psichogiou M, Tassios PT, Daikos GL. Carbapenemases in Klebsiella pneumoniae and other Enterobacteriaceae: an evolving crisis of global dimensions. Clin Microbiol Rev. 2012;25(4):682–707. doi:10.1128/CMR.05035-1123034326
- Wilson KH, Blitchington RB, Greene RC. Amplification of bacterial 16S ribosomal DNA with polymerase chain reaction. J Clin Microbiol. 1990;28(9):1942–1946.2095137
- CLSI.M100 Performance Standards for Antimicrobial Susceptibility Testing. Wayne, PA: Clinical and Laboratory Standards Institute. 2019.
- Yang FC, Yan JJ, Hung KH, Wu JJ. Characterization of ertapenem-resistant Enterobacter cloacae in a Taiwanese University Hospital. J Clin Microbiol. 2012;50(2):223–226. doi:10.1128/JCM.01263-1122135256
- Tenover FC, Arbeit RD, Goering RV, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33(9):2233–2239.7494007
- Dallenne C, da Costa A, Decré D, Favier C, Arlet G. Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J Antimicrob Chemother. 2010;65(3):490–495. doi:10.1093/jac/dkp49820071363
- Gayet S, Chollet R, Molle G, Pagès JM, Chevalier J. Modification of outer membrane protein profile and evidence suggesting an active drug pump in Enterobacter aerogenes clinical strains. Antimicrob Agents Chemother. 2003;47(5):1555–1559. doi:10.1128/aac.47.5.1555-1559.200312709321
- Doumith M, Ellington MJ, Livermore DM, Woodford N. Molecular mechanisms disrupting porin expression in ertapenem-resistant Klebsiella and Enterobacter spp. clinical isolates from the UK. J Antimicrob Chemother. 2009;63(4):659–667. doi:10.1093/jac/dkp02919233898
- Bergmans HEN, Van Die IM, Hoekstra WPM. Transformation in Escherichia coli: stages in the process. J Bacteriol. 1981;146(2):564–570.7012133
- Barton BM, Harding GP, Zuccarelli AJ. A general method for detecting and sizing large plasmids. Anal Biochem. 1995;226(2):235–240. doi:10.1006/abio.1995.12207793624
- Du H, Chen L, Chavda KD, et al. Genomic characterization of Enterobacter cloacae isolates from China that coproduce KPC-3 and NDM-1 carbapenemases. Antimicrob Agents Chemother. 2016;60(4):2519–2523. doi:10.1128/AAC.03053-1526787700
- Bankevich A, Nurk S, Antipov D, et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012;19(5):455–477. doi:10.1089/cmb.2012.002122506599
- Carattoli A, Zankari E, Garciá-Fernández A, et al. In Silico detection and typing of plasmids using plasmidfinder and plasmid multilocus sequence typing. Antimicrob Agents Chemother. 2014;58(7):3895–3903. doi:10.1128/AAC.02412-1424777092
- Wang L-H, Wei -D-D, Wan L-G, Yu Y, Deng Q, Liu Y. Diversity of the genetic environment of the bla KPC-2 gene among Klebsiella pneumoniae clinical isolates in a Chinese Hospital. Microb Drug Resist. 2015;22(1):15–21. doi:10.1089/mdr.2014.028126313117
- Diene SM, Merhej V, Henry M, et al. The rhizome of the multidrug-resistant enterobacter aerogenes genome reveals how new “Killer Bugs” are created because of a sympatric lifestyle. Mol Biol Evol. 2013;30(2):369–383. doi:10.1093/molbev/mss23623071100
- Zeng KJ, Doi Y, Patil S, Huang X, Tian GB. Emergence of the plasmid-mediated mcr-1 gene in colistin-resistant Enterobacter aerogenes and Enterobacter cloacae. Antimicrob Agents Chemother. 2016;60(6):3862–3863. doi:10.1128/AAC.00345-1626976876
- Szabó D, Silveira F, Hujer AM, et al. Outer membrane protein changes and efflux pump expression together may confer resistance to ertapenem in Enterobacter cloacae. Antimicrob Agents Chemother. 2006;50(8):2833–2835. doi:10.1128/AAC.01591-0516870780
- Conte V, Monaco M, Giani T, et al. Molecular epidemiology of KPC-producing Klebsiella pneumoniae from invasive infections in Italy: increasing diversity with predominance of the ST512 clade II sublineage. J Antimicrob Chemother. 2016;71(12):3386–3391. doi:10.1093/jac/dkw33727585968
- Netikul T, Kiratisin P, Nguyen MH. Genetic characterization of carbapenem-resistant enterobacteriaceae and the spread of carbapenem-resistant klebsiella pneumonia ST340 at a university hospital in Thailand. PLoS One. 2015;10(9):e0139118. doi:10.1371/journal.pone.013911626436895
- Ho PL, Cheung YY, Wang Y, et al. Characterization of carbapenem-resistant Escherichia coli and Klebsiella pneumoniae from a healthcare region in Hong Kong. Eur J Clin Microbiol Infect Dis. 2016;35(3):379–385. doi:10.1007/s10096-015-2550-326740321
- Yigit H, Queenan AM, Anderson GJ, et al. Novel carbapenem-hydrolyzing β-lactamase, KPC-1, from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob Agents Chemother. 2001;45(4):1151–1161. doi:10.1128/AAC.45.4.1151-1161.200111257029
- Dai W, Sun S, Yang P, Huang S, Zhang X, Zhang L. Characterization of carbapenemases, extended spectrum β-lactamases and molecular epidemiology of carbapenem-non-susceptible Enterobacter cloacae in a Chinese hospital in Chongqing. Infect Genet Evol. 2013;14:1–7. doi:10.1016/j.meegid.2012.10.01023220359
- Zhou T, Zhang X, Guo M, et al. Phenotypic and molecular characteristics of carbapenem-non-susceptible Enterobacteriaceae from a teaching hospital in Wenzhou, Southern China. Jpn J Infect Dis. 2013;66(2):96–102.23514904
- Tang Y, Li G, Liang W, Shen P, Zhang Y, Jiang X. Translocation of carbapenemase gene blaKPC-2 both internal and external to transposons occurs via novel structures of Tn1721 and exhibits distinct movement patterns. Antimicrob Agents Chemother. 2017;61(10):e01151–17.28784666