
Abstract
Introduction
As a therapeutic antiviral agent, the clinical application of amantadine (AM) is limited by the emergence of drug-resistant viruses. To overcome the drug-resistant viruses and meet the growing demand of clinical diagnosis, the use of biological nanoparticles (NPs) has increased in order to develop novel anti-influenza drugs. The antiviral activity of selenium NPs with low toxicity and excellent activities has attracted increasing attention for biomedical intervention in recent years.
Methods and results
In the present study, surface decoration of selenium NPs by AM (Se@AM) was designed to reverse drug resistance caused by influenza virus infection. Se@ AM with less toxicity remarkably inhibited the ability of H1N1 influenza to infect host cells through suppression of the neuraminidase activity. Moreover, Se@AM could prevent H1N1 from infecting Madin Darby Canine Kidney cell line and causing cell apoptosis supported by DNA fragmentation and chromatin condensation. Furthermore, Se@AM obviously inhibited the generation of reactive oxygen species and activation of phosphorylation of AKT.
Conclusion
These results demonstrate that Se@AM is a potentially efficient antiviral pharmaceutical agent for H1N1 influenza virus.
Introduction
Influenza virus is a segmented RNA virus which is the most highly contagious pathogen worldwide and affects millions of people with influenza each year in seasonal epidemics.Citation1,Citation2 H1N1 influenza virus, which belongs to influenza A type viruses, is a highly infectious respiratory disease.Citation3 This virus was discovered and identified in Mexico and the USA in 2009 and caused 8,768 deaths in 207 countries.Citation4 The mode of spread of H1N1 influenza is as follows: sneezing, cough, and contaminated materials.Citation5 Owing to the mutation of the genome and antigenic shifts in the emerging cross-species infection by avian influenza virus, the influenza virus has a high variability, which may lead to novel influenza strain among humans.Citation6 The influenza infection cycle comprises of several steps: first, the influenza viruses attach to the host cell surface receptor and fuse with the endosomal membrane; second, uncoating of nucleocapsid and multiplication of the genetic material occurs; and finally, the influenza protein and new viron is expressed and released.Citation7,Citation8 Hemagglutinin (HA) and neuraminidase (NA) found on the surface of influenza virus are the most important glycoproteins. HA is a cell-anchoring viral glycoprotein which plays an important role in viral infection by combining sialic acid-containing receptors on host cells and mediating the entry and fusion of the virus.Citation9,Citation10 NA plays an important role in assisting the virus to cleave the linkage between sialic acid and hemagglutinin.Citation11
Existing antiviral drugs approved by the US Food and Drug Administration are NA inhibitors such as oseltamivir/ zanamivir and M2 ion channel inhibitors such as rimantadine/ amantadine (AM).Citation12,Citation13 The M2 proton channel is a critical factor in viral replication; the replication cycle is arrested and infection of the host is halted when proton transport through the channel is inhibited.Citation14,Citation15 The M2 protein is a 97-residue integral membrane protein with a TM domain of 19 residues and a 54-residue cytoplasmic tail, with several point mutations in pore-lining residues of the A/M2 TM domain resulting in resistance to AM.Citation16,Citation17 Because of the emergence of such drug resistance, AM and rimantadine are no longer recommended as common clinical anti-influenza treatments.Citation18,Citation19 Therefore, the antiviral therapies must be promoted in an attempt to control the pandemic influenza A virus.
Nanomaterials with unique chemical and physical properties have emerged as a promising alternative for virus control.Citation20 The design of new antiviral nanodrug should consider strategies to effectively control viral infection and also deal with the cytotoxicity related to the exposure of biological materials.Citation21,Citation22 According to Tao et al, consensus M2e peptide was combined with gold nanoparticles (NPs) against H1N1 influenza A viruses.Citation23 According to Ye et al, graphene oxide is a promising antiviral agent due to its unique properties.Citation24 According to Vonnemann et al, different sizes of polyvalent NPs inhibited virus.Citation25 Meanwhile, the potential of NPs against viral infections through immunization was reported by Sokolova et al.Citation26 Wang et al reported that the SiO2-layered double hydroxide NPs enhance the response of hepatitis B virus DNA vaccine.Citation27 According to Wang et al, polyoxometalate with a broad spectrum may be a new type of antiviral agent.Citation28 According to Barras et al, carbon nanodots have high-efficiency functionality as entry inhibitors in the early stage of virus infection.Citation29 According to Khanal et al, phenylboronic-acid-modified NPs possess potential antiviral therapeutic application.Citation30 Among them, selenium NPs (SeNPs) with their unique antimicrobial activities have attracted considerable attention.Citation31,Citation32 Se is an essential nutritional trace element with the ability to regulate cellular redox homeostasis.Citation33–Citation35 The deficiency of Se could increase the susceptibility to infections, including respiratory virus and hepatitis B virus infections.Citation36,Citation37 In this study, we present novel SeNPs that can inhibit the ability of H1N1 influenza virus to infect host cell. We look forward to verifying that AM-modified SeNPs (Se@AM) have excellent antiviral activity. Reactive oxygen species (ROS) plays an important role in many physiological processes; oxidative stress is explicated between consumption of ROS and cellular defense mechanisms.Citation38,Citation39 The imbalance of redox is associated with many pathologies, such as skin disease, diabetes, cancer, Leigh syndrome and other diseases.Citation40 This study was performed to investigate how Se@AM inhibits H1N1 influenza virus-induced host cells apoptosis through ROS-mediated signaling pathways.
Materials and methods
Materials
The Madin–Darby Canine Kidney (MDCK) cells were obtained from ATCC® CCL-34TM. H1N1 influenza virus was provided by the Virus Laboratory, Guangzhou Women and Children’s Medical Center, Guangzhou Medical University. Fetal bovine serum and DMEM were purchased from Thermo Fisher Scientific, Waltham, MA, USA. Na2SeO3, vitamin C, AM, MTT, propidium iodide, 4′,6-diamidino-2-phenylindole (DAPI), and 2′,7′-dichlorofluorescin (DCF)-diacetate were purchased from Sigma-Aldrich Co. (St Louis, MO, USA). AKT and caspase-3 antibodies were purchased from Cell Signaling Technology (Boston, MA, USA).
Preparation of Se@AM
SeNPs were synthesized as follows: 0.25 mL stock solution (0.1 M) of Na2SeO3 was gradually added to 2 mL stock solution (50 mM) of vitamin C. Then, 0.8 mL of 1 µM AM was added to the SeNPs solution. We detected the excess AM, vitamin and Na2SeO3 by dialysis for 24 hours. Se@AM NPs were sonicated and then filtered through 0.2 µm size pores. The concentration of SeNPs and AM was measured by inductively coupled plasma atomic emission spectroscopy.
Characterization of Se@AM
The morphology of Se@AM NPs was characterized by transmission electron microscopy (TEM, H-7650). Elemental composition of Se@AM was determined by energy dispersive X-ray spectroscopy (EDX) (EX-250 system; Horiba, Kyoto, Japan). Zeta potential and size distribution of Se@AM were monitored by Malvern Zetasizer (Malvern Instruments Limited, Manchester, UK) software. Fourier-transform infrared spectroscopy samples were recorded using the potassium bromide-disk method (Equinox 55 IR spectrometer). X-ray photoelectron spectroscopy (XPS) measurement was carried out on ESCALab 250 spectrometer.Citation41
Cell viability by MTT assay
Cytotoxicity of Se@AM NPs was performed as previously described.Citation42 H1N1 influenza virus was incubated for 2 h and then rinsed to remove the viruses that were not internalized. The indicated concentrations of AM with or without SeNPs were added to MDCK cells for 24 h. Then, 20 µL/well MTT was added and incubated for 5 h. The formazan crystals were dissolved by adding dimethyl sulfoxide (150 µL/well) and measured at 570 nm.
TEM of Se@AM-treated H1N1
H1N1 influenza virus was treated with Se@AM, and then attached to the carbon-coated collodion grid. The grids were examined by TEM.Citation43
The NA inhibition assay by Se@AM
MDCK cells were infected with H1N1 influenza virus for 2 h. After 24 h, the cell culture supernatants were harvested. Influenza virus NA activity was determined by quantifying the fluorescent product resulting from the cleavage of the substrate 4-methylumbelliferyl-α-D-N-acetylneuraminic acid sodium salt hydrate solution by NA.Citation44 The reaction mixture consisted of the tested compounds, after incubation for 30 min at 37°C. The fluorescence of the mixture was recorded at excitation wavelength of 360 nm and emission at 460 nm.
Detection of mitochondrial membrane potential (ΔΨm)
The fluorescence intensity from JC-1 monomers was used to estimate the status of ΔΨm in MDCK cells exposed to Se@oseltamivir (OTV), as previously described.Citation45 Cells were trypsinized and resuspended with 10 µg/mL of JC-1. The MDCK cells were then harvested and analyzed by flow cytometry.
Annexin-V-FLUOS staining assay
Plasma membrane alterations in MDCK cells treated with Se@AM were detected as previously described.Citation46 Briefly, MDCK cells were treated with Se@AM for 24 h and stained with annexin-V-FLUOS, then washed with PBS 3 times, and observed by flow cytometric analysis.
Terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick-end labeling (TUNEL)-DAPI co-staining assay
MDCK cells were observed under fluorescence microscope with TUNEL and incubated with DAPI for nuclear staining, as previously described.Citation47
Caspase-3 activity
The caspase-3 activity was detected under fluorescence microscope with the excitation at 380 nm and emission at 460 nm wavelengths, as previously described.Citation48
TEM analysis of thin sections of MDCK cells
TEM analysis of MDCK cells was performed in situ as previously described.Citation49
Determination of ROS generation
The ROS level was monitored by analyzing the fluorescence intensity of DCF with excitation at 500 nm and emission at 529 nm wavelengths.Citation50
Western blotting analysis
The total proteins were obtained after MDCK cells were treated with Se@AM and incubated with lysis buffer. BCA assay was used to quantify the protein concentration. The bolts were developed with enhanced chemiluminescence reagent that examines the target proteins on the X-ray film.Citation51
Statistical analysis
All the data are presented as mean ± SD. Differences between the 2 groups were evaluated using 2-tailed Student’s t-test. One-way analysis of variance was used in multiple-group comparisons. Difference with P<0.05 or P<0.01 was considered statistically significant.
Results and discussion
Preparation of Se@AM
Functionalized Se@AM was synthesized. SeNPs were modified with AM to form more compact and stable NPs (). Light images of AM, SeNPs, and Se@AM are shown in . As shown in , the Tyndall effect of Se@AM indicated that Se@AM NPs were synthesized. The morphology of Se@AM presented uniformly spherical particles as shown in . As shown in , EDX indicated the signal of C (20%) and N (8%) from AM, while Se atoms were 72%. The size of Se@AM was decreased from 200 to 70 nm, which indicated much smaller size as shown in . The zeta potential of Se@AM (−36.9 mv) was lower than SeNPs (−23.5 mv), demonstrating the higher stability of Se@AM than SeNPs as shown in . The size distribution of Se@AM in revealed that Se@AM was stable for 30 days. Se@AM was further characterized to confirm the chemical binding of AM to the surface of SeNPs. The Fourier transform infrared (FTIR) spectra of unmodified AM, SeNPs, and Se@AM are shown in . The FTIR spectrum of Se@AM resembles that of AM, giving clear evidence that AM ligand forms a part of the nanocomposite. AM displays IR absorbance peaks at 2,903, 1,264, and 739 cm−1 corresponding to –CH2, –NH2, and C–N. The absence of these peaks in Se@AM indicated the formation of Se@AM. The XPS spectra were also recorded to examine the interaction between AM and SeNPs. As shown in , the N 1s peak in the spectrum of Se@AM further confirmed that AM has been successfully conjugated to the SeNPs.
Figure 1 Light images of AM, SeNPs, and Se@AM.
Notes: (A) The color change of AM, SeNPs, and Se@AM. (B and C) Tyndall effect of Se@AM.
Abbreviations: AM, amantadine; Se@AM, AM-modified SeNPs; SeNPs, selenium nanoparticles.

Figure 2 Characterizations of SeNPs and Se@AM.
Notes: (A) TEM images of SeNPs and Se@AM. (B) EDX analysis of Se@AM. (C and D) Size distribution of SeNPs and Se@AM. (E) Zeta potentials of SeNPs and Se@AM. (F) Stability of Se@AM in aqueous solutions.
Abbreviations: AM, amantadine; EDX, energy dispersive X-ray spectroscopy; Se@AM, AM-modified SeNPs; SeNPs, selenium nanoparticles; TEM, transmission electron microscopy.
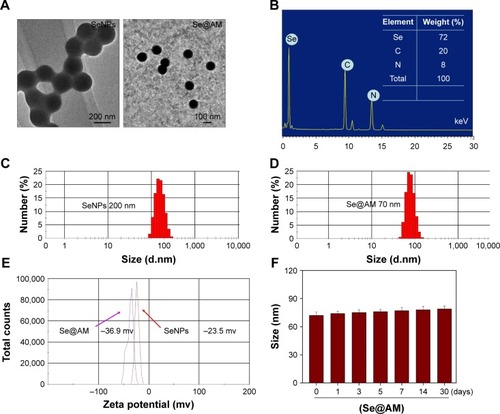
Figure 3 The FTIR spectra and XPS of Se@AM.
Notes: All IR spectra were acquired in the form of potassium bromide plates. (A) FTIR spectrums. (B) XPS detection.
Abbreviations: AM, amantadine; FTIR, Fourier transform infrared; Se@AM, AM-modified SeNPs; SeNPs, selenium nanoparticles; XPS, X-ray photoelectron spectroscopy.
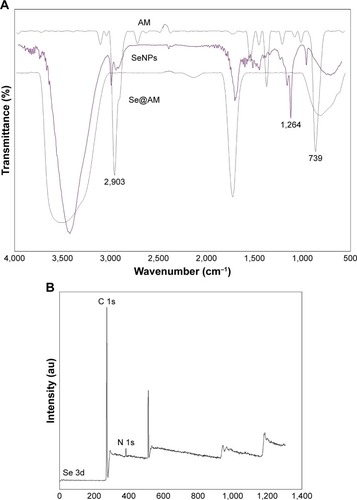
In vitro antiviral activity of Se@AM
MDCK cells treated with H1N1 influenza virus showed cell viability of 32.34%. AM and SeNPs increased the cell viability to 53.23% and 58.87%, as shown in , but the cell viability of Se@AM was significantly increased to 79.26%. The results indicate that the antiviral effect of Se@ AM was superior to that of free SeNPs and AM. As shown in , cells treated with H1N1 influenza virus showed cytoplasmic shrinkage, loss of cell-to-cell contract, and reduction in cells numbers. The MDCK cell morphology changed only slightly by co-treatment with Se@AM and appeared healthy in shape. Synergy was evaluated by calculation of in vitro fractional inhibitory concentration-index values: minimum inhibitory concentration (MIC) of Se in Se@AM (125 µM); MIC of drug B combination concentration of AM in Se@AM (0.1 µM); MIC of drug A alone corresponded to free SeNPs (1 mM); MIC of drug B alone corresponded to free AM (0.4 µM). Fractional inhibitory concentration (FIC) was calculated as (MIC of drug A combination/MIC of drug A alone) + (MIC of drug B combination/MIC of drug B alone) = 125 µM/1 mM + 0.1 µM/0.4 µM = 0.375. FIC was 0.375, below 0.5, indicating synergy. In this study, the FIC index was basically interpreted as follows: FIC <0.5, synergy; FIC between 0.5 and 2, indifference; FIC >2, antagonism. The results suggest that Se@AM effectively inhibited the proliferation of H1N1 influenza virus (). The titer of H1N1 was detected by Reed-Muench assay (TCID=103.5).
Table 1 Titer of H1N1
Figure 4 Effects of Se@AM on the growth of H1N1 infection of MDCK cells by MTT assay.
Notes: (A) Antiviral activity of Se@AM. Free concentration of SeNPs was 1 mM and AM 0.4 µm. AM (0.1 µM) and SeNPs (0.125 µM) were loaded onto the Se@AM. (B) Morphological changes in H1N1-infected MDCK cells were observed by phase-contrast microscopy. Bars with different characters are statistically different at *p<0.05 or **p<0.01 level.
Abbreviations: AM, amantadine; MDCK, Madin–Darby Canine Kidney; Se@AM, AM-modified SeNPs; SeNPs, selenium nanoparticles.
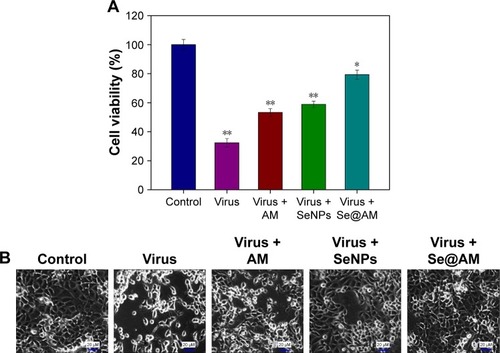
H1N1 morphologic changes
The group of H1N1 virus showed typical elliptical or spherically shaped normal H1N1 virus containing virus matrix and capsid (). After interaction of Se@AM with H1N1 virus for 12 and 24 h, the parts of H1N1 viral edges were lost and the viral morphology was destroyed (). The results demonstrate that Se@AM could directly interact with virus particles and lead to disruption of viral function.
Figure 5 Morphologic abnormalities in Se@AM-treated H1N1.
Notes: (A) The HR-TEM negative staining. (B and C) H1N1 virus interacting with Se@AM at 12 and 24 h. White arrows indicate the Se@AM nanoparticles.
Abbreviations: AM, amantadine; HR, high resolution; Se@AM, AM-modified SeNPs; SeNPs, selenium nanoparticles; TEM, transmission electron microscopy.
Identifying NA as the potential target of Se@AM
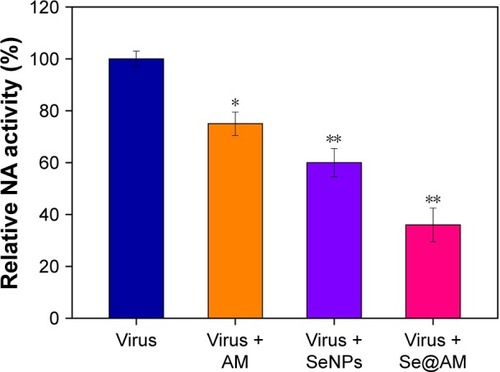
Surface glycoprotein NA plays an important role in the entry of virus. Untreated H1N1 (100%) or Se@AM-treated H1N1were used to measure the NA enzymatic activity. Compared with AM (75%) and SeNPs (60%), Se@AM (36%) inhibited obvious enzymatic activity of influenza H1N1 viral NA, indicating that the Se@AM NPs had inhibitory effect on NA activity (). The anti-influenza mechanism of Se@AM is postulated as: the Se@AM nanoparticles bind tightly to the NA protein, thus forbidding the attachment of H1N1 virus to MDCK cells.
Figure 6 Inhibition of NA activity by Se@AM in H1N1 infection of cells.
Notes: The NA inhibition assay was performed using a microplate reader. Bars with different characters are statistically different at *p<0.05 or **p<0.01 level.
Abbreviations: AM, amantadine; NA, neuraminidase; Se@AM, AM-modified SeNPs; SeNPs, selenium nanoparticles.
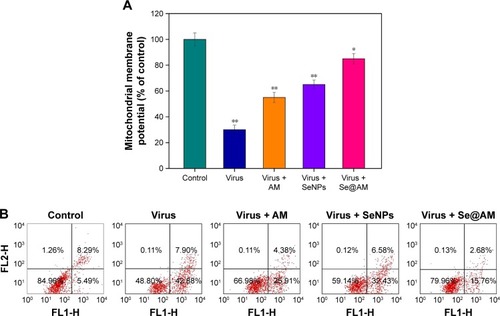
Depletion of mitochondrial membrane potential (ΔΨm) and translocation of phosphatidylserine induced by Se@OTV
MDCK cells were treated with mitochondrial-selective JC-1 dye. As shown in , treatments of MDCK cells with H1N1 influenza virus (30%) resulted in elevation of mitochondrial depolarization and dysfunction. Compared with AM (55%) and SeNPs (65%), when MDCK cells were exposed to Se@AM, the percentage of mitochondrial membrane potential significantly increased to 85%. These results demonstrate that Se@AM inhibited H1N1 influenza virus by apoptosis in MDCK cells by inducing mitochondrial dysfunction. The percentage of living cells treatment with H1N1 influenza virus was 48.80% as shown in . Compared with AM (66.98%) and SeNPs (59.14%), when MDCK cells were exposed to Se@AM, the percentage of MDCK cells treated with Se@AM revealed a decrease in cell number (79.96%). The results demonstrate that Se@ AM restrained H1N1 virus infection of MDCK cells mainly through inhibiting apoptosis.
Figure 7 Depletion of mitochondrial membrane potential and translocation of phosphatidylserine induced by virus, AM, SeNPs, and Se@AM.
Notes: (A) Mitochondrial membrane potential of H1N1 infection of MDCK cells exposed to virus, AM, SeNPs, and Se@AM. (B) Translocation of phosphatidylserine induced by virus, AM, SeNPs and Se@AM in H1N1 infection of MDCK cells as determined by annexin-V-FLUOS labeling assay. Bars with different characters are statistically different at P<0.05 (*) or P<0.01 (**) level.
Abbreviations: AM, amantadine; MDCK, Madin–Darby Canine Kidney; NA, neuraminidase; Se@AM, AM-modified SeNPs; SeNPs, selenium nanoparticles.
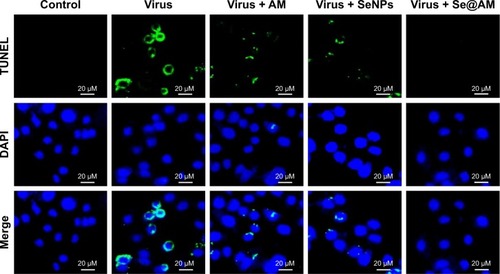
Inhibition of H1N1 infection
As shown in , MDCK cells exhibited apoptotic features by H1N1 influenza virus with DNA fragmentation and nuclear condensation. Co-treatment with Se@AM remarkably prevented the H1N1 influenza virus-induced changes in nuclear morphology. The results indicate that Se@AM rescues the apoptosis of MDCK cells by H1N1 influenza virus.
Figure 8 Se@AM inhibited apoptosis in H1N1 infection of MDCK cells.
Notes: DNA fragmentation and nuclear condensation as detected by TUNEL-DAPI co-staining assay. All results are representative of 3 independent experiments.
Abbreviations: AM, amantadine; DAPI, 4′,6-diamidino-2-phenylindole; MDCK, Madin–Darby Canine Kidney; Se@AM, AM-modified SeNPs; SeNPs, selenium nanoparticles; TUNEL, terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick-end labeling.
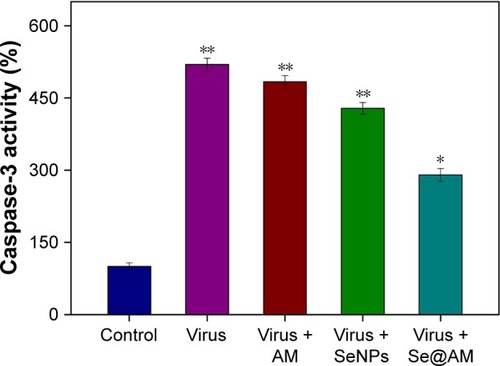
Inhibition of caspase-3 activation
Caspase-3 activity was used to determine cell apoptosis. As shown in , treatment of MDCK cells with H1N1 obviously increased the caspase-3 activity to 519.7%. The caspase-3 activity by AM and SeNPs was 483.6% and 428.3%, respectively. However, Se@AM significantly decreased the caspase-3 activity to 290.2%. The results show that the Se@AM inhibits H1N1 influenza virus activity.
Figure 9 Inhibition of caspase-3 activity by Se@AM in H1N1 infection of cells.
Notes: Cells were treated with Se@AM and caspase-3 activity detected by synthetic fluorogenic substrate. Bars with different characters are statistically different at *p<0.05 or **p<0.01 level.
Abbreviations: AM, amantadine; Se@AM, AM-modified SeNPs; SeNPs, selenium nano particles.
TEM image of thin sections of cells
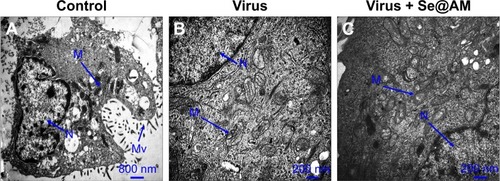
Mitochondria and microvilli were observed, with no morphological changes in untreated cells as shown in . When MDCK cells were treated with H1N1 influenza virus, TEM image indicated distorted organelles and condensed chromatin, which indicated apoptosis of MDCK cells (). As shown in , the mitochondria of MDCK cells recovered their shape after treatment with Se@AM, which could reveal that H1N1 influenza virus induced apoptosis.
Figure 10 TEM images of thin sections of MDCK cells treated with different groups.
Notes: (A) Control, (B) cells treated with virus, and (C) cells treated with virus + Se@AM at 24 h.
Abbreviations: AM, amantadine; MDCK, Madin–Darby Canine Kidney; Se@AM, AM-modified SeNPs; SeNPs, selenium nanoparticles; TEM, transmission electron microscopy; N, nucleus; M, mitochondria; Mv, microvilli.
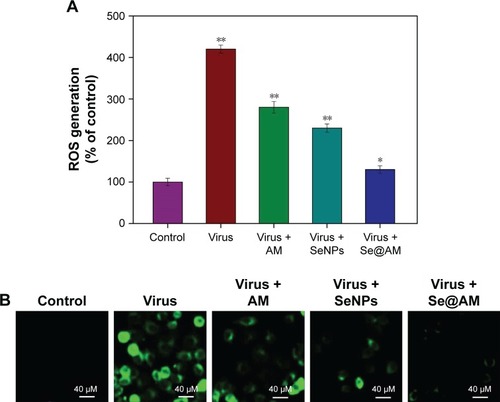
Inhibition of ROS generation by Se@AM
The ROS generation was monitored through DCF assay to indicate the action mechanisms of Se@AM. As shown in , the ROS generation by H1N1 influenza virus was increased to 420%. AM and SeNPs slightly inhibited the ROS generation to 280% and 230%. However, Se@AM remarkably decreased the rate of ROS generation (130%). The fluorescent intensity of DCF was found by H1N1 influenza virus as shown in . The fluorescent intensity of DCF in MDCK cells treated with H1N1 influenza virus was much stronger than in those treated with Se@AM. These results show that Se@AM could downregulate the level of ROS in the antiviral action.
Figure 11 ROS overproduction induced by Se@AM in H1N1 infection of MDCK cells.
Notes: (A) ROS levels were detected by DCF fluorescence intensity. (B) H1N1 infection of MDCK cells preincubated with 10 µM DCF for 30 min and then treated with Se@AM. Bars with different characters are statistically different at P<0.05 (*) or P<0.01 (**) level.
Abbreviations: AM, amantadine; DCF, 2′,7′-dichlorofluorescein; MDCK, Madin–Darby Canine Kidney; ROS, reactive oxygen species; Se@AM, AM-modified SeNPs; SeNPs, selenium nanoparticles.
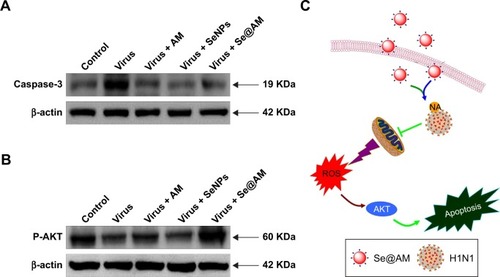
ROS activates apoptotic signaling pathways by Se@AM
The over-generation of ROS could cause DNA damage and affect the regulation of apoptosis signaling pathways. Compared with AM and SeNPs, treatments of MDCK cells with Se@AM remarkably inhibited the expression levels of caspase-3, as shown in . Meanwhile, the level of AKT was significantly increased by Se@AM (). Taken together, Se@AM depressed H1N1 influenza virus-induced host cell apoptosis by ROS-mediated AKT signaling pathways ().
Figure 12 Intracellular apoptotic signaling pathways by Se@AM in H1N1 infection of MDCK cells.
Notes: (A) The protein expression of caspase-3. (B) Phosphorylation status expression levels of AKT pathways. (C) The main signaling pathway of ROS-mediated AKT signaling pathways. β-actin was used as loading control.
Abbreviations: AM, amantadine; MDCK, Madin–Darby Canine Kidney; ROS, reactive oxygen species; Se@AM, AM-modified SeNPs; SeNPs, selenium nanoparticles; NA, neuraminidase.
Scheme 1 Synthetic route toward Se@AM.
Abbreviations: AM, amantadine; Se@AM, AM-modified SeNPs; SeNPs, selenium nano particles.
Conclusion
In summary, an efficient chemical method for AM surface-modified SeNPs was described in this study. Se@AM with lower toxicity exhibits enhanced abilities to prevent H1N1 influenza virus infection. The NA activity of H1N1 influenza virus was inhibited by Se@AM. The molecular mechanisms demonstrated that Se@AM inhibited caspase-3 activity and mediated apoptosis through ROS generation. Furthermore, Se@AM inhibited MDCK cell apoptosis by decreasing the level of ROS to trigger AKT pathways. Therefore, Se@AM might provide a promising Se subtype with antiviral properties against H1N1 influenza virus.
Acknowledgments
This work was supported by the Technology Planning Project of Guangzhou (201804010183), Medical Scientific Research Foundation of Guangdong Province (A2018306), Medical Scientific Research Foundation of Guangdong Province (A2018289), The Technology Planning Project of Guangdong Province (2014A020212697), the Technology Planning Project of Guangzhou (201607010120), Guangzhou medical health science and technology project (20181A010022), Science and Technology Project of Guangzhou (201707010014), and National Natural Science Foundation of China (81600399).
Disclosure
The authors report no conflicts of interest in this work.
References
- LiYLinZZhaoMSilver nanoparticle based codelivery of oseltamivir to inhibit the activity of the H1N1 influenza virus through ROS-mediated signaling pathwaysACS Appl Mater Interfaces2016837243852439327588566
- WangCZHanHHTangXYSialylglycan-assembled supra-dots for ratiometric probing and blocking of human-infecting influenza virusesACS Appl Mater Interfaces2017930251642517028703004
- YangMSunderlandKMaoCVirus-derived peptides for clinical applicationsChem Rev201711715103771040228723101
- LiYHLinZFZhaoMQReversal of H1N1 influenza virus-induced apoptosis by silver nanoparticles functionalized with amantadineRSC Adv20166928967989686
- LiYLinZGuoMInhibitory activity of selenium nanoparticles functionalized with oseltamivir on H1N1 influenza virusInt J Nanomedicine2017125733574328848350
- JungJHParkBHOhSJChoiGSeoTSIntegration of reverse transcriptase loop-mediated isothermal amplification with an immunochromatographic strip on a centrifugal microdevice for influenza A virus identificationLab Chip201515371872525426967
- LovedayEKDiederichSPasickJJeanFHuman microRNA-24 modulates highly pathogenic avian-origin H5N1 influenza A virus infection in A549 cells by targeting secretory pathway furinJ Gen Virol201596Pt 1303925234642
- LondriganSLShortKRMaJInfection of mouse macrophages by seasonal influenza viruses can be restricted at the level of virus entry and at a late stage in the virus life cycleJ Virol20158924123191232926423941
- LiuYZhangLWeiWColorimetric detection of influenza A virus using antibody-functionalized gold nanoparticlesAnalyst2015140123989399525899840
- HaiWGodaTTakeuchiHSpecific recognition of human influenza virus with PEDOT bearing sialic acid-terminated trisaccharidesACS Appl Mater Interfaces2017916141621417028379685
- LinZLiYGuoMThe inhibition of H1N1 influenza virus-induced apoptosis by silver nanoparticles functionalized with zanamivirRSC Adv201772742750
- LinZLiYGuoMInhibition of H1N1 influenza virus by selenium nanoparticles loaded with zanamivir through p38 and JNK signaling pathwaysRSC Adv20177563529035296
- ChenQGuoYInfluenza viral hemagglutinin peptide inhibits influenza viral entry by shielding the host receptorACS Infect Dis20162318719327623031
- GleedMLBusathDDWhy bound amantadine fails to inhibit proton conductance according to simulations of the drug-resistant influenza A M2 (S31N)J Phys Chem B201511931225123125426702
- GleedMLIoannidisHKolocourisABusathDDResistance-mutation (N31) effects on drug orientation and channel hydration in amantadine-bound influenza A M2J Phys Chem B201511935115481155926268449
- Rey-CarrizoMBarniol-XicotaMMaCEasily accessible polycyclic amines that inhibit the wild-type and amantadine-resistant mutants of the M2 channel of influenza A virusJ Med Chem201457135738574724941437
- WuYCanturkBJoHFlipping in the pore: discovery of dual inhibitors that bind in different orientations to the wild-type versus the amantadine-resistant 531N mutant of the influenza A virus M2 proton channelJ Am Chem Soc201413652179871799525470189
- DurrantMGEggettDLBusathDDInvestigation of a recent rise of dual amantadine-resistance mutations in the influenza A M2 sequenceBMC Genet201516Suppl 2S325953496
- Van NguyenHNguyenHTLeLTInvestigation of the free energy profiles of amantadine and rimantadine in the AM2 binding pocketEur Biophys J2016451637026391902
- de SouzaESilvaJMHanchukTDSantosMIKobargJBajgelmanMCCardosoMBViral inhibition mechanism mediated by surface- modified silica nanoparticlesACS Appl Mater Interfaces2016826165641657227284685
- GustafssonSMihranyanAStrategies for tailoring the pore-size distribution of virus retention filter papersACS Appl Mater Interfaces2016822137591376727144657
- LiYHLinZFXuTTDelivery of VP1 siRNA to inhibit the EV71 virus using functionalized silver nanoparticles through ROS-mediated signaling pathwaysRSC Adv20177314531463
- TaoWHurstBLShakyaAKConsensus M2e peptide conjugated to gold nanoparticles confers protection against H1N1, H3N2 and H5N1 influenza A virusesAntiviral Res2017141627228161578
- YeSShaoKLiZAntiviral activity of graphene oxide: how sharp edged structure and charge matterACS Appl Mater Interfaces2015738215712157926370151
- VonnemannJSiebenCWolffCVirus inhibition induced by polyvalent nanoparticles of different sizesNanoscale2014642353236024430614
- SokolovaVWestendorfAMBuerJUberlaKEppleMThe potential of nanoparticles for the immunization against viral infectionsJ Mater Chem B201532447674779
- WangJZhuRGaoBThe enhanced immune response of hepatitis B virus DNA vaccine using SiO2@LDH nanoparticles as an adjuvantBiomaterials201435146647824099705
- WangJLiuYXuKBroad-spectrum antiviral property of polyoxometalate localized on a cell surfaceACS Appl Mater Interfaces20146129785978924878685
- BarrasAPagneuxQSaneFHigh efficiency of functional carbon nanodots as entry inhibitors of herpes simplex virus type 1ACS Appl Mater Interfaces20168149004901327015417
- KhanalMVausselinTBarrasAPhenylboronic-acid-modified nanoparticles: potential antiviral therapeuticsACS Appl Mater Interfaces2013523124881249824180242
- RamyaSShanmugasundaramTBalagurunathanRBiomedical potential of actinobacterially synthesized selenium nanoparticles with special reference to anti-biofilm, anti-oxidant, wound healing, cytotoxic and anti-viral activitiesJ Trace Elem Med Biol201532303926302909
- StevanovicMFilipovicNDjurdjevicJLukicMMilenkovicMBoccacciniA45S5Bioglass (R)-based scaffolds coated with selenium nanoparticles or with poly(lactide-co-glycolide)/selenium particles: processing, evaluation and antibacterial activityColloids Sur B Biointerfaces2015132208215
- YangYXieQZhaoZNFunctionalized selenium nanosystem as radiation sensitizer of I-125 seeds for precise cancer therapyACS Appl Mater Interfaces2017931258572586928718286
- JiangWFuYYangFGracilaria lemaneiformis polysaccharide as integrin-targeting surface decorator of selenium nanoparticles to achieve enhanced anticancer efficacyACS Appl Mater Interfaces2014616137381374825073123
- HuangYLuoYZhengWChenTRational design of cancer-targeted BSA protein nanoparticles as radiosensitizer to overcome cancer radioresistanceACS Appl Mater Interfaces2014621192171922825314331
- YuLSunLNanYZhuLYProtection from H1N1 influenza virus infections in mice by supplementation with selenium: a comparison with selenium-deficient miceBiol Trace Elem Res20111411–325426120490710
- ChengZZhiXSunGSodium selenite suppresses hepatitis B virus transcription and replication in human hepatoma cell linesJ Med Virol201688465366326331371
- LiYLinZZhaoMMultifunctional selenium nanoparticles as carriers of HSP70 siRNA to induce apoptosis of HepG2 cellsInt J Nanomedicine2016113065307627462151
- LiYGuoMLinZPolyethylenimine-functionalized silver nanoparticle-based co-delivery of paclitaxel to induce HepG2 cell apoptosisInt J Nanomedicine2016116693670227994465
- ZhuBLiYLinZSilver nanoparticles induce HePG-2 cells apoptosis through ROS-mediated signaling pathwaysNanoscale Res Lett20161119826909778
- LiuWLiXWongYSSelenium nanoparticles as a carrier of 5-fluorouracil to achieve anticancer synergismACS Nano2012686578659122823110
- LiYLiXWongYSThe reversal of cisplatin-induced nephrotoxicity by selenium nanoparticles functionalized with 11-mercapto-1-undecanol by inhibition of ROS-mediated apoptosisBiomaterials201132349068907621864903
- XiangDXChenQPangLZhengCLInhibitory effects of silver nanoparticles on H1N1 influenza A virus in vitroJ Virol Methods20111781–213714221945220
- XiaoSSiLTianZPentacyclic triterpenes grafted on CD cores to interfere with influenza virus entry: a dramatic multivalent effectBiomaterials201678748526686050
- ZhangYLiXHuangZZhengWFanCChenTEnhancement of cell permeabilization apoptosis-inducing activity of selenium nanoparticles by ATP surface decorationNanomedicine201391748422542821
- WuHLiXLiuWSurface decoration of selenium nanoparticles by mushroom polysaccharides-protein complexes to achieve enhanced cellular uptake and antiproliferative activityJ Mater Chem2012221996029610
- LiYLiXZhengWFanCZhangYChenTFunctionalized selenium nanoparticles with nephroprotective activity, the important roles of ROS-mediated signaling pathwaysJ Mater Chem2012221996029610
- LiXMaLZhengWChenTInhibition of islet amyloid polypeptide fibril formation by selenium-containing phycocyanin and prevention of beta cell apoptosisBiomaterials201435308596860425034964
- LiJMWangYYZhaoMXMultifunctional QD-based co-delivery of siRNA and doxorubicin to HeLa cells for reversal of multidrug resistance and real-time trackingBiomaterials20123392780279022243797
- FuXYangYLiXRGD peptide-conjugated selenium nanoparticles: antiangiogenesis by suppressing VEGF-VEGFR2-ERK/AKT pathwayNanomedicine20161261627163926961468
- XiaYLinZLiYTargeted delivery of siRNA using RGDfC-conjugated functionalized selenium nanoparticles for anticancer therapyJ Mater Chem B201753369416952