
Abstract
Iron oxide nanoparticles (IONPs) have been employed for hyperthermia treatments, stem cell therapies, cell labeling, and imaging modalities. The biocompatibility and cytotoxic effects of iron oxide nanoparticles when used in biomedical applications, however, are an ongoing concern. Endothelial cells have a critical role in this research dealing with tumors, cardiovascular disease and inflammation. However, there is little information dealing with the biologic effects of IONPs on the endothelial cell. This paper deals with the influence of dextran and citric acid coated IONPs on the behavior and function of human umbilical vein endothelial cells (HUVECs). After exposing endothelial cells to IONPs, dose-dependent effects on HUVECs viability, cytoskeleton and function were determined. Both citric acid and dextran coated particles appeared to be largely internalized by HUVECs through endocytosis and contribute to eventual cell death possibly by apoptosis. Cytoskeletal structures were greatly disrupted, as evidenced by diminished vinculin spots, and disorganized actin fiber and tubulin networks. The capacity of HUVECs to form a vascular network on Matrigel™ diminished after exposure to IONPs. Cell migration/invasion were inhibited significantly even at very low iron concentrations (0.1 mM). The results of this study indicate the great importance of thoroughly understanding nanoparticle-cell interactions, and the potential to exploit this understanding in tumor therapy applications involving IONPs as thermo/chemoembolization agents.
Supplementary data
Figure S1 FT-IR spectra of citric acid.
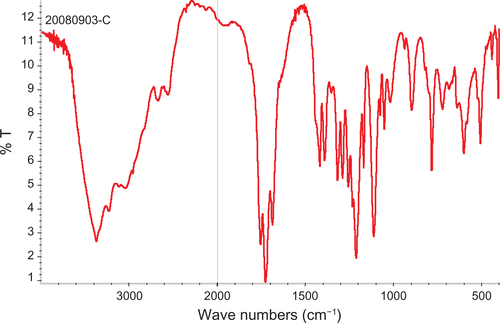
Figure S2 FT-IR spectra of dextran.
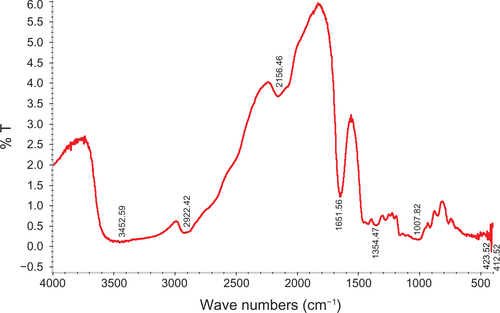
Figure S3 FT-IR spectra of bare iron oxide particles.
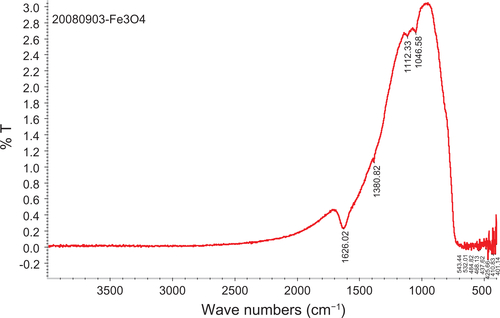
Figure S4 Optical microscopy of HUVECs incubated without or with different IONPs for 24 h. (a) control cell without any particles; (b–d) cells were incubated with citrate-IONP at iron concentration of 0.1, 1 and 10 nM; (e–g) cells were incubated with dextran-IONP at iron concentration of 0.1, 1 and 10 nM.
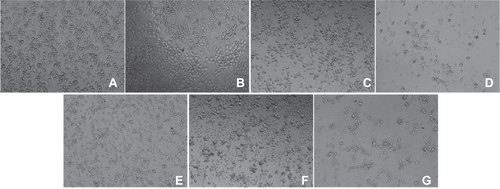
Figure S5 Annexin V kit fluorescent staining of HUVECs incubated without or with different IONPs for 6 h. (a) control cells without any particles; (b, d) with citrate-IONP; (d, e) with dextran-IONP; (b, c) cells were incubated with IONPs at iron concentrations of 0.1 nM. cells were incubated with IONPs at iron concentration of 1 nM. Arrows denote possible apoptotic cells. Bar 100 μm.
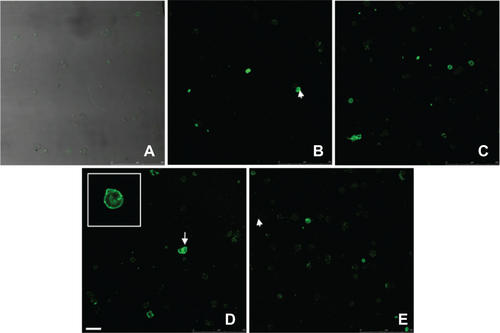
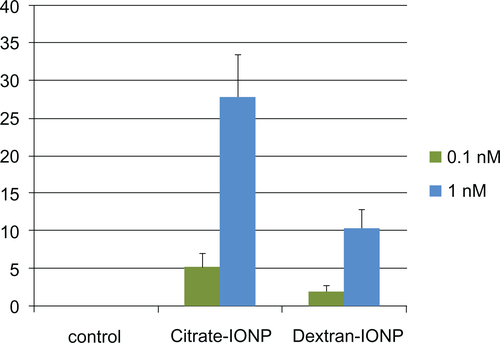
Figure S6 The percentage of apoptotic cells were counted and expressed graphically after 6 h exposure to IONPs.
Acknowledgements
We are sincerely grateful to Dr Yesen Li for his excellent assistance in nanoparticle preparations. We also thank Dr Zhongcai Chen for his kind help with cell experiments and fluorescent staining. Technical support from Emory-Georgia Tech Nanotechnology Center for Personalized and Predictive Oncology and Zhejiang California Nanosystems Institute is acknowledged.
This research was supported by the Chinese National Programs for High Technology Research and Development (Grant No. 2006AA02Z485), Specific Prevention and Control Technology for Major Disease (Grant No. 2008C13027-2), Natural Science Foundation of Zhejiang Province (Grant No. J20060714), Special Foundation for Young Scientists of Zhejiang Province (Grant No. 2009QN009).
Disclosure
The authors have no conflicts of interest that are directly relevant to the content of this work.