

Abstract
Serum albumin concentration (CP) is a remarkably strong prognostic indicator of morbidity and mortality in both sick and seemingly healthy subjects. Surprisingly, the specifics of the pathophysiology underlying the relationship between CP and ill-health are poorly understood. This review provides a summary that is not previously available in the literature, concerning how synthesis, catabolism, and renal and gastrointestinal clearance of albumin interact to bring about albumin homeostasis, with a focus on the clinical factors that influence this homeostasis. In normal humans, the albumin turnover time of about 25 days reflects a liver albumin synthesis rate of about 10.5 g/day balanced by renal (≈6%), gastrointestinal (≈10%), and catabolic (≈84%) clearances. The acute development of hypoalbuminemia with sepsis or trauma results from increased albumin capillary permeability leading to redistribution of albumin from the vascular to interstitial space. The best understood mechanism of chronic hypoalbuminemia is the decreased albumin synthesis observed in liver disease. Decreased albumin production also accounts for hypoalbuminemia observed with a low-protein and normal caloric diet. However, a calorie- and protein-deficient diet does not reduce albumin synthesis and is not associated with hypoalbuminemia, and CP is not a useful marker of malnutrition. In most disease states other than liver disease, albumin synthesis is normal or increased, and hypoalbuminemia reflects an enhanced rate of albumin turnover resulting either from an increased rate of catabolism (a poorly understood phenomenon) or enhanced loss of albumin into the urine (nephrosis) or intestine (protein-losing enteropathy). The latter may occur with subtle intestinal pathology and hence may be more prevalent than commonly appreciated. Clinically, reduced CP appears to be a result rather than a cause of ill-health, and therapy designed to increase CP has limited benefit. The ubiquitous occurrence of hypoalbuminemia in disease states limits the diagnostic utility of the CP measurement.
Disclosure
The authors report no conflicts of interest in this work.
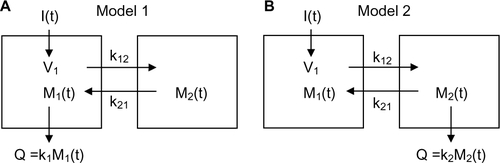
Figure S1 Schematic diagram of the two different compartmental models.
Notes: I(t): input to system as function of time t; Mi(t): amount of solute in compartment i; ki: rate constant for excretion from compartment i; kij: rate constant for exchange from compartment i to compartment j; Q: amount of excretion.
Supplementary materials
Measurements of albumin synthesis and catabolism
There are two fundamentally different approaches that have been used to measure human albumin synthesis and catabolism. The first approach is based on the analysis of the clearance of a bolus injection of a tracer of albumin. In the steady state, this clearance (ie, catabolism) must be equal to the albumin synthesis rate. Since albumin has a half-time of about 17 days, this approach requires measurements of tracer albumin over 2–3 weeks during which it is assumed that there is steady state, conditions that may not be valid in many disease states. The second approach uses labeled albumin precursors to measure the acute rate of liver albumin synthesis over a period of a few hours. This latter approach does not provide any direct measurement of albumin catabolic rates. Each of these methods is discussed here.
Notation:
D = tracer albumin dose
Cp(t) = tracer plasma albumin concentration as function of time
= steady state, nonlabeled, plasma albumin concentration
Q = total body metabolic rate
MT = total amount of nonlabeled albumin in body
M1(t) = amount of tracer in compartment 1 (plasma) as function of time
M2(t) = amount of tracer in compartment 2 (extravascular) as function of time
kout = Q/MT = fractional turnover rate for whole body
k1 = metabolic rate constant of compartment 1
k2 = metabolic rate constant of compartment 2
k12 = rate constant for exchange from compartment 1 to compartment 2
k21 = rate constant for exchange from compartment 2 to compartment 1
tT = turnover time = 1/kout
Clss = steady state clearance
Vss = steady-state volume of distribution
A1, A2 = coefficient of first (second) exponential describing tracer concentration
a1, a2 = rate constant of first (second) exponential describing tracer concentration
Steady-state measurement of albumin clearance and volume of distribution
The standard tracer pharmacokinetic experiment is to inject a tracer dose of albumin (eg, 131I-albumin) and determine the plasma tracer concentration (C(t)) over a time period that is long enough to be able to accurately extrapolate to infinity. The goal of these tracer measurements is to determine the two fundamental steady-state physiological parameters: Q, the total rate of metabolism of albumin, and MT, the total amount of albumin in the body. These two factors are determined experimentally from two related pharmacokinetic parameters determined from the tracer kinetic experiment, the steady-state clearance (Clss) and volume of distribution (Vss), defined by:
(S1)
where
is the steady-state plasma albumin concentration. In addition, one can define the “fractional turnover rate” and the “turnover time”:
(S2)
There are two different theoretical approaches that have been used to determine the albumin pharmacokinetics (compartmental and noncompartmental), and each of these is discussed in the Supplementary material. The compartmental approach used in the older literature describes the kinetics by an equivalent two- or three-compartmental system and determines Clss and Vss from the exponential decay at long durations (from 5 to 17 days). The major assumption required for this compartmental approach is that the exchange between the plasma and extravascular space is fast relative to the metabolic rate. This is a good assumption for albumin where the turnover rate is about 25 days while the intercompartmental exchange rate is about 1 day.Citation1 The noncompartmental approach, first described by Meier and ZierlerCitation2 in 1955, has been used in the more recent publications. The only theoretical assumption required for this approach is that the albumin metabolism occurs in the plasma compartment, that is, the effective albumin concentration at the site of metabolism is equal to the plasma concentration. There is some experimental evidence that indirectly supports this assumption, and it is usually assumed that it is valid for albumin.
Noncompartmental pharmacokinetic analysis
This is a general approach that does not require any assumptions about the exchange kinetics between plasma and the different tissue interstitial spaces. It is assumed that the system is linear. The basic experiment is to input a tracer dose D and then measure the tracer concentration C(t) as a function of time. It can be shown that under very general conditions the steady-state clearance (Clss) is described by the following relation:
(S3)
This is valid for an arbitrary input time course of the dose D and metabolism of albumin in any or all of the different extravascular spaces.
There is a similar, but less general, expression for Vss:
(S4) where I(t) is the time course of tracer input. Usually, it can be assumed that the input time course is a bolus input (I(t) = δ(t)) with very short time course compared to the albumin distribution:
(S5)
This relation is less general than the Clss relation (EquationEquation S3(S3) ) and is only valid if the metabolism occurs in the plasma compartment. If the metabolism occurs in an extravascular compartment, Vss cannot be determined without some additional assumptions about the pharmacokinetics in the extravascular space.Citation3 As shown later for the two compartment models (Figure S1), this Vss relationship should be a good approximation for albumin even if the metabolism occurs in the extravascular space because the blood–tissue exchange rate is relatively fast (about 1 day) compared with the metabolic rate (about 20 days).
Compartment pharmacokinetic analysis
In this approach, it is assumed that the kinetics can be described by a compartmental model from which the clearance can then be determined. This is discussed in detail here for a two-compartment model, but the same analysis would apply to more complicated three-compartment models.
Two different limiting cases of the two-compartment model is shown earlier, differing in the site of albumin metabolism. M1 and M2 are the amounts of solute in the two compartments, V1 is the volume of compartment 1 and k12 and k21 are exchange rates from compartment 1 to compartment 2 and in the opposite direction, respectively. The concentration in compartment 1 is equal to:
(S6)
In Model 1, the albumin metabolism (Q) occurs from compartment 1:
(S7)
In Model 2, the metabolism occurs in compartment 2:
(S8)
For either model, C1(t) for a bolus dose D is described by a sum of two exponentials:
(S9)
Substituting EquationEquation S9(S9) into the noncompartmental EquationEquations S3
(S3) and EquationS4
(S4) :
(S10)
Model 1 is described by the following two differential equations:
(S11) where I1(t) is the time-dependent input to compartment 1. Solving EquationEquation S11
(S11) for the case where I1(t) is a bolus input of dose D (I1=Dδ(t)), M1(t) is described by a two-exponential function:
(S12)
This is a surprisingly complex expression, and it can be seen that the coefficients for the two exponentials do not, in general, have a simple relationship to the parameters describing the two individual compartments.
The steady-state clearance can be determined directly from the steady-state solution to EquationEquation S11(S11) . Using EquationEquation S7
(S7) for the definition of Q:
(S13)
The steady-state volume of distribution is defined by EquationEquation S1(S1) :
(S14)
Since at steady state from EquationEquation S11(S11) , M2ss=(k12/k21)M1ss:
(S15)
One obtains exactly the same values for Clss and Vss using the general noncompartmental relations (EquationEquations S3(S3) –EquationS5
(S5) for bolus input) and using EquationEquation S10
(S10) with the coefficients for Model 1 (EquationEquation S12
(S12) ). This is expected since in Model 1 the metabolism occurs in the plasma compartment, and for this condition, these noncompartmental relations are always satisfied.
The corresponding differential equations for Model 2 are:
(S16)
Solving, EquationEquation S16(S16) for the case where I1(t) is a bolus input of dose D (I1=D δ(t)), M1(t) is described by a two-exponential function:
(S17)
The steady-state relation between M1 and M2 is obtained from EquationEquation S16(S16) :
(S18)
Solving EquationEquation S16(S16) for the steady-state clearance:
(S19)
Using the general noncompartmental relations (EquationEquations S3(S3) –EquationS5
(S5) for bolus input) and using EquationEquation S10
(S10) with the coefficients for Model 2 (EquationEquation S17
(S17) ), one obtains exactly the same value for Clss. This is expected since the noncompartment Clss relation is valid even if the metabolism is occurring in an extravascular compartment, as it is for Model 2.
Substituting EquationEquation S18(S18) in EquationEquation S14
(S14) , the steady-state volume of distribution for Model 2 is:
(S20)
The noncompartment relationship for Vss should not be valid for Model 2 because the metabolism does not occur in the plasma compartment. As predicted, substituting the Model 2 coefficients (EquationEquation S17(S17) in EquationEquations S10
(S10) and EquationS5
(S5) ), the noncompartmental prediction
is incorrect (ie, differs from EquationEquation S20
(S20) ):
(S21)
It can be seen that if k21≫k2 (the exchange between the two compartments is fast compared with the metabolic rate), then the noncompartmental expression becomes equal to the true Vss. This is a good approximation for albumin where the metabolic rate is about 20 days while the intercompartmental exchange is about 1 day.
At durations greater than ~4 days, the second term in EquationEquation S12(S12) or EquationS17
(S17) becomes negligible relative to the first and the concentration in compartment 1 (eg, plasma) can be approximated by:
(S22)
Although not always explicitly stated, the albumin compartmental kinetic analysis depends on the assumption that the exchange between the two compartments is fast compared with the metabolic rate, that is, k1≪k12 (Model 1) or k2≪k21 (Model 2). Using this assumption, the expression for B in EquationEquation S12(S12) can be approximated by:
(S23)
Substituting this approximation for B into the Model 1 expression for a1 (EquationEquation S12(S12) ):
(S24) where the last equality uses the Model 1 steady-state clearance and volume of distribution relations (EquationEquations S13
(S13) and EquationS15
(S15) ). Substituting this approximation for B in the Model 1 expression for A1 (EquationEquation S12
(S12) ) and assuming k1≪k12:
(S25)
Finally, substituting these expressions for a1 and A1 into EquationEquation S22(S22) :
(S26)
Although this was explicitly derived for Model 1, it can be shown that this same approximation is valid for Model 2 at durations greater than ~4 days and assuming that k2≪k21. In summary, with this assumption the tracer plasma concentration for the two-compartmental model at durations greater than ~4 days can be approximated by a single exponential with a rate constant α equal to the “fractional turnover” rate for the whole body (EquationEquation S2(S2) ) and the zero time intercept equal to the tracer-injected dose (D) diluted in the steady-state volume of distribution, allowing the experimental determination of both Clss and Vss. This result provides the theoretical justification for most of the older albumin tracer kinetic analysis.
Acute measurements of albumin synthesis
In this approach, labeled albumin precursors are given as a bolus injection and the rate of appearance of these labels in albumin is measured over a period of few hours. This provides a direct measurement of the short-term fraction of intravascular (ie, plasma) albumin synthesized per hour referred to as the “fractional synthesis rate.” Fractional synthesis is related to the plasma albumin clearance (ClP) and plasma volume (Vp) by the relation:
(S27)
The absolute synthesis rate is then determined from EquationEquation S27(S27) with the plasma volume measured by some technique such as 125I albumin distribution.
Initially, studies used 14carbonate infusions to label the arginine guanido carbon via urea cycle metabolism with the assumption that the specific activity of the arginine precursor for albumin synthesis is identical to the urea-specific activity.Citation4
(S28)
The urea synthesis rate is directly measured using 2C urea. These were short-term experiments, with the specific activity usually measured at 5 hours (additional small corrections are made for albumin removal from plasma during this 5-hour period).
In recent years, the standard approach for measuring albumin synthesis has been based on the precursor “flooding technique” in which a large bolus of ([1-13C] leucineCitation5 or [2Hring] phenyalanineCitation6) is injected, uniformly labeling all the precursor pools, and albumin isotopic enrichment is measured versus time.Citation5 There is a lag period (the “secretion time”) in the appearance of enriched albumin of about 30 minutes. The fractional synthesis rate is determined from:
(S29)
The two time points are selected for a period when the enrichment is increasing roughly linearly (after the lag period) – usually between about 40 and 80 minutes after the bolus injection. A major advantage of using this relatively short time interval (ie, 40 minutes) is that no correction is required for loss of albumin from the intravascular space by metabolism or diffusion.Citation5
References
- TakedaYReeveEBStudies of the metabolism and distribution of albumin with autologous I131-albumin in healthy menJ Lab Clin Med19636118320213984638
- MeierPZierlerKLOn the theory of the indicator-dilution method for measurement of blood flow and volumeJ Appl Physiol195461273174413174454
- NakashimaEBenetLZAn integrated approach to pharmacokinetic analysis for linear mammillary systems in which input and exit may occur in/from any compartmentJ Pharmacokinet Biopharm19891766736862635739
- McFarlaneASIronsLKojARegoecziEThe measurement of synthesis rates of albumin and fibrinogen in rabbitsBiochem J19659553654014340105
- BallmerPEMcNurlanMAMilneEHeysSDBuchanVCalderAGGarlickPJMeasurement of albumin synthesis in humans: a new approach employing stable isotopesAm J Physiol19902596 Pt 1E797E8032260648
- BallmerPEReichenJMcNurlanMASterchiABAndersonSEGarlickPJAlbumin but not fibrinogen synthesis correlates with galactose elimination capacity in patients with cirrhosis of the liverHepatology199624153598707282