
Abstract
Background
Procoagulant microvesicles (MVs) are submicron membrane fragments released from activated cells and cells undergoing apoptosis. The procoagulant activity of MVs is enhanced in the presence of tissue factor (TF). MVs and TF are active mediators that induce pro-inflammatory response and prothrombotic tendency and have been linked to the severity of several disorders, including malaria infection. The current study aimed to measure the levels of circulating procoagulant MVs and TF-bearing MVs in malaria patients and correlate these levels with other hematological parameters and parasitemia.
Materials and Methods
Levels of MVs and TF-bearing MVs in the plasma of children and adult patients infected with Plasmodium falciparum were measured alongside matched healthy controls.
Results
Patients with Plasmodium falciparum infection had ~3.8 times MVs (p < 0.0001) and ~13.0 times TF-bearing MVs compared to the matched healthy controls. MVs showed inverse significant correlation with platelet count (p = 0.0055), hemoglobin (p = 0.0004) and parasitemia.
Conclusion
Elevated levels of MVs and TF-bearing MVs could be useful biomarkers to evaluate the procoagulant activity, inflammatory response and parasitemia levels in malaria infection, aiding in better management of the disease.
Introduction
Malaria is one of the major causes of morbidity and mortality globally. The World Health Organization (WHO) considers malaria a major health burden, with a total of 241 million cases reported in 2020, compared to 227 million cases in 2019.Citation1 Malaria is especially prevalent in tropical and subtropical countries, with 95% of the cases occurring in Africa.Citation2 Although the incidence of malaria in Saudi Arabia has been decreasing over the years, mainly due to the effectiveness of the control strategies under the malaria elimination program, the Jazan region in southwestern Saudi Arabia (near the African horn) continues to be considered an endemic area of malaria.Citation1 Several reports have indicated the presence of hypercoagulability in malarial infection, particularly in infections caused by Plasmodium falciparum, which is characterized by increase in inflammatory response, activation of coagulation factors, low levels of coagulation inhibitors, destruction of platelets, chronic activation of endothelial cells (ECs), disruption of the phospholipid structure of red blood cells (RBCs), and the generation of microvesicles (MVs).Citation3,Citation4 Some of these hypercoagulability factors have been linked to the development of disseminated intravascular coagulation (DIC) in Plasmodium falciparum infection. Furthermore, thrombosis has been reported in cerebral malaria, a severe complication of Plasmodium falciparum infection.Citation5
MVs are submicron membrane fragments, which are released from cells undergoing apoptosis and from activated cells, mainly platelets.Citation6 MVs are heterogeneous in size (ranging from 0.1 to 1.0 μm) and content (containing antigenic features of their parent cells). Furthermore, MVs can be derived from almost all cell types in vivo and in vitro and can be found in blood, tissues, urine, saliva, and all other body fluids. Elevated levels of MVs and tissue factor-bearing MVs (TF-bearing MVs) have been observed in a broad range of physiological (such as during physical exerciseCitation7 and pregnancy)Citation8 and pathological conditionsCitation9–11 and in infectious diseases, including parasitic infection (mainly malaria infection)Citation12–18, viral infection,Citation19,Citation20 and bacterial infection.Citation21 MVs have been proposed to play a key role in the pathogenesis, mediating inflammatory responses, severity and manifestation in different type of malaria infection.Citation14,Citation15,Citation17
MVs play a key role in hemostasis, thrombosis, inflammation, cancer, angiogenesis, cell-to-cell communication, and waste removal.Citation22,Citation23 In addition, MVs have the ability to drive prothrombotic tendency in many diseases,Citation9 and they are linked to thrombosis and recurrent thrombosis.Citation24 The procoagulant activity of MVs is due to the anionic, or negatively charged, phospholipids, mainly phospholipids, ie, phosphatidylserine (PS), exposed on their surface due to the flip-flop mechanism of MV formation.Citation25 The anionic phospholipids enhance the binding of coagulation complexes and support thrombin generation in vivo and in vitro.Citation26 This procoagulant activity is enhanced in the presence of tissue factor (TF), the principal initiator of coagulation.Citation27,Citation28 Blood-borne TF and TF-bearing MVs have been observed in the circulation of many diseases.
TF is an integral membrane protein that is sequestrated from circulation under normal physiological condition. TF plays a major role in the activation of the coagulation system, and it is another procoagulant factor that contributes to hypercoagulability.Citation27,Citation28 Circulatory TF has been reported in malaria infection and has been shown to support coagulation complexes.Citation18
The current study aimed to measure the levels of circulating procoagulant MVs and TF-bearing MVs in malaria patients and correlate their levels with other hematological parameters and parasitemia, which could help in the better understanding, management, and prediction of the severity of the disease and future complications.
Materials and Methods
Study Design
The current cross-sectional study involved 48 malaria infected patients (Plasmodium falciparum) and 42 age and sex matched healthy controls residing in the Jazan region. Malaria infected patients were febrile patients diagnosed for malaria using rapid test and blood smear examination (thick and thin films with Giemsa stain) from 4 different hospitals in the Jazan region. The control group comprised only apparently healthy individuals.
Study Settings
The study was conducted in the Jazan region, located in the southwest of the Kingdom of Saudi Arabia.Citation29
Blood Sampling
Venous blood was collected from each patient into ethylenediaminetetraacetic acid (EDTA) and sodium citrate-anticoagulant tubes. Directly after collection, the EDTA tubes were used for complete blood count (CBC) analysis and for malaria diagnosis, which were used for malaria diagnosis using malaria rapid diagnostic test (RDT). Further, thin and thick blood films were prepared stained with diluted Giemsa stain. The sodium citrate-anticoagulant tube was used for the analysis of MVs and TF-bearing MVs.
Microscopy
The thin and thick blood smears were stained and used to assess the presence of malaria infection and parasite density as previously described.Citation30 The parasite density was estimated as described by Fransisca et al.Citation30 The parasite density was categorized into four groups as follows: <100 parasites/μL; 100–999 parasites/μL; 1000–9999 parasites/μL; and ≥10,000 parasites/μL.Citation31
AllTest Malaria P.f./P.v. Rapid Test Cassette
This study used the AllTest Rapid Test—Malaria P.f./P.v. kit, product code IMPV-402 (Hangzhou AllTest Biotech, Hangzhou, China). This RDT detects Plasmodium falciparum-specific HRP2 on the P.f. test line according to the manufacturer's guidelines.
Complete Blood Cell Measurement
The EDTA tubes were used for CBC measurement using Sysmex XN-550 Hematology Analyzer (Sysmex, Kobe, Japan).
Plasma Preparation for the Measurement of MVs and TF-Bearing MVs
The sodium citrate tubes were centrifuged at room temperature for 30 min at 3000 rpm. Then, the plasma, the upper part of the blood, was separated and centrifuged again at 13,000 rpm for 2 min to obtain platelet-free plasma (PFP). The PFP was stored at −80°C for the MV and TF-bearing MV analysis.
Analysis of MVs and TF-Bearing MVs
The PFP was used for the measurement of (i) procoagulant MVs, using the Zymuphen MP-Activity enzyme-linked immunosorbent assay (ELISA) Kit (Aniara Diagnostica, OH, USA), and (ii) TF-bearing MVs, using the Zymuphen MP-TF ELISA Kit (Aniara Diagnostica, OH, USA). The measurements were conducted according to the manufacturer’s instructions.
The Zymuphen MP-Activity assay relies on the detection of the procoagulant phospholipids—ie, the negatively charged phospholipids, mainly PS—exposed on the surface of the MVs using Annexin-V streptavidin. The procoagulant phospholipids on the MVs will support the binding of the bovine activated factor V-activated factor X complex (FVa-FXa) and prothrombin added to the reaction mixture. This will allow the formation of the prothrombinase complex (FVa-FXa) on the surface of the captured MVs. The prothrombinase complex converts prothrombin to thrombin. Then, the data are expressed as nM PS equivalent. The Zymuphen MP-TF ELISA relies on the detection of TF on the outer surface of MVs using plate coated anti-TF. The TF on the sample mixture will form a complex with the added activated factor II-factor X (FVIIa-FX) known as TF-FVIIa-FX complex. This allows the activation of FXa. The color development is read, and the data are reported as pg/mL concentration of TF-bearing MVs.
Ethical Consideration
The current study was approved by the Jazan University Scientific Research Ethical Committee (Ref. No. REC42/1/122). Signed informed consents were obtained from the adult study participants and from the children’s guardians. The study was carried out according to the Declaration of Helsinki.
Statistical Analysis
The statistical analysis was performed using GraphPad Prism (version 8.0). The data in the current study are reported as mean ± standard deviation (SD). Chi-square test was applied for the analysis of the demographic data. Unpaired Student’s t-test was used for group comparison. In addition, single-sample t-test was used for the evaluation of correlation coefficients. Significant statistical differences were considered when p-values were less than 0.05.
Multivariate linear regression analysis was performed to assess the association of independent factors with malaria infection in the study. Variables with p <0.05 were included for multivariate analysis.
Results
Demographic Characteristics
A total of 48 malaria infected patients (37 males and 11 females) and 42 healthy controls (33 males and 9 females) matched in age and gender were recruited in the present study. The age range of the patient group was 3–61 (30.0 ± 14.3) years, while that of the control group was 4–59 (30.4 ± 15) years. There were no significant differences in age and gender between the study groups (p >0.5; ). All the patients had Plasmodium falciparum infection.
Table 1 Demographic and Hematological Characteristics for Malaria Patients and Controls
Plasmodium falciparum Parasitemia
Overall, 15 patients had low parasitemia, 10 had mild parasitemia, 13 had moderate parasitemia, and three had high parasitemia. Seven patients were excluded from the parasitemia evaluation as parasitemia was not observed in these patients.
Complete Blood Count Data
The CBC data for the patients and controls are shown in . The patients had slightly higher WBC count than the controls (p >0.5), but it was within the normal range. The RBCs (p <0.01), hemoglobin (Hb) (p <0.0001), and platelet count (p <0.01) were lower in the patients compared to controls and were statistically significant ().
Procoagulant MVs & TF-Bearing MVs
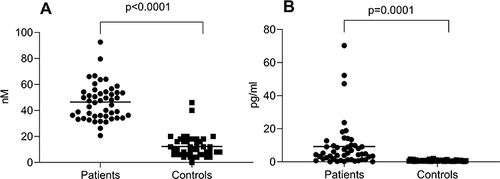
Malaria patients had significantly higher (~3.8 times) procoagulant MVs than the controls (46.4 ± 14 vs 12.3 ± 9.0; p <0.0001; and ). Additionally, malaria patients had significantly higher (~13.0 times) TF-bearing MVs than the controls (9.2 ± 13 vs 0.7 ± 0.4; p =0.0001; and ).
Table 2 Levels of Circulating Procoagulant MVs and TF-Bearing MVs in Malaria Patients and Controls
Figure 1 Levels of procoagulant microvesicles (A) and tissue factor-bearing microvesicles (B) in malaria patients (patients) compared to matched-healthy controls (controls). (A) Levels of microvesicles in the plasma of the patients compared to controls. (B) Levels of tissue factor-bearing microvesicles in the plasma of the patients compared to controls. Individual data are shown from malaria patients (n=48) and healthy controls (n=42) with the horizontal line representing the mean of data. The differences between the patients and controls were determined using independent unpaired t-test.
Correlation Studies of MVs and TF-Bearing MVs with RBCs, Hemoglobin, WBCs, and Platelets
Correlation studies showed significant positive correlation between procoagulant MVs and TF-bearing MVs. Furthermore, procoagulant MVs had significant negative correlation with platelet count (r = −0.6983, R squared = 0.4876; p <0.0001). TF-bearing MVs had negative correlation with platelet count in the studied population (patients and controls) (r = −0.2903, R squared = 0.08426; p = 0.0055; ). In addition, a significant negative correlation was observed with procoagulant MVs and Hb (r = −0.3623, R squared = 0.1313; p = 0.0004), while correlation with RBC count was not significant (r = −0.1788, R squared = 0.03198; p = 0.0899; ). On the other hand, TF-bearing MVs had no correlation with Hb and RBC count (p >0.05; ). Procoagulant MVs and TF-bearing MVs did not exhibit correlation with WBC count (p >0.05).
Table 3 Correlation Between (i) Microvesicles (MVs), (ii) Tissue Factor-Bearing Microvesicles (TF-MVs) and Hematological Parameters (RBCs, Hemoglobin [Hb] and Platelets) in the Studied Population (Combined Malaria Patients and Controls)
Multivariate Linear Regression of the MVs, TF-Bearing MVs, RBCs, Hb
Using stepwise multivariable analyses with MVs, TF-bearing MVs, RBCs, Hb, and platelets (). MVs (β=0.01, p <0.001), Hb (β=−0.052, p =0.001), and platelets (β=−0.001, p <0.001) were significantly associated with malaria.
Table 4 Multivariate Linear Regression of the Correlated Parameters (MVs, TF-Bearing, Platelets, RBCs, Hemoglobin [Hb]) with Controls and Patients with Malaria in Jazan Region
Classification of Procoagulant MVs and TF-Bearing MVs Based on Parasitemia Induced by Plasmodium falciparum
The procoagulant MV levels varied with the parasitemia level as follows: moderate (49.3 ± 12.5; n = 13) > mild (45.5 ± 10.6; n = 10) > low (42.6 ± 13.1; n = 15) (p >0.05) but not in high parasitemia (33.9 ± 7.0; n = 3). In addition, TF-bearing MVs were elevated in moderate parasitemia. This trend was not observed with platelets, RBCs, and Hb (p >0.05; ). All mean values were non-significantly different between the three severity groups of malaria (ANOVA, p >0.05).
Table 5 Sub-Analysis of MVs, TF-Bearing MVs and Hematological Parameters According to the Level of Parasitemia in Plasmodium falciparum Malaria Infection
Discussion
The current study investigated the levels of procoagulant MVs and TF-bearing MVs in malaria patients. The findings of this study reveal that the levels of circulating procoagulant MVs are higher in malaria patients compared to healthy controls ( and ). These findings are consistent with those of previous studies, which reported increased levels of procoagulant MVs in malaria patientsCitation12–16 and also in inflammatory conditions.Citation19,Citation20,Citation32 Campos et al suggested that MVs play a role in the paroxysms of non-immune patients infected with Plasmodium vivax.Citation14 In addition, MVs contribute to the pathogenesis and mediate inflammatory response induced by Plasmodium falciparumCitation17 and play a role in the pathogenesis of severe malaria infection, including in neurological manifestation.Citation15 Similar observation was reported in experimental cerebral malaria, and the absence of high levels of MVs was protective in mice. The protective mechanism is mainly due to the absence of the adenosine triphosphate (ATP)-binding cassette transporter A1 (ABCA1) gene.Citation33 The ABCA1 is involved in the regulation of the transmembrane lipid, mainly PS, and in the flip-flop mechanism, which is responsible for the formation of MVs.Citation33 MVs also contribute to the enhanced formation and sequestration of more MVs.Citation34
In the current study, the increase in the level of procoagulant MVs was ~3.8-fold, which is higher than the increase reported in dengue infection (~2.7-fold)Citation20 and known hypercoagulable state condition of sickle cell disease (~2.5-fold)Citation10 but less than that of coronavirus disease of 2019 (COVID-19) infection (~5.6-fold).Citation19 The method used in the study is a functional assay that can detect the ability of the procoagulant MVs to support thrombin formation. MVs have the ability to support coagulation complexes and subsequent thrombin generation in healthy individuals,Citation26 in malaria infection,Citation18 in other pathological conditions with inflammatory mechanism, including human endotoxemia,Citation35 and in cases of thrombosis (venous and arterial thrombosis).Citation24 The procoagulant activity of MVs has been found to be 100 times that of activated platelets.Citation36 The procoagulant activity of MVs increased in the presence of TF, and the negative charges on the surface of MVs enhanced TF.Citation37 Elevated levels of procoagulant MVs are considered a marker for prothrombotic tendency in many pathological conditions.Citation24,Citation26 In addition, MVs-associated thrombin generation has been posited as a useful marker to predict the risk of recurrence in patients with thrombosis.Citation24,Citation38 In vivo, procoagulant MVs and TF-bearing MVs have been shown to participate in the thrombus model of vessel injury.Citation39
Endothelial cells release procoagulant soluble TF in response to inflammatory cytokines, including tumor necrosis factor (TNF). Inflammatory cytokines, including TNF, are increased in malaria infection.Citation18,Citation40 The presence of circulatory TF-bearing MVs has been linked to thrombosis, the severity and mortality of COVID-19,Citation32,Citation41 and severe complications in malaria infection. The current study reported a significant increase (~13-fold) in the level of active TF (TF-bearing MVs) in the plasma of malaria patients. This observation has been reported previously in malaria infectionCitation18 and other disorders.Citation19,Citation20,Citation41 TF expression by ECs has been reported in Plasmodium falciparum infection, which has the ability to support coagulation complexes.Citation18 The expression of TF from ECs is mainly driven by the effect of parasitized RBC and parasitized RBC-derived MVs.Citation18 The exact mechanism is not fully known, but it can be postulated that MVs contain effective molecules that can be transferred to ECs. The presence of high level of circulatory TF-bearing MVs in the current study is of concern, as this could be an early sign of future complications, including thrombosis.
Elevated levels of TF have been attributed to an increase the procoagulant activity in acute coronary syndrome, DIC, and thrombosis.Citation42,Citation43 DIC is a common complication in complicated cerebral malaria.Citation44 The presence of fibrin clot, including D-dimer, fibrin degradation product, and soluble fibrin, and associated DIC is a feature of malaria infection, especially complicated malaria.Citation44 The presence of TF has been observed in some patients with cerebral malaria, which supports coagulation activation and fibrin deposition in the brain,Citation18 and has been associated with DIC.Citation44 In a case report, DIC was reported in a 56-year-old female with complicated Plasmodium falciparum infection.Citation45
Although the majority of circulatory MVs (70–90%) are derived from platelets in normal individuals at basal level,Citation46 elevated levels of circulatory MVs derived from different cell types have been found in different disorders.Citation47 In malaria patients infected with either Plasmodium falciparum or Plasmodium vivax, elevated levels of MVs derived from platelets, RBCs, ECs, and to lesser extent from monocytes have been observed and are associated with disease severity.Citation12–16 The current study showed inverse relationship between MVs and platelet count (P <0.05) and RBC count (P = 0.0899) (). MVs have been found to correlate with thrombocytopenia and coma depth in cerebral malaria.Citation15 The same study reported no correlation between MVs and RBCs, which is similar to the present findings. Thrombocytopenia was observed in the current study, as in previously published reports.Citation15 Plasmodium falciparum infection affects platelets and directly contributes to thrombocytopenia,Citation48 which is a feature of malaria infection regardless of its severity.Citation49 Platelets play various roles in hemostasis, thrombosis, inflammation, immunity, and angiogenesis due to their wide array of stored chemokines and chemoattractants;Citation50 they play a similar role in malaria infection.Citation51 Although thrombocytopenia is a prominent feature of malaria infection, especially in severe malaria infection, it has not been proposed to be a prognostic marker in children with cerebral malaria;Citation52 however, MVs have been proposed as a novel prognostic marker.Citation13,Citation17 Thus, it is thought that most of the MVs in the current study might have been derived from platelets, RBCs, and ECs. This justification could be explained by the involvement of platelets and RBCs in the pathogenesis of malaria infection, and the role of both the cells in altering ECs is well-documented in the case of malaria infection.Citation34 Moreover, RBC destruction is a common feature of malaria infection.Citation53 Parasitized red cells release 10 times more MVs than unparasitized red cells in vivo and in vitro.Citation12 Increased levels of circulatory endothelial- and platelets-derived MVs is a feature of thrombocytopenia.Citation54 Therefore, increased levels of circulating MVs derived from RBCs, platelets, and ECs are expected.
The findings of the present study showed an association between elevated levels of MVs with parasitemia and RBC count with parasitemia. The increase in the level of MVs with an increase in the load of the parasite can be used to predict parasite load and for parasitemia evaluation. Mfonkeu et al observed no correlation between RBC-derived MVs and parasitemia, which was possibly due to the treatment undergone by the patients in their study. In contrast, the present study recorded an increase in the MV levels with increase in parasitemia levels from low to mild to moderate. Although, thrombocytopenia frequently occurs in malaria infection, as mentioned previously, it can be used neither as a prognostic marker nor to predict parasitemia. The present findings confirmed that thrombocytopenia varies based on the levels of parasitemia (). Therefore, this study indicates that MVs might be used as a prognostic marker.Citation13,Citation17 TF-bearing MVs and Hb showed variations similar to that of thrombocytopenia. The highest MV levels were observed with moderate parasitemia and not with high parasitemia. This can be attributed to the number of patients with high parasitemia in the present study (high parasitemia patients [n = 3], moderate parasitemia [n = 13], mild parasitemia [n = 10], and low parasitemia [n = 15]). It is worth mentioning that the statistical analysis was non-significant when comparing the MV levels with parasitemia levels using unpaired t-test and one-way ANOVA.
As mentioned previously, the origin of the MVs in the current study was possibly from platelets and RBCs. Nantakomol et al reported high levels of RBC-derived MVs in parasitemia induced by Plasmodium falciparum compared to those induced by Plasmodium vivax and Plasmodium malariae.Citation12 The same study reported high levels of parasitemia in severe malaria but not in uncomplicated malaria. Therefore, it remains unclear whether elevated levels of MVs and their correlation with parasitemia can predict the severity and future complications of malaria infection, and further studies are needed to elucidate these preliminary findings.
The regulation of the inflammatory response in early stages of malaria infection is crucial. A turning point in malaria infection, which might lead to severe malaria, is the excessive production of pro-inflammatory cytokines due to dysregulation of the inflammation process.Citation55 MVs and TF play a role in fine-tuning regulation between hemostasis, thrombosis and inflammation.Citation56,Citation57 Moreover, the role of MVs and TF in driven inflammatory response in malaria infection has been suggested.Citation58,Citation59 It is well-known that MVs are immunogenicCitation60 and correlate with the levels of pro-inflammatory cytokine TNF in cerebral malaria,Citation16 while TF is considered a structural membrane of class two cytokine receptor family.Citation61 In an in vitro study on mice infected with Plasmodium berghei, MVs were found to activate macrophage and induce high inflammatory response, including TNF production.Citation62 Moreover, MVs are active carriers of effective mediators, such as TF, membrane components, chemoattractants, cytokines, and micro-RNA.Citation28,Citation60,Citation63 In the brain vessels of patients with Plasmodium falciparum malaria, platelet MVs have been shown to transfer the antigenic features of the platelet within MVs to infected RBCs, thus mediating the cytoadherence of the infected RBCs to ECs.Citation13,Citation17 MVs can internalize with infected Plasmodium falciparum RBCs.Citation60 Moreover, MVs have been shown to fuse with other cells ex vivo and in vitro.Citation64,Citation65 Other vesicles (exosomes) have been shown to transfer the effective mediators by fusing with the RBC membrane in Plasmodium falciparum infection, with possible involvement of Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) and Plasmodium falciparum erythrocyte membrane protein 3 (PfEMP 3).Citation66 Whether PfEMP1 and PfEMP3 play a role similar to that of exosomes in the fusion of MVs in malaria infection is not yet fully clear and needs further investigation. MVs have been proposed to be a potential novel mediator in the prediction of cerebral dysfunction in severe Plasmodium falciparum malaria.Citation15
The current study showed association of MVs, Hb, and platelets with malaria infection. The association of hematological parameters including low hemoglobin and thrombocytopenia with malaria infection has been reported previously.Citation67,Citation68 Therefore, understanding the exact role of the heterogenous MVs and TF-bearing MVs in malaria infection and their interaction with other cells could contribute to clarifying the crucial association between prothrombotic tendency and inflammatory responses that lead to complications in malaria infection.
The limitations of the current study include the lack of complicated malaria patients as well as did not consider the measurement of their inflammatory mediators, any thrombotic data, existence comorbidities of patients, and did not follow up with patients. In addition, the current study applied a single technique (functional assay) to evaluate the procoagulant activity of MVs and TF-bearing MVs, but combining different techniques including flow cytometry, transmission electron microscopy and nanoparticle tracking analysis to characterize the type, origin, size and absolute count of MVs and TF-bearing MVs will provide a more comprehensive picture of the behavior in malaria infection. Nevertheless, the data generated in the current study are in line and add to the growing field of MVs and TF-bearing research on malaria infection.
Conclusions
This study showed significantly elevated levels of procoagulant MVs and TF-bearing MVs in malaria patients; the MV levels are linked primarily to platelet count and only to a lesser extent to RBCs. The procoagulant MVs and TF-bearing MVs have the ability to support and initiate the formation of coagulation complexes, which contribute to the hypercoagulability in malaria infection. The procoagulant activity of MVs and TF enhances the hypercoagulability state in malaria, which increases the risk of prothrombotic tendency and augments the inflammatory responses. MVs and TF may constitute a potential novel mediator in the prediction of prothrombotic tendency, augmentation of inflammatory responses, and future complications in malaria patients. However, further research on the mechanism of the contribution of MVs and TF-bearing MVs are needed to explain the exact contribution to the hypercoagulability and inflammation. In addition, the association of MVs and TF-bearing MVs with other circulating ECs marker might explain the magnitude of ECs contribution in malaria infection severity and complication.
Abbreviations
MVs, microvesicles; TF, tissue factor; TF-MVs, tissue factor-bearing microvesicles; WHO, World Health Organization; ECs, endothelial cells; RBCs, red blood cells; WBCs, white blood cells; Hb, hemoglobin; DIC, disseminated intravascular coagulation; PS, phosphatidyl serine; CBC, complete blood count; PFP, platelet-free plasma; RDT, rapid diagnostic test; SD, standard deviation; PfEMP1, Plasmodium falciparum erythrocyte membrane protein 1; PfEMP 3, Plasmodium falciparum erythrocyte membrane protein 3; ABCA1, ATP-binding cassette transporter A1; TNF, tumor necrosis factor.
Data Sharing Statement
The data that support the findings of this study are available from the authors upon reasonable request.
Informed Consent Statement
Signed informed consents were obtained from the adult study participants or from children’ guardians. The study was carried out according to the Declaration of Helsinki.
Institutional Review Board Statement
The current study was approved by the Jazan University Research Ethical Committee (Ref. No. REC42/1/122), Jazan University.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.”
Disclosure
The authors declare no conflict of interest.
Acknowledgments
The authors would like to thank the administrative and laboratory staff of all participated hospitals for their support and fruitful cooperation. The authors would also like to thank all patients participated in this study.
Additional information
Funding
References
- World Health Organization. World malaria report 2021; 2021. Available from: https://www.who.int/publications/i/item/9789240040496. Accessed March 31, 2023.
- Vos T, Lim SS, Abbafati C, et al. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: a systematic analysis for the global burden of disease study 2019. Lancet. 2020;396(10258):1204–1222. doi:10.1016/S0140-6736(20)30925-9
- Ghosh K, Shetty S. Blood coagulation in falciparum malaria--a review. Parasitol Res. 2008;102(4):571–576. doi:10.1007/s00436-007-0832-0
- Turner GD, Ly VC, Nguyen TH, et al. Systemic endothelial activation occurs in both mild and severe malaria. Correlating dermal microvascular endothelial cell phenotype and soluble cell adhesion molecules with disease severity. Am J Pathol. 1998;152(6):1477–1487.
- Moxon CA, Alhamdi Y, Storm J, et al. Parasite histones are toxic to brain endothelium and link blood barrier breakdown and thrombosis in cerebral malaria. Blood Adv. 2020;4(13):2851–2864. doi:10.1182/bloodadvances.2019001258
- Piccin A, Murphy WG, Smith OP. Circulating microparticles: pathophysiology and clinical implications. Blood Rev. 2007;21(3):157–171. doi:10.1016/j.blre.2006.09.001
- Shill DD, Lansford KA, Hempel HK, Call JA, Murrow JR, Jenkins NT. Effect of exercise intensity on circulating microparticles in men and women. Exp Physiol. 2018;103(5):693–700. doi:10.1113/EP086644
- Aharon A, Brenner B. Microparticles and pregnancy complications. Thromb Res. 2011;127(Suppl):S67–71. doi:10.1016/S0049-3848(11)70019-6
- Burton JO, Hamali HA, Singh R, et al. Elevated levels of procoagulant plasma microvesicles in dialysis patients. PLoS One. 2013;8(8):e72663. doi:10.1371/journal.pone.0072663
- Hamali H, Elhussein O, Jamil A, Hussain S, Alshraim M, Alshehri A. Elevated levels of pro‑coagulant microvesicles in children in‑steady state sickle cell disease. J Appl Hematol. 2015;6:115–118. doi:10.4103/1658-5127.165650
- Shet AS, Aras O, Gupta K, et al. Sickle blood contains tissue factor-positive microparticles derived from endothelial cells and monocytes. Blood. 2003;102(7):2678–2683. doi:10.1182/blood-2003-03-0693
- Nantakomol D, Dondorp AM, Krudsood S, et al. Circulating red cell-derived microparticles in human malaria. J Infect Dis. 2011;203(5):700–706. doi:10.1093/infdis/jiq104
- Combes V, Taylor TE, Juhan-Vague I, et al. Circulating endothelial microparticles in Malawian children with severe falciparum malaria complicated with coma. JAMA. 2004;291(21):2542–2544. doi:10.1001/jama.291.21.2542-b
- Campos FMF, Franklin BS, Teixeira-Carvalho A, et al. Augmented plasma microparticles during acute Plasmodium vivax infection. Malar J. 2010;9(1):327. doi:10.1186/1475-2875-9-327
- Pankoui Mfonkeu JB, Gouado I, Fotso Kuaté H, et al. Elevated cell-specific microparticles are a biological marker for cerebral dysfunctions in human severe malaria. PLoS One. 2010;5(10):e13415. doi:10.1371/journal.pone.0013415
- Sahu U, Sahoo PK, Kar SK, Mohapatra BN, Ranjit M. Association of TNF level with production of circulating cellular microparticles during clinical manifestation of human cerebral malaria. Hum Immunol. 2013;74(6):713–721. doi:10.1016/j.humimm.2013.02.006
- Faille D, Combes V, Mitchell AJ, et al. Platelet microparticles: a new player in malaria parasite cytoadherence to human brain endothelium. FASEB J. 2009;23(10):3449–3458. doi:10.1096/fj.09-135822
- Francischetti IMB, Seydel KB, Monteiro RQ, et al. Plasmodium falciparum-infected erythrocytes induce tissue factor expression in endothelial cells and support the assembly of multimolecular coagulation complexes. J Thromb Haemost. 2007;5(1):155–165. doi:10.1111/j.1538-7836.2006.02232.x
- Hamali HA, Saboor M, Dobie G, et al. Procoagulant microvesicles in COVID-19 patients: possible modulators of inflammation and prothrombotic tendency. Infect Drug Resist. 2022;15:2359–2368. doi:10.2147/IDR.S355395
- Hamali HA, Mobarki AA, Akhter MS, et al. Elevated levels of procoagulant microvesicles in patients with dengue fever. Future Virol. 2020;15(10):701–706. doi:10.2217/fvl-2020-0202
- Nieuwland R, Berckmans RJ, McGregor S, et al. Cellular origin and procoagulant properties of microparticles in meningococcal sepsis. Blood. 2000;95(3):930–935. doi:10.1182/blood.V95.3.930.003k46_930_935
- Morel O, Toti F, Hugel B, et al. Procoagulant microparticles: disrupting the vascular homeostasis equation? Arterioscler Thromb Vasc Biol. 2006;26(12):2594–2604. doi:10.1161/01.ATV.0000246775.14471.26
- Abid Hussein MN, Böing AN, Sturk A, Hau CM, Nieuwland R. Inhibition of microparticle release triggers endothelial cell apoptosis and detachment. Thromb Haemost. 2007;98(5):1096–1107. doi:10.1160/TH05-04-0231
- Bidot L, Jy W, Bidot CJ, et al. Microparticle-mediated thrombin generation assay: increased activity in patients with recurrent thrombosis. J Thromb Haemost. 2008;6(6):913–919. doi:10.1111/j.1538-7836.2008.02963.x
- Zwaal RFA, Comfurius P, Bevers EM. Surface exposure of phosphatidylserine in pathological cells. Cell Mol Life Sci. 2005;62(9):971–988. doi:10.1007/s00018-005-4527-3
- Berckmans RJ, Nieuwland R, Böing AN, Romijn FP, Hack CE, Sturk A. Cell-derived microparticles circulate in healthy humans and support low grade thrombin generation. Thromb Haemost. 2001;85(4):639–646. doi:10.1055/s-0037-1615646
- Mackman N, Tilley RE, Key NS. Role of the extrinsic pathway of blood coagulation in hemostasis and thrombosis. Arterioscler Thromb Vasc Biol. 2007;27(8):1687–1693. doi:10.1161/ATVBAHA.107.141911
- Giesen PLA, Rauch U, Bohrmann B, et al. Blood-borne tissue factor: another view of thrombosis. Proc Natl Acad Sci U S A. 1999;96(5):2311–2315. doi:10.1073/pnas.96.5.2311
- GASTAT. Riyadh: General Authority for Statistics. Kingdom of Saudi Arabia: Census; 2010.
- World Health Organization. Microscopy for the Detection, Identification and Quantification of Malaria Parasites on Stained Thick and Thin Blood Films in Research Settings. Geneva, Switzerland: World Health Organization; 2015.
- Fransisca L, Kusnanto JH, Satoto TBT, et al. Comparison of rapid diagnostic test Plasmotec Malaria-3, microscopy, and quantitative real-time PCR for diagnoses of plasmodium falciparum and plasmodium vivax infections in mimika regency, Papua, Indonesia. Malar J. 2015;14:103. doi:10.1186/s12936-015-0615-5
- Guervilly C, Bonifay A, Burtey S, et al. Dissemination of extreme levels of extracellular vesicles: tissue factor activity in patients with severe COVID-19. Blood Adv. 2021;5(3):628–634. doi:10.1182/bloodadvances.2020003308
- Combes V, Coltel N, Alibert M, et al. ABCA1 gene deletion protects against cerebral malaria: potential pathogenic role of microparticles in neuropathology. Am J Pathol. 2005;166(1):295–302. doi:10.1016/S0002-9440(10)62253-5
- Combes V, Coltel N, Faille D, Wassmer SC, Grau GE. Cerebral malaria: role of microparticles and platelets in alterations of the blood-brain barrier. Int J Parasitol. 2006;36(5):541–546. doi:10.1016/j.ijpara.2006.02.005
- Mooberry MJ, Bradford R, Hobl EL, Lin FC, Jilma B, Key NS. Procoagulant microparticles promote coagulation in a factor XI-dependent manner in human endotoxemia. J Thromb Haemost. 2016;14(5):1031–1042. doi:10.1111/jth.13285
- Sinauridze EI, Kireev DA, Popenko NY, et al. Platelet microparticle membranes have 50- to 100-fold higher specific procoagulant activity than activated platelets. Thromb Haemost. 2007;97(3):425–434. doi:10.1160/TH06-06-0313
- Wolberg AS, Monroe DM, Roberts HR, Hoffman MR. Tissue factor de-encryption: ionophore treatment induces changes in tissue factor activity by phosphatidylserine-dependent and -independent mechanisms. Blood Coagul Fibrinolysis an Int J Haemost Thromb. 1999;10(4):201–210. doi:10.1097/00001721-199906000-00007
- Hron G, Kollars M, Binder BR, Eichinger S, Kyrle PA. Identification of patients at low risk for recurrent venous thromboembolism by measuring thrombin generation. JAMA. 2006;296(4):397–402. doi:10.1001/jama.296.4.397
- Hrachovinová I, Cambien B, Hafezi-Moghadam A, et al. Interaction of P-selectin and PSGL-1 generates microparticles that correct hemostasis in a mouse model of hemophilia A. Nat Med. 2003;9(8):1020–1025. doi:10.1038/nm899
- Szotowski B, Antoniak S, Poller W, Schultheiss HP, Rauch U. Procoagulant soluble tissue factor is released from endothelial cells in response to inflammatory cytokines. Circ Res. 2005;96(12):1233–1239. doi:10.1161/01.RES.0000171805.24799.fa
- Rosell A, Havervall S, von Meijenfeldt F, et al. Patients with COVID-19 have elevated levels of circulating extracellular vesicle tissue factor activity that is associated with severity and mortality-brief report. Arterioscler Thromb Vasc Biol. 2021;41(2):878–882. doi:10.1161/ATVBAHA.120.315547
- Østerud B, Bjørklid E. The tissue factor pathway in disseminated intravascular coagulation. Semin Thromb Hemost. 2001;27(6):605–617. doi:10.1055/s-2001-18866
- Abbate R, Cioni G, Ricci I, Miranda M, Gori AM. Thrombosis and acute coronary syndrome. Thromb Res. 2012;129(3):235–240. doi:10.1016/j.thromres.2011.12.026
- Moxon CA, Chisala NV, Mzikamanda R, et al. Laboratory evidence of disseminated intravascular coagulation is associated with a fatal outcome in children with cerebral malaria despite an absence of clinically evident thrombosis or bleeding. J Thromb Haemost. 2015;13(9):1653–1664. doi:10.1111/jth.13060
- Liechti ME, Zumsteg V, Hatz CFR, Herren T. Plasmodium falciparum cerebral malaria complicated by disseminated intravascular coagulation and symmetrical peripheral gangrene: case report and review. Eur J Clin Microbiol Infect Dis off Publ Eur Soc Clin Microbiol. 2003;22(9):551–554. doi:10.1007/s10096-003-0984-5
- Nielsen MH, Beck-Nielsen H, Andersen MN, Handberg A. A flow cytometric method for characterization of circulating cell-derived microparticles in plasma. J Extracell Vesicles. 2014;3(1):20795. doi:10.3402/jev.v3.20795
- Herring JM, McMichael MA, Smith SA. Microparticles in health and disease. J Vet Intern Med. 2013;27(5):1020–1033. doi:10.1111/jvim.12128
- Asare R, Opoku-Okrah C, Danquah KO, et al. Assessment of platelet indices and platelet activation markers in children with Plasmodium falciparum malaria. Malar J. 2020;19(1):143. doi:10.1186/s12936-020-03218-4
- Moulin F, Lesage F, Legros A-H, et al. Thrombocytopenia and Plasmodium falciparum malaria in children with different exposures. Arch Dis Child. 2003;88(6):540LP–541. doi:10.1136/adc.88.6.540
- Hamali HA. The mighty role of platelets in immunity, inflammation, cancer and angiogenesis. Majmaah J Heal Sci. 2020;8(1):65–81. doi:10.5455/mjhs.2020.01.008
- Coltel N, Combes V, Wassmer SC, Chimini G, Grau GE. Cell vesiculation and immunopathology: implications in cerebral malaria. Microbes Infect. 2006;8(8):2305–2316. doi:10.1016/j.micinf.2006.04.006
- Chimalizeni Y, Kawaza K, Taylor T, Molyneux M. The platelet count in cerebral malaria, is it useful to the clinician? Am J Trop Med Hyg. 2010;83(1):48–50. doi:10.4269/ajtmh.2010.09-0614
- Brejt JA, Golightly LM. Severe malaria: update on pathophysiology and treatment. Curr Opin Infect Dis. 2019;32(5):413–418. doi:10.1097/QCO.0000000000000584
- Trappenburg MC, van Schilfgaarde M, Marchetti M, et al. Elevated procoagulant microparticles expressing endothelial and platelet markers in essential thrombocythemia. Haematologica. 2009;94(7):911–918. doi:10.3324/haematol.13774
- Erdman LK, Finney CAM, Liles WC, Kain KC. Inflammatory pathways in malaria infection: tLRs share the stage with other components of innate immunity. Mol Biochem Parasitol. 2008;162(2):105–111. doi:10.1016/j.molbiopara.2008.08.006
- Ardoin SP, Shanahan JC, Pisetsky DS. The role of microparticles in inflammation and thrombosis. Scand J Immunol. 2007;66(2–3):159–165. doi:10.1111/j.1365-3083.2007.01984.x
- Verhamme P, Hoylaerts MF. Hemostasis and inflammation: two of a kind? Thromb J. 2009;7:15. doi:10.1186/1477-9560-7-15
- Egorina EM, Sovershaev MA, Hansen J-B. The role of tissue factor in systemic inflammatory response syndrome. Blood Coagul Fibrinolysis an Int J Haemost Thromb. 2011;22(6):451–456. doi:10.1097/MBC.0b013e328346ef3f
- Crews FT, Zou J, Coleman LGJ. Extracellular microvesicles promote microglia-mediated pro-inflammatory responses to ethanol. J Neurosci Res. 2021;99(8):1940–1956. doi:10.1002/jnr.24813
- Mantel P-Y, Hoang AN, Goldowitz I, et al. Malaria-infected erythrocyte-derived microvesicles mediate cellular communication within the parasite population and with the host immune system. Cell Host Microbe. 2013;13(5):521–534. doi:10.1016/j.chom.2013.04.009
- Butenas S. Tissue factor structure and function. Scientifica. 2012;2012:964862. doi:10.6064/2012/964862
- Couper KN, Barnes T, Hafalla JCR, et al. Parasite-derived plasma microparticles contribute significantly to malaria infection-induced inflammation through potent macrophage stimulation. PLoS Pathog. 2010;6(1):e1000744. doi:10.1371/journal.ppat.1000744
- Morel O, Toti F, Hugel B, Freyssinet J-M. Cellular microparticles: a disseminated storage pool of bioactive vascular effectors. Curr Opin Hematol. 2004;11(3):156–164. doi:10.1097/01.moh.0000131441.10020.87
- Pluskota E, Woody NM, Szpak D, et al. Expression, activation, and function of integrin alphaMbeta2 (Mac-1) on neutrophil-derived microparticles. Blood. 2008;112(6):2327–2335. doi:10.1182/blood-2007-12-127183
- Del Conde I, Shrimpton CN, Thiagarajan P, López JA. Tissue-factor-bearing microvesicles arise from lipid rafts and fuse with activated platelets to initiate coagulation. Blood. 2005;106(5):1604–1611. doi:10.1182/blood-2004-03-1095
- Trelka DP, Schneider TG, Reeder JC, Taraschi TF. Evidence for vesicle-mediated trafficking of parasite proteins to the host cell cytosol and erythrocyte surface membrane in Plasmodium falciparum infected erythrocytes. Mol Biochem Parasitol. 2000;106(1):131–145. doi:10.1016/s0166-6851(99)00207-8
- Kimbi HK, Sumbele IUN, Nweboh M, et al. Malaria and haematologic parameters of pupils at different altitudes along the slope of Mount Cameroon: a cross-sectional study. Malar J. 2013;12(1):193. doi:10.1186/1475-2875-12-193
- Francischetti IMB, Seydel KB, Monteiro RQ. Blood coagulation, inflammation, and malaria. Microcirculation. 2008;15(2):81–107. doi:10.1080/10739680701451516