Abstract
Despite remarkable progress in the development of both viral and nonviral gene delivery vectors for airway disease treatment, poor gene transfer efficiency to the airway epithelium is a major obstacle in clinical application. To take advantage of the unique features of viral and nonviral vectors, we have developed complexes of adenovirus vector and anionic liposomes (AL-Ad5) by the calcium-induced phase change method. In the current study, based on the fact that there are overexpressed folate receptors on the surface of airway epithelia, we further modified the AL-Ad5 complexes with folate (F-AL-Ad5) to improve the transduction ability of Ad5 in airway epithelia. The transduction efficiencies of the obtained F-AL-Ad5 and AL-Ad5 complexes were assessed in primary-cultured airway epithelia in vitro. Our results indicated that compared with naked adenovirus vector, both AL-Ad5 and F-AL-Ad5 could significantly enhance the gene transduction efficiency of adenovirus vector in primary-cultured airway epithelial cells. Moreover, the enhancement mediated by F-AL-Ad5 was more dramatic than that by AL-Ad5. These results suggested that F-AL-Ad5 may be a useful strategy to deliver therapeutic genes to the airway epithelia and is promising in clinical application.
Introduction
Many studies have developed adenovirus-mediated gene transfer to cultured epithelial cells and various in vivo airway models,Citation1–Citation4 but low efficiency of gene transfer to the airway epithelium is a major obstacle to clinical application.Citation5 It has been well documented that the uptake of adenovirus vector is dependent on the expression of coxsackievirus–adenovirus receptor (CAR) on the cell surface.Citation6,Citation7 However, Walters’s studyCitation8 demonstrated that CAR was not present on the apical surface of airway epithelium but located in the basolateral membrane. Thus, airway epithelial cell was a difficult target for gene therapy. To solve this key problem, high doses of virus and long periods of physical contact between the vector and targeted airway epithelial cells were commonly required for efficient gene transfer. But high doses of virus can initiate effective cellular and humoral immune responses.Citation9,Citation10 Moreover, the airway epithelium could respond to a range of stimuli by producing biologically active mediators such as cytokines, lipid mediators, peptide products, reactive oxygen species, and so on, which function to recruit circulating leukocytes to the airway, modulate airway tone, regulate secretions, and promote antimicrobial and antiviral activity.Citation11 Furthermore, the extended vector contact with the airway epithelium was prevented by natural clearance mechanisms as well as mucous and soluble factors in the airway surface fluid.Citation12
On the other hand, many studies have focused on the gene delivery system based on nonviral vectors. Among those nonviral vectors, cationic liposomes have been most widely studied and proved to be able to increase intracellular delivery potency and reduce inflammatory risk compared with viral vectors. However, a critical factor of transfection barriers is that upper-airway epithelial cells are covered by a negatively charged and viscous mucus layer that often traps and neutralizes the cationic liposome-DNA complexes, leading to inefficient transfection rate in vivo after topical administration.Citation13,Citation14 Furthermore, Duncan’s results suggested that pulmonary surfactant inhibits cationic liposome-mediated gene delivery to respiratory epithelial cells.Citation15 In addition, cytotoxicity of cationic liposomes should be of concern. These limitations restricted the application potential of cationic liposomal vectors in airway epithelia gene therapy.
Some studies have taken the anionic liposomal DNA delivery vectors as an alternative to cationic liposomes.Citation16 Anionic lipids, in general, are less cytotoxicCitation17 and some of them are the endogenetic components of eukaryotic cell membranes.Citation18 It was demonstrated that a novel anionic lipoplex DNA delivery system encoding a gene of interest resulted in high transfection efficiency, good serum stability and low toxicity in vitro.Citation19
Although much progress in nonviral vectors has been made, as mentioned above, the low transfection efficiency in vivo is still a major obstacle in their clinical applications. Therefore, it is urgent to develop some novel strategies to overcome these limitations and enhance gene expression in the airway epithelium. One of those novel strategies, combinations of viral and nonviral vectors, has been proposed for airway gene therapy.Citation20,Citation21 In a previous report, we developed a novel delivery system by incorporating adenovirus vector into the anionic liposomes referred as AL-Ad5. Transmission electron microscopy using a negative stain technique demonstrated that in AL-Ad5 the adenoviruses were surrounded by lipid membranes. We also demonstrated that this combination strategy significantly enhanced the adenovirus-mediated gene transfer to both MDCK and CHO cells which are deficient of coxsackie–adenovirus receptors.Citation22
In our present study, to test whether such a combination strategy could improve the gene transducibility of adenovirus vector in CAR-deficient airway epithelia, we established and characterized a primary-culture model of differentiated murine tracheal epithelium. Moreover, according to the report that the polarized human airway epithelia expressed abundant folate receptors (FRs) on their apical surface,Citation23 the folatemodified phosphoethanolamine (F-PE) was incorporated into the formulation to form the folate-modified AL-Ad5 complexes (F-AL-Ad5). The transduction efficiencies of both AL-Ad5 and F-AL-Ad5 were assayed in vitro.
Materials and methods
Materials
The folate-modified egg phosphatidylethanolamine, folate-polyethyleneglycol-succinyl-eggphosphatidylethanolamine (F-PEG-SUC-EPE, F-PE), was synthesized as earlier reportedCitation24 by reaction between F-PEG-NH2 (conjugate prepared by folic acid and PEG-bis-amine) and SUCEPE (conjugate prepared by succinic anhydride and egg phosphatidylethanolamine). The fluorescein DHPE, N-fluorescein-5-thiocarbamoyl-1,2-dihexadecanoyl-sn-glycero-3-phosphoethanolamine triethylammonium salt (excitation/emission maxima ∼496/519 nm) and 5-bromo-4-chloro-3-indolyl-β-galactosidase (X-gal) were purchased from Invitrogen. The Ultroser-G serum substitute was obtained from Pall France/Division Pall BioPharmaceuticals (Pall 15950–017). The type VI acid-soluble human placental collagen (C7521) and mouse monoclonal anti-human pan cytokeratin (C2562) were from Sigma-Aldrich. FITC-labeled goat anti-mouse IgG (sc-2010) was purchased from Santa Cruz Biotechnology. The β-galactosidase enzyme assay system was from Applied Biosystems, and a bicinchoninic acid (BCA) assay kit was from Pierce, USA. The other agents such as CaC12, ethylenediaminetetraacetic acid (EDTA), sodium pyruvate, Pronase, DNase and insulin were obtained from Sigma Chemical Co (St Louis, MO).
Specific pathogen-free C57BL/6N male mice, 5–7 weeks old, were purchased from the Laboratory Animal Center of Sichuan University (Chengdu, China). The study was approved by the Animal Ethics committee of Sichuan University West China Medical School, and all procedures with animals were conducted according to the guidelines of the local Animal Use and Care Committees of Chengdu and executed according to the National Animal Welfare Law of China.
The adenovirus vectors used for this study are based on the Ad5 backbone, E1/E3-deleted. Ad5-LacZ expressing β-galactosidase was purchased from the Vector Gene Technology Company (VGTC, Beijing, China). The adenovirus titre was detected using plaque-forming assay on HEK 293 cells.Citation25
Preparation of F-AL-Ad5 and optimization of folate-PE mole ratio in formulation of F-AL-Ad5
The folate-modified complexes of adenoviral anionic liposomes (F-AL-Ad5) were prepared by the calcium-induced phase changes method according to a previous report.Citation22 Briefly, the lipid mixture of PC/CHEMS/Chol/folate-PE (5:4:1:0.01 mol/mol) was dissolved in chloroform and dried into a thin film on the wall of a 12-mm glass tube under a stream of nitrogen gas. The film was further dried under reduced pressure for 4 hours to ensure no trace of chloroform. Then, the dried, thin film was hydrated with Tes-buffered saline (100 mM NaCl, 2 mM histidine, 2 mM Tes, pH 7.4). Blank folate-modified anionic liposomes (F-AL) were formed by sonicating for 100 seconds intermittently (100 W) in an ice-bath sonicator. Ca2+ was introduced by direct addition of 100 mM CaCl2 to the preformed F-AL (final concentration 10 mM) and incubated at 25°C for 1 hour. The resulting precipitate was pelleted by centrifuging at 2500 × g for 10 minutes. The pellet was resuspended at a lipid concentration of 10 μmol/mL in a concentrated viral solution by vortexing for 10 minutes at room temperature. EDTA (100 mM) was added directly to this solution with a final concentration of 15 mM, and 10–20 μL of 100 mM NaOH was added to adjust the pH to 7.4. The solution was vortexed for 10 minutes followed by incubation for 30 minutes to form the folate-modified complexes of adenoviral anionic liposomes (8.5 × 108 viral particles/mg lipids).
Three types of fluorescently labeled formulations were prepared, in which the varying molar ratio of folate-PE was 1%, 5%, and 10%, respectively. Additionally, to fluorescently label the formulations, 1% fluorescein DHPE was included in the lipid component of each formulation. The polarized primary-cultured tracheal epithelial cells were used to test the cell uptake of the preformed fluorescently labeled formulations by flow cytometric analysis at excitation of 488 nm (BD FACS Canto II Flow Cytometer, USA).
Isolation and culture of tracheal epithelial cells
The primary culture of tracheal epithelia was carried out by an air–liquid interface method as the previous reports described.Citation26,Citation27 Firstly, the following four kinds of solution are prepared including the collection media, dissociation media, culture media, and differentiation media in the same way the reports described.Citation26,Citation27 Then, mice tracheae were obtained from C57BL/6N mice and digested with dissociation media at 37°C for 60 minutes. The digestion was stopped with fetal calf serum. The cells were pelleted (200 × g for 10 minutes) and suspended in culture medium. This suspension was incubated at 37°C for 2 hours in a 100-mm culture dish (Corning, USA) to remove the nonepithelial cells. Nonadherent cells were collected, washed, and seeded at 4 × 105 cells in 200 μL of culture medium inside the transwell inserts (Costar Transwell clear, 0.4 μm pore) which were precoated with type VI acid-soluble human placental collagen, with 600 μL culture medium outside. The cells were incubated at 37°C in 6% CO2 in a humidified incubator for 3 days. The medium inside of the insert was removed along with any nonadherent cells and debris, and the medium outside (bathing the basolateral surface) was replaced with 600 μL of differentiation media on day 4, then twice weekly. Any apical fluid should be sucked off every day, and the apical side should not be fed. Once the cells have reached confluence and formed tight junctions, the transduction studies were carried out.
Evaluation of the primary-cultured tracheal epithelium cells
Epithelia were tested for transepithelial resistance (Rte) by a Millicell–ERS (Electrical Resistance System, Millipore) and for morphology by scanning electron microscopy (SEM), taking the tracheal specimens from 5-week-old mice as control. SEM was performed according to the previous report.Citation26 Briefly, before viewed under a JSM–5900LV (JEOL, Japan) scanning electron microscope, the samples were washed with phosphate buffered saline (PBS), and then fixed firstly in PBS with 2.5% glutaraldehyde and secondly in 1% osmium tetroxide in PBS. Subsequently, the specimens were dehydrated through an alcohol series, treated with hexamethyldisilazane, and allowed to air-dry overnight.
To investigate the presence of cytokeratin, which was proved to be a typical character of airway epithelia, a fluorescence immunohistochemistry test was performed on primary-cultured cells using mouse monoclonal antihuman pan cytokeratin (Sigma-Aldrich, C2562) as a primary antibody and FITC-labeled goat anti-mouse IgG as a secondary antibody (Santa Cruz Biotechnology Inc, CA).
Infection of polarized primary cultures of murine tracheal epithelium cells by naked Ad5, AL-Ad5, and F-AL-Ad5
Fourteen days after seeding, the polarized primary cultures of murine tracheal epithelium cells were rinsed twice with PBS and then infected by adding 50 μL of the formulation of naked Ad5, AL-Ad5, and F-AL-Ad5 to the apical surface and the basolateral surface of the cultures, respectively. When infected from basolateral sides, the epithelia were turned upside down and 50 μL of formulation solution carefully applied to the bottom of the costar filter. A multiplicity of infection (MOI) of 40 was used in all transductions. After 4 hours, the suspension was removed, the cells were rinsed twice with PBS, and 200 μL of the culture medium was added to the inside insert and 600 μL to the outside insert. One day after infection, the transduction efficiency was assayed by X-gal staining and quantifying the LacZ gene expression, in which the β-galactosidase activities were determined using the β-galactosidase enzyme assay system (Applied Biosystems), and the total protein content of the lysates was measured by a BCA assay (Pierce, USA) using a standard of bovine serum albumin (BSA). Infectivity was quantified as pictogram of β-galactosidase (based on a standard curve for β-galactosidase activity) per microgram of total cell protein (pg/μg protein). The data were represented as mean ± standard deviation (SD) of three wells and was representative of three independent experiments.
The function of EDTA in the transduction of naked Ad5 was investigated. The naked Ad5 formulation was prepared in exactly the same way as the AL-Ad5 was prepared. As described above, during preparation of AL-Ad5 formulation, EDTA was introduced by calcium-induced phase changes method then diluted with DMEM medium at a final concentration of 1 mM. Therefore, the naked Ad5 prepared by the calcium-induced phase changes method also contained 1 mM EDTA when the transduction was performed. Meanwhile, the naked Ad5 solution containing 10 mM EDTA was also prepared. The transduction process was carried out in the same way as the operation mentioned above.
Statistical analysis
At least two independent sets of experiments were carried out, with similar results obtained. The results from one set were chosen to show in the present paper. Data obtained were represented as mean ± SD) and statistically analyzed using one-way ANOVA with Bonferroni post-tests (P < 0.05; P < 0.01).
Results
Particle size and zeta potential of folatemodified complexes of adenovirus vector and anionic liposomes (F-AL-Ad5)
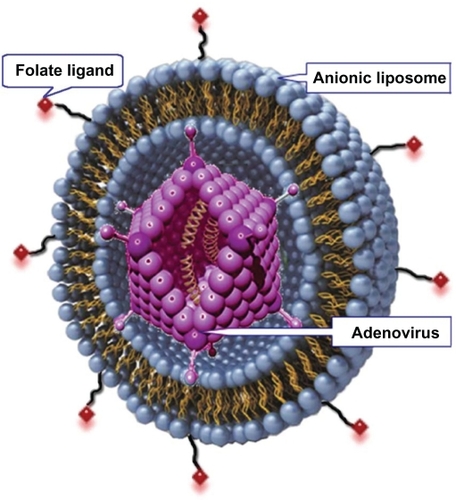
shows the schematic structure representation of F-AL-Ad5. The average particle size of F-AL-Ad5 (1% folate-PE) detected by Malvern Zetasizer Nano ZS90 (Malvern instruments Ltd, UK) was 225 ± 18 nm with a PDI (polydispersity index) of 0.253, which was similar to the size of AL-Ad5 (211 ± 10 nm) (P > 0.05). There was also little difference in zeta potential between F-AL-Ad5 and AL-Ad5 (−42.5 ± 1.8 mV for F-AL-Ad5 versus −42.2 ± 2.2 mV for AL-Ad5, P > 0.05).
Figure 1 Schematic representation of the structure of folate-modified adenoviral anionic liposome complexes (F-AL-Ad5).
Characterization of primary-cultured airway epithelia
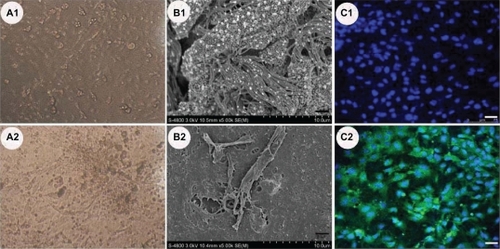
To test whether the primary culture of airway epithelia was performed successfully, investigations such as the morphology detection under light fields, SEM detection, Rte detection, and fluorescence immunohistochemistry assay for cytokeratin were carried out. Some of the results are summarized in . From the light fields’ images () under invert microscope, it could be seen that there were some island-like cells on the surface layer () from day 8. SEM was performed on primary-culture murine tracheal epithelial cells at a different time point, taking the tracheal specimens from 5-week-old animals of the same strain as control. A flattened epithelium was observed on day 4, with no features of differentiation. However, SEM analyses on days 8 and 21 revealed 13% ± 2% (n = 3 observations) and 21% ± 3% (n = 3 observations) ciliated cells,Citation26 respectively. By day 25, the percentage of ciliated cells was 26% ± 5% (n = 2 observations) (). The result of immunohistochemical analyses on day 8 () demonstrated the presence of cytokeratins, which was a typical characteristic of murine airway epithelial cells. The Rte detection result indicated that the transepithelial resistance of the primary-cultured cells was 2000–3000 Ω cm2 from day 4, then decreased and stabilized at about 1000 Ω cm2 by day 14, which indicated the cultured cells were well polarized.
Figure 2 Characterization on the primary-cultured airway epithelia. A1, A2) (×200) Cells under the light fields with some island-like cells on the surface layer A1). B1, B2) Scanning electron micrographs of a 5-week-old mouse trachea B1) and primary cultures of murine tracheal epithelium on day 25 B2). C2) Immunohistochemical detection of cytokeratins characterized in primary-culture murine tracheal epithelial cells with a negative control of C1). The bar in B1) and B2) is 2 μm and that of C1) and C2) is 30 μm.
Optimization of folate-PE by quantification of cell uptake using flow cytometry
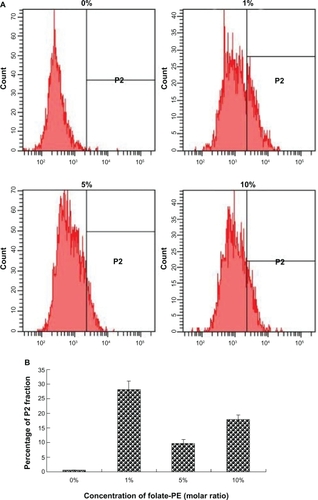
The optimal ratio of folate-PE used to prepare F-AL-Ad5 was determined by quantification of cell uptake using flow cytometry. As shown in , when the molar ratio of folate-PE in the total lipid was 1%, the uptake ratio of liposomes into tracheal epithelial cells was the highest. Therefore, 1% of folate-PE (molar ratio) was used to prepare the formulation of F-AL-Ad5 in the following experiment.
Figure 3 Flow cytometry data after incubation of fluorescently labeled F-AL-Ad5 with the polarized primary-cultured tracheal epithelial cells for 4 hours at 37°C. The profile of cell population count and fluorescence intensity A), and profile of the percentage parent in P2 area with concentration of folate-PE B) indicated the 1% formulation led to the highest uptake among all the test formulations.
Abbreviations: F-AL-Ad5, folate-modified complexes of adenovirus vector and anionic liposomes; folate-PE, folate-modified phosphoethanolamine; P2, positive cells.
Gene expression in the primary-cultured airway epithelia
To evaluate the transduction efficiency of obtained F-AL-Ad5 and AL-Ad5 complexes in primary-cultured cells, the formulation solutions were added dropwise to the apical and basolateral surfaces, respectively. When infected from basolateral sides, the transwell inserts were turned upside down and 50 μL of formulation solution carefully applied to the bottom of the costar filter. Additionally, in our preliminary experiments, we investigated the formulation’s transduction efficiencies with MOI of 20, 40, and 80 (pfu), respectively. The results indicated that at different MOI of adenovirus vectors, the tested groups exhibited similar alteration tendency of transduction efficacies. At MOI 40, the readout was sensitive enough to detect the LacZ gene expression. Therefore, in the present study, we carried out all the transduction experiments at MOI 40.
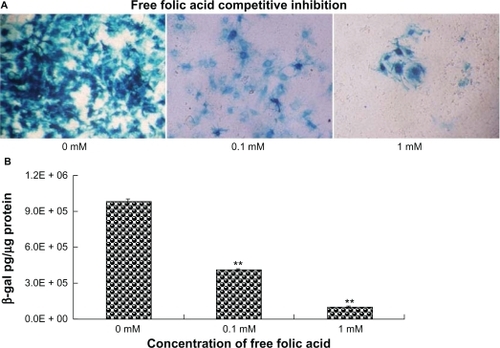
The result () showed that a higher LacZ expression was observed in the group of cells infected from the apical surface by AL-Ad5 and F-AL-Ad5 compared with cells infected with naked adenovirus vector (P < 0.01). Among all the tested formulations, we observed that F-AL-Ad5 significantly increased the transfection efficiency along the polarized airway epithelia (P < 0.01). Moreover, the transfection of F-AL-Ad5 could be competitively inhibited by 0.1 mM or 1 mM free folic acid (), indicating that the cellular delivery of F-AL-Ad5 is mediated by folate receptors.
Figure 4 Transduction levels of polarized primary-cultured tracheal epithelial cells. Primary cultures were transduced with naked Ad5 (MOI 40), AL-Ad5, and F-AL-Ad5 (total lipids 0.2 mg/mL), containing Ad5-LacZ at an MOI of 40 applied to the apical and basolateral surface, respectively, followed by X-gal staining A) (×200) and quantitative detection of β-galactosidase gene expression B).
Notes: Results are presented as mean ± standard deviation (n = 3), *P < 0.05, **P < 0.01, ANOVA.
Abbreviations: Ad5, adenovirus vector; AL, anionic liposome; F, folate; MOI, multiplicity of infection.
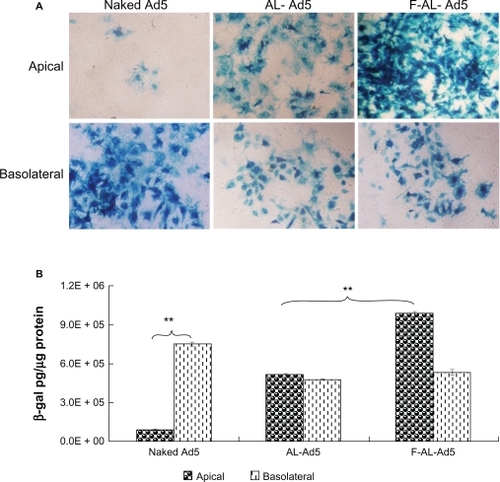
Figure 5 Inhibition effect on LacZ gene expression enforced by free folic acid. The primary-cultured tracheal epithelial cells were pre-incubated with 50 μL of free folic acid (0, 0.1, 1 mM) before adding the F-AL-Ad5 transfective agent to the apical surface followed by X-gal staining A) (×200) and quantitative detection of β-galactosidase gene expression B).
Notes: Results are presented as mean ± standard deviation (n = 3), *P < 0.05, **P < 0.01, ANOVA.
Abbreviations: Ad5, adenovirus vector; AL, anionic liposome; F, folate.
We also found that reduced transfection efficiency occurred when F-AL-Ad5 was administered basolaterally. This indicated the absence of FR at the basolateral side of murine airway epithelia. Additionally, LacZ expression of naked-adenoviral infected cells was significantly higher when the treatment was applied basolaterally compared with that applied apically. This suggested that naked adenovirus vector had the ability to infect polarized airway epithelia via the basolateral side, which may be due to the presence of CAR on the basolateral membrane.Citation8
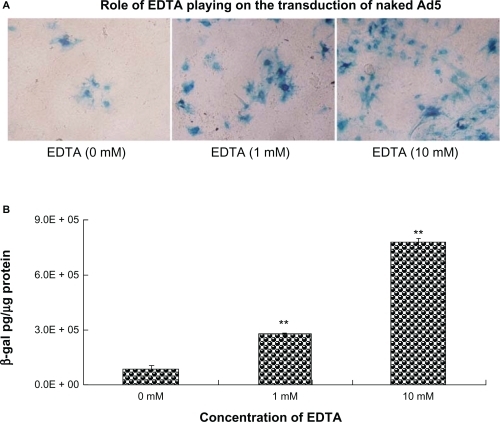
The result of transduction efficiency mediated by naked Ad5 with or without EDTA is demonstrated in . Both the profile of qualitative X-gal staining () and quantitative detection of LacZ gene expression () indicated that EDTA could enhance the transduction efficiency of naked Ad5 in the polarized epithelium cells. shows that in the group of naked Ad5 with 1 mM EDTA, the transduction efficiency was 2.2-fold higher than that in the group without EDTA. When the concentration of EDTA increased to 10 mM, the transduction efficiency increased ∼8.15-fold. This result suggested the contribution of EDTA for higher gene expression of F-AL-Ad5 and AL-Ad5.
Figure 6 Effect of EDTA on the transduction of naked Ad5. EDTA was mixed with naked Ad5 solution before the transduction at a final concentration of 1 and 10 mM, respectively. These mixtures containing Ad5-LacZ at an MOI of 40 were applied to the primary-cultured tracheal epithelia from the apical surface followed by X-gal staining A) (×200) and quantitative detection of β-galactosidase gene expression B).
Notes: Results are presented as mean ± standard deviation (n = 3), *P < 0.05, **P < 0.01, ANOVA.
Abbreviations: Ad5, adenovirus vector; EDTA, ethylenediaminetetraacetic acid; MOI, multiplicity of infection.
Discussion
Improving the efficiency of gene transfer remains an important goal in developing new treatments for cystic fibrosis and other diseases. Adenovirus vectors and nonviral vectors each have specific advantages, but they also have limitations. Adenovirus vectors can efficiently escape from the endosome and enter the nucleus, but the virus shows limited binding to airway epithelia. Nonviral cationic vectors can be efficiently internalized by airway epithelia, but they do not catalyze subsequent steps in gene transfer. In our previous study, the combination strategy of anionic liposomes and adenovirus vector was found to increase the efficiency of adenovirusmediated gene transfer to coxsackie–adenovirus receptor deficient cells.Citation22 In this novel delivery system, by incorporating adenovirus vector into liposomes, we were able to take advantage of adenovirus-dependent processes that facilitate gene expression such as escape from the endosome and entry into the nucleus.Citation28 These processes are significant barriers to nonviral vector-mediated gene transfer.
In the present study, we hypothesized that combination strategy could also improve the efficiency of gene transduction mediated by adenovirus vector in CAR-deficient airway epithelium. The transduction results suggested the LacZ gene expression mediated by naked Ad5 from the apical side was as poor as predicted due to lack of CAR on the apical surface of airway epithelia. But the complexes of AL-Ad5 increased the LacZ gene expression when infected from the apical side. This result was consistent with a previous paper reporting that anionic liposomes increased the efficiency of adenovirusmediated gene transfer to CAR deficient cells.Citation22
Additionally, an ideal gene carrier system should efficiently accumulate in specific target tissues with minimal toxicity to nontarget tissues.Citation29 In order to specifically increase transfection efficiency of delivering system to targeted cell populations, various targeting ligands including antibodies, growth factors, peptides, transferrin, and folate have been conjugated to polymers and lipids.Citation30,Citation31 Folate receptors are vastly over-expressed in a wide variety of human tumors, but rarely are found on normal cell surfaces.Citation32 To further facilitate the uptake of AL-Ad5 by airway epithelia, folate was incorporated into this delivery system based on the report that the polarized human airway epithelia expressed abundant FR on their apical surface.Citation23 The obtained results in this study indicated that folate modification to AL-Ad5 complexes could further enhance the gene transduction in airway epithelium in vitro and the enhancement could be inhibited by free folate in the medium, suggesting FR was playing an important role in the gene transduction of airway epithelia. Meanwhile, when these folate-modified AL-Ad5 complexes were applied to transduce the human ovarian cancer cells SKOV3, which are rich in folate receptorsCitation33 and deficient of coxsackie–adenovirus receptors,Citation34 a dramatic enhancement of gene expression was observed compared with that of unmodified AL-Ad5 complexes (data not shown). This result also suggested ligand modification was a useful approach to further improve the uptake of AL-Ad5 complexes by the targeted cell population.
In previous reports, airway epithelia cell lines such as CRL-9483 or 16HBE14o-cells were used to evaluate the efficiency of gene transfer,Citation35,Citation36 but the air–liquid interface primary-cultured airway epithelial cells are more appropriate models in vitro to assess transduction efficiency of carriers in airway epithelia, which accurately represents the behavior of polarized, differentiated airway epithelial cells. Therefore, in the current study, we set up a primaryculture model of differentiated mouse tracheal epithelium on semipermeable support membranes. The characteristics of the primary-culture model were investigated by SEM, immunohistochemistry, and electrophysiology, which confirmed the successful establishment of this model.
To further explore the possible reason that anionic liposomes could enhance the transduction level of Ad5 in airway epithelium, we investigated the function of EDTA on transduction. It was known that the CAR was located at the basolateral side of airway epithelia, and the tight junctions (TJs) play a central role in sealing the intercellular space between the apical and basolateral compartments of epithelial and endothelial cells. Therefore, when adenoviral vectors were applied to the mucosal surface of airway epithelia, the transduction efficiency was poor. Modulation of the barrier function of the TJ has been investigated as a strategy to enhance transmucosal drug absorption.Citation37–Citation39 Some agents such as a phospholipid detergent, surfactant, ethyleneglycol-bis (2-aminoethyl ether)-N,N,N’,N’-tetraacetic acid (EGTA) are attractive as potential therapies to enhance absorption of gene transfer vectors due to the rapid onset of action (within minutes) and their relatively rapid recovery (within hours) after treatment.Citation40 It has been reported that the integrity of the intercellular junctions requires a finite concentration of extracellular Ca2+. Removal of Ca2+ by chelators such as EGTA leads to significant increases in paracellular permeability and gene transfer efficiency.Citation41,Citation42 It was reported that gene transfer via the apical surface of human airway epithelia in vitro was improved by formulating vectors with EGTA in a hypotonic buffer. When Ca2+ chelators were applied to rabbit tracheal epithelia or human nasal epithelia in vivo, the transepithelial voltage decreased, and amiloride sensitivity was lost, suggesting that epithelial junctions opened.Citation43,Citation44 In our study, the complexes of anionic liposomes and adenovirus vector (AL-Ad5) were prepared under the help of EDTA by the calcium-induced phase changes method. A final concentration of 1 mM of EDTA remained in the obtained formulation of AL-Ad5 complexes. EDTA is also one kind of Ca2+ chelator, whose structure and function are similar to EGTA. When used in murine airway epithelia, AL-Ad5 exhibited an increased gene expression compared with naked adenovirus vector. To investigate whether the enhanced gene expression was due to the residual EDTA or the lipid formulation, different concentrations of EDTA was added to naked Ad5 before the transduction. As shown in , when naked Ad5 was mixed with 1 mM EDTA, whose concentration was the same as that in the formulation of AL-Ad5 or F-AL-Ad5, a 2.2-fold enhancement of gene expression was observed in primary-cultured differentiated cells. But the increased transduction efficacy was still lower than that obtained by the formulation of AL-Ad5 (2.72 ± 0.13) × 105 for naked Ad5 versus (5.15 ± 0.81) × 105 for AL-Ad5, P < 0.05). These results suggested that the higher transduction efficiency of AL-Ad5 was not only due to the residual EDTA, but was also contributed to by the formation of Ad5-liposome complexes. EDTA could open the tight junction of airway epithelia according to the other reports.Citation43,Citation44 Meanwhile, in our previous experiments, no cytotoxity was observed on A549 cells for the AL-Ad5 formulation.Citation22
Conclusion
In the present study, we prepared the complexes of anionic liposomes and adenovirus vector (AL-Ad5) by the calciuminduced phase changes method, and also incorporated folate-PE into lipid components to obtain folate-modified complexes (F-AL-Ad5). The results from the in vitro study indicated that in primary-cultured airway epithelial cells, AL-Ad5 and especially F-AL-Ad5 showed significant enhancement in gene transduction efficiency. This novel gene delivery system may be a useful strategy to deliver therapeutic genes to airway epithelia in vivo and other cells with overexpressed folate receptor.
Acknowledgements
We are thankful for the financial support of the National Natural and Science Foundation of China (No. 30973657) and the National Science and Technology Major Project of China (No. 2009ZX09310-002). We also thank Prof Weiyao Lu in Fudan University for synthesizing folate-PE.
Disclosure
The authors report no conflicts of interest in this work.
References
- GoldmanMJYangYWilsonJMGene therapy in a xenograft model of cystic fibrosis lung corrects chloride transport more effectively than the sodium defectNat Genet1995921261317719338
- RosenfeldMAYoshimuraKTrapnellBCIn vivo transfer of the human cystic fibrosis transmembrane conductance regulator gene to the airway epitheliumCell19926811431551370653
- ZabnerJPetersenDMPugaAPSafety and efficacy of repetitive adenovirus-mediated transfer of CFTR cDNA to airway epithelia of primates and cotton ratsNat Genet19946175837511023
- ZuckermanJBRobinsonCBMcCoyKSA Phase I study of adenovirus-mediated transfer of the human cystic fibrosis transmembrane conductance regulator gene to a lung segment of individuals with cystic fibrosisHum Gene Ther199910182973298510609658
- WestJRodmanDMGene therapy for pulmonary diseasesChest2001119261361711171744
- TomkoRPXuRPhilipsonLHCAR and MCAR: the human and mouse cellular receptors for subgroup C adenoviruses and group B coxsackievirusesProc Natl Acad Sci U S A1997947335233569096397
- ViglBZgraggenCRehmanNBanziger-ToblerNEDetmarMHalinCCoxsackie- and adenovirus receptor (CAR) is expressed in lymphatic vessels in human skin and affects lymphatic endothelial cell function in vitroExp Cell Res2009315233634719007771
- WaltersRWGrunstTBergelsonJMFinbergRWWelshMJZabnerJBasolateral localization of fiber receptors limits adenovirus infection from the apical surface of airway epitheliaJ Biol Chem199927415102191022610187807
- GriesenbachUAltonEWGene transfer to the lung: lessons learned from more than 2 decades of CF gene therapyAdv Drug Deliv Rev200961212813919138713
- IwamotoHSTrapnellBCMcConnellCJDaughertyCWhitsettJAPulmonary inflammation associated with repeated, prenatal exposure to an E1, E3-deleted adenoviral vector in sheepGene Ther1999619810610341881
- PolitoAJProudDEpithelia cells as regulators of airway inflammationJ Allergy Clin Immunol199810257147189867498
- KnowlesMRBoucherRCMucus clearance as a primary innate defense mechanism for mammalian airwaysJ Clin Invest2002109557157711877463
- GaoXKimKSLiuDNonviral gene delivery: what we know and what is nextAAPS J200791E92E10417408239
- RoseneckerJNaundorfSGerstingSWInteraction of bronchoalveolar lavage fluid with polyplexes and lipoplexes: analysing the role of proteins and glycoproteinsJ Gene Med200351496012516051
- DuncanJEWhitsettJAHorowitzADPulmonary surfactant inhibits cationic liposome-mediated gene delivery to respiratory epithelial cells in vitroHum Gene Ther1997844314389054518
- SrinivasanCBurgessDJOptimization and characterization of anionic lipoplexes for gene deliveryJ Control Release20091361627019331848
- PatilSDRhodesDGBurgessDJAnionic liposomal delivery system for DNA transfectionAAPS J200464e2915760094
- LiangHHarriesDWongGCPolymorphism of DNA-anionic liposome complexes reveals hierarchy of ion-mediated interactionsProc Natl Acad Sci U S A200510232111731117816061807
- PatilSDRhodesDGBurgessDJBiophysical characterization of anionic lipoplexesBiochim Biophys Acta20051711111115904657
- CroyleMAChengXSandhuAWilsonJMDevelopment of novel formulations that enhance adenoviral-mediated gene expression in the lung in vitro and in vivoMol Ther200141222811472102
- PriceALimberisMGruneichJAWilsonJMDiamondSLTargeting viral-mediated transduction to the lung airway epithelium with the anti-inflammatory cationic lipid dexamethasone-spermineMol Ther200512350250916099413
- ZhongZShiSHanJZhangZSunXAnionic liposomes increase the efficiency of adenovirus-mediated gene transfer to coxsackie-adenovirus receptor deficient cellsMol Pharm20107110511519968324
- SinnPLHickeyMAStaberPDLentivirus vectors pseudotyped with filoviral envelope glycoproteins transduce airway epithelia from the apical surface independently of folate receptor alphaJ Virol200377105902591012719583
- LuWYLiuMPanJLiHMaJPreparation of folate-liposome and its delivery into cultured HeLa CellsJ Shanghai Med Univ20002748
- MitterederNMarchKLTrapnellBCEvaluation of the concentration and bioactivity of adenovirus vectors for gene therapyJ Virol19967011749875098892868
- DavidsonDJKilanowskiFMRandellSHSheppardDNDorinJRA primary culture model of differentiated murine tracheal epitheliumAm J Physiol Lung Cell Mol Physiol2000279766778
- DavidsonDJGrayMAKilanowskiFMMurine epithelial cells: isolation and cultureJ Cyst Fibros20042596215463928
- FasbenderAZabnerJChillonMComplexes of adenovirus with polycationic polymers and cationic lipids increase the efficiency of gene transfer in vitro and in vivoJ Biol Chem199727210647964899045673
- LutenJvan SteenbergenMJLokMCDegradable PEG-folate coated poly(DMAEA-co-BA)phosphazene-based polyplexes exhibit receptorspecific gene expressionEur J Pharm Sci200833324125118207707
- HashimotoMMorimotoMSaimotoHShigemasaYSatoTLactosylated chitosan for DNA delivery into hepatocytes: the effect of lactosylation on the physicochemical properties and intracellular trafficking of pDNA/chitosan complexesBioconjug Chem200617230931616536460
- KimYKChoiJYYooMKReceptor-mediated gene delivery by folate-PEG-baculovirus in vitroJ Biotechnol2007131335336117727999
- SudimackJLeeRJTargeted drug delivery via the folate receptorAdv Drug Deliv Rev200041214716210699311
- ZhengYCaiZSongXPreparation and characterization of folate conjugated N-trimethyl chitosan nanoparticles as protein carrier targeting folate receptor: in vitro studiesJ Drug Target200917429430319255895
- KimJSLeeSHChoYSChoiJJKimYHLeeJHEnhancement of the adenoviral sensitivity of human ovarian cancer cells by transient expression of coxsackievirus and adenovirus receptor (CAR)Gynecol Oncol200285226026511972385
- IssaMMKoping-HoggardMTommeraasKTargeted gene delivery with trisaccharide-substituted chitosan oligomers in vitro and after lung administration in vivoJ Control Release2006115110311216901570
- TuHYChenXLiJSignal transduction in respiratory syncytial virus infection-induced thymic stromal lymphopoietin expression in human epithelial cellsNan Fang Yi Ke Da Xue Xue Bao200727101581158317959543
- MorimotoKNakamuraTMorisakaKEffect of medium-chain fatty acid salts on penetration of a hydrophilic compound and a macromolecular compound across rabbit corneasArch Int Pharmacodyn Ther198930218262484010
- SoderholmJDOmanHBlomquistLVeenJLindmarkTOlaisonGReversible increase in tight junction permeability to macromolecules in rat ileal mucosa in vitro by sodium caprate, a constituent of milk fatDig Dis Sci1998437154715529690393
- CoyneCBRibeiroCMBoucherRCJohnsonLGAcute mechanism of medium chain fatty acid-induced enhancement of airway epithelial permeabilityJ Pharmacol Exp Ther2003305244045012606647
- LindmarkTSchipperNLazorovaLde BoerAGArturssonPAbsorption enhancement in intestinal epithelial Caco-2 monolayers by sodium caprate: assessment of molecular weight dependence and demonstration of transport routesJ Drug Target1998532152239606011
- DuanDYueYYanZMcCrayPBJrEngelhardtJFPolarity influences the efficiency of recombinant adenoassociated virus infection in differentiated airway epitheliaHum Gene Ther1998918276127769874274
- CoyneCBKellyMMBoucherRCJohnsonLGEnhanced epithelial gene transfer by modulation of tight junctions with sodium caprateAm J Respir Cell Mol Biol200023560260911062138
- WangGZabnerJDeeringCIncreasing epithelial junction permeability enhances gene transfer to airway epithelia In vivoAm J Respir Cell Mol Biol200022212913810657931
- MylesCSorscherEMatalonSEnhancement of adenovirusmediated gene transfer in lungs and epithelial cells by EGTAChest2002121Suppl 335S11893677