Abstract
The identification of extracellular phospholipid vesicles as conveyors of cellular information has created excitement in the field of drug delivery. Biological therapeutics, including short interfering RNA and recombinant proteins, are prone to degradation, have limited ability to cross biological membranes, and may elicit immune responses. Therefore, delivery systems for such drugs are under intensive investigation. Exploiting extracellular vesicles as carriers for biological therapeutics is a promising strategy to overcome these issues and to achieve efficient delivery to the cytosol of target cells. Exosomes are a well studied class of extracellular vesicles known to carry proteins and nucleic acids, making them especially suitable for such strategies. However, the considerable complexity and the related high chance of off-target effects of these carriers are major barriers for translation to the clinic. Given that it is well possible that not all components of exosomes are required for their proper functioning, an alternative strategy would be to mimic these vesicles synthetically. By assembly of liposomes harboring only crucial components of natural exosomes, functional exosome mimetics may be created. The low complexity and use of well characterized components strongly increase the pharmaceutical acceptability of such systems. However, exosomal components that would be required for the assembly of functional exosome mimetics remain to be identified. This review provides insights into the composition and functional properties of exosomes, and focuses on components which could be used to enhance the drug delivery properties of exosome mimetics.
Introduction
Cells are well known to communicate via soluble mediators or cell-cell contact, but in recent decades, intercellular communication through extracellular vesicles has also increasingly gained attention. The first notion of such vesicles arose when Wolf described the formation of “platelet dust” upon storage of blood platelets.Citation1 These phospholipid-rich particles were shown to exert coagulant activity and were later determined to be actively shed membrane-derived vesicles.Citation2 Since then our knowledge about such vesicles has expanded dramatically, and vesicle secretion is now widely accepted to occur in most, if not all, cell types. Characterization studies identified three main populations of extracellular vesicles, which are commonly classified based on their intracellular origin. Cells that undergo apoptosis fractionate their cellular content into subcellular apoptotic bodies in order to prevent leakage of possibly toxic or immunogenic cellular contents into the extracellular matrix (, left panel).Citation3 Apoptotic bodies appear as a heterogeneous group of vesicles, with sizes ranging from 50 nm to 5 μm and a buoyant density of 1.16–1.28 g/mL.Citation4–Citation7 They contain a variety of cellular contents, including DNA, RNA, and histones, and display “eat-me” signaling molecules, causing them to be rapidly cleared by macrophages.Citation8,Citation9 Due to their specific cellular content and high density, they may be distinguished from two other major vesicle populations, which show considerably more overlap.
Figure 1 Schematic presentation of the biogenesis and composition of the three main classes of extracellular vesicles. Apoptotic bodies (left panel) are formed when cells enter apoptosis, and may contain nuclear material such as histones and DNA. They are heterogenous in size (50–5000 nm), irregularly shaped and harbor a variety of cellular proteins. Microvesicles (middle panel) are formed by budding and subsequent fission of the plasma membrane. Selective incorporation of membrane proteins and cytosolic proteins takes place during formation, resulting in vesicles which may be enriched in specific proteins and lipids compared to the parent cell. Microvesicles are thought to be smaller than apoptotic bodies (50–1000 nm) and more homogenously shaped. Selective enrichment of cellular content also occurs during the formation of exosomes (right panel), however exosomes originate from budding into the limiting membrane of large endosomal structures named multivesicular bodies (denoted with MVB). This process is facilitated by endosomal proteins. Subsequent fusion of MVBs with the plasma membrane results in release of the exosomes. Exosomes are small (<100 nm), relatively homogenous in size, and may contain (endosomal) proteins involved in their assembly, such as CD9, Alix and TSG101.
Abbreviation: NC, nucleus.
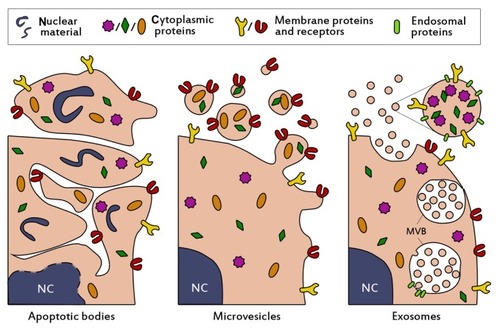
One of these populations originates from budding and fission from the plasma membrane into the extracellular space (, middle panel) and contains vesicles of about 50–1000 nm in size. Such vesicles are interchangeably referred to as microvesicles,Citation10 ectosomes,Citation11 shedding vesicles,Citation12 microparticles,Citation13,Citation14 plasma membrane-derived vesicles,Citation15 or even exovesicles.Citation16 In order to avoid confusion and promote standardization of nomenclature, the term “microvesicles” will be used in this review to denote this vesicle population. The buoyant density of microvesicles is still not well definedCitation7 and also the intracellular mechanisms for vesicle release remain unclear. Microvesicle secretion may take place in resting cells, but the vesicle shedding rate increases dramatically upon stimulation. The stimuli and intensity of stimuli required for vesicle formation can vary among cell types. As a common principle, increasing intracellular levels of Ca2+ result in increased secretion of microvesicles.Citation17,Citation18 For example, erythrocytes can be stimulated with high levels of extracellular Ca2+ in combination with a suitable ionophore.Citation19,Citation20 In various human tumor cell lines, microvesicle production was increased at extracellular Ca2+ concentrations up to 25 mM, but concentrations higher than 10 mM also decreased cell viability.Citation21 Other common stimulators for microvesicle release include lipopolysaccharide for monocytes,Citation22 and activation of the P2X7 receptor with ATP for macrophages and other myeloid cells.Citation23,Citation24
The last population of secreted membrane vesicles comprises exosomes, which differ from microvesicles mainly in their intracellular origin. Whereas microvesicles are supposedly generated by budding from the plasma membrane, exosomes appear to be formed by tightly controlled inward budding into large multivesicular bodies in the cytosol. These multivesicular bodies are able to fuse with the plasma membrane, causing the release of exosomes into the extracellular space (, right panel).Citation25 In theory, exosomes and microvesicles are clearly distinguishable by their origin, but in practice such a distinction is seldom possible. Therefore, attempts are made to characterize and separate both populations based on phenotypical features, such as buoyant density, size, morphology, and protein and lipid composition. Exosomes typically share characteristics with the vesicles inside multivesicular bodies. They are commonly 40–100 nm in size,Citation26 although somewhat larger diameters have been reported.Citation27 When analyzed by electron microscopy, exosomes show a typical cup-shaped morphology,Citation28 but in biological fluids they are likely to exist as spherical structures.Citation29 In addition to being more homogenous in size than microvesicles, they are thought to be more dense and float on a sucrose density gradient at heights of 1.13–1.19 g/mL.Citation26 Furthermore, exosomes often contain specific proteins which are incorporated during the formation process in multivesicular bodies, such as Alix, TSG101, and tetraspanins (CD9, CD63),Citation7,Citation30 which are frequently used as markers for their identification.
Exosomes and microvesicles are involved in a large variety of body processes. They are concentrated carriers of genetic and proteomic information, and thus are believed to play important roles in intercellular communication. Secreted vesicles can transfer their messages in different ways. Firstly, they may activate target cells via ligands expressed on their surface. For example, it has been demonstrated that antigen-presenting exosomes derived from dendritic cells provoke T cell-mediated immune responses in vivo.Citation31 In addition to eliciting immune responses, ligand-receptor signaling via exosomes can also play a role in other regulatory processes, such as angiogenesis,Citation32 hemostasis,Citation33 and cancer progression. Citation34 Secondly, secreted vesicles may transfer surface receptors from one cell to another by budding and subsequent fusion with plasma membranes of target cells.Citation35,Citation36 This mechanism may be exploited by the human immunodeficiency virus, increasing susceptibility to infection by transferring CD4 receptors from infected cells to noninfected cells.Citation37 Additionally, the shedding of microvesicles has been proposed to be a mechanism by which cells protect themselves from potentially harmful substances.Citation12 A third mechanism of action is based on the horizontal transfer of proteins and genetic material, such as micro RNAs (miRNA), between cells. The cytosolic content of a donor cell can be transferred directly to the recipient cell following fusion or internalization of microvesicles and exosomes.Citation38,Citation39 This results in the release of cargo into the cytosol of the target cell and subsequent intracellular signaling. An example of protein transfer between cells has recently been presented by Sarkar et al, who elegantly demonstrated that the intracellular apoptotic protein caspase-1 was transferred between monocytes and vascular smooth muscle cells via microvesicles, inducing apoptosis in the latter.Citation40 The encapsulation of caspase-1 in microvesicles was shown to be necessary for the apoptotic effect of the enzyme.
First evidence of exosome-mediated transfer of mRNAs and miRNAs was recently presented by Valadi et al, who showed that exosomes from mouse mast cells contained substantial amounts of RNA.Citation41 Many of the miRNAs were enriched compared with the parent cells, indicative of a selective incorporation process. Furthermore, mouse exosomes containing mRNA could be taken up by human mast cells, which resulted in the expression of mouse proteins in these cells. Additional evidence of functional miRNA transfer by exosomes was provided by Montecalvo et al, who showed that exosomes released from dendritic cells contained a variety of miRNAs.Citation42 These exosomes were taken up by acceptor dendritic cells and effectively repressed target mRNA expression. Thus, while it has been suggested that only a minority of all circulating miRNA is confined within exosomes,Citation43,Citation44 exosomal miRNA may potently regulate gene expression in target cells in vivo. In addition to mRNA and miRNA, mitochondrial DNA (mtDNA) has been found in exosomes of astrocytes, myoblasts, and glioblastoma cells.Citation45,Citation46 Given that horizontal transfer of RNA by exosomes or microvesicles is important for cellular communication, vesicle-associated RNA may also serve as a diagnostic tool in disease. Disease states may cause specific miRNAs to be enriched in extracellular vesicles compared with healthy controls. For example, extracellular vesicles from patients with prostate cancer were selectively enriched in miR-141 compared with healthy controls,Citation47 and EGFRvIII mRNA in circulating microvesicles was associated with clinically distinct subtypes of glioblastoma.Citation48 Viral infections may also be detected by analysis of exosomal RNA content.Citation49
Exosomes and microvesicles are naturally adapted for the transport and intracellular delivery of proteins and nucleic acids. This makes them particularly attractive for the delivery of pharmaceutical proteins and nucleic acids, such as short interfering RNA (siRNA). Intracellular delivery of siRNA is a challenging task, given that naked siRNAs are rapidly degraded in the circulation, their large size and negative charge limits membrane passage and cellular uptake, some siRNA sequence motifs may elicit undesired immune responses, and targeting to specific tissues and cells is required to reduce adverse effects caused by off-target silencing.Citation50,Citation51 Encapsulation of nucleic acid-based therapeutics in endogenous transporting vesicles is a promising novel strategy to overcome most of these delivery issues. Exosomes may be most suitable for such strategies, because they are small (40–100 nm), relatively homogenous in size, and well studied. Their size <100 nm is advantageous for their use as drug delivery systems, because this allows them to evade rapid clearance by the mononuclear phagocyte system and enhances passage through fenestrations in the vessel wall, as might occur during inflammation.Citation52
Exosome-based drug delivery systems: biotechnological versus synthetic approaches
The successful use of exosomes for the targeted delivery of siRNA has been recently demonstrated by Alvarez- Erviti et al.Citation53 They harvested dendritic cells from mice and transfected them to express the neuronal targeting ligand, RVG, coupled to the exosomal membrane protein, Lamp2b. This protein was expressed by the cells and incorporated in secreted exosomes. The exosomes were harvested, purified, and loaded with siRNA against an important protein in Alzheimer pathogenesis (BACE1) by electroporation. When the modified exosomes were injected intravenously in wild-type mice, a 60% decrease of BACE1 mRNA in the brain cortex was observed after 3 days. This ultimately resulted in a decrease (55%) of the harmful β-amyloid 1–42 protein in the brain. Moreover, no increase in serum interleukin-6, interferon gamma-induced protein 10, tumor necrosis factor alpha and interferon alpha concentrations was observed after injection of the exosomes, suggesting that the modified exosomes were immunologically inert. However, immunological responses to repeated administration of exosomes were not evaluated, albeit repeated administration of exosomes loaded with siRNA against GAPDH did not result in a loss of silencing efficiency.
The biotechnological approach to create exosome-based delivery systems used by Alvarez-Erviti et al was the first demonstration of an exosome-based drug delivery system which showed efficient in vivo delivery of siRNA.Citation53 Other strategies to exploit exosomes for therapeutic purposes have also been reported. In 2005, Delcayre et al described an “exosome display technology” in which various antigens were fused to the C1C2 domain of lactadherin.Citation54 This protein domain binds to the lipid phosphatidylserine exposed by exosomes,Citation55 resulting in the presentation of the fused antigen to the immune system. When Chinese hamster ovary cells were transfected with fusion constructs of C1C2 and interleukin-2 or granulocyte/monocyte colony-stimulating factor, the exosomes derived from these cells were significantly enriched with the recombinant cytokines compared with the parent cells. Moreover, the recombinant exosomes were able to induce proliferative responses in interleukin-2 and granulocyte/monocyte colony-stimulating factor-dependent cell lines, respectively.Citation54 The therapeutic potential of C1C2-coupled antigen display by exosomes was further explored in subsequent studies. These showed that tumors secreting exosome-bound ovalbumin grew slower than tumors secreting soluble ovalbumin, due to an enhanced immune stimulatory effect of the former.Citation56 Furthermore, the tumor-associated antigens, carcinoembryonic antigen and HER2, elicited potent antitumor immune responses when recombinantly coupled with exosomes.Citation57 The antitumor potential of this approach was also demonstrated in two prostate cancer models, in which tumor growth was severely attenuated by vaccination with exosomes displaying the tumor antigens, prostate-specific antigen or prostatic acid phosphatase.Citation58 The feasibility of antitumor therapy based on immunostimulatory exosomes was evaluated in two Phase I trials.Citation59,Citation60 In these trials, dendritic cells of patients with stage III/IV melanoma were isolated and pulsed with MAGE3 tumor antigens. Exosomes presenting MAGE3 were isolated and readministered to melanoma patients. Therapy appeared to be well tolerated by all patients and induced the desired immune effects in some patients, showing clinical feasibility for exosome-based therapeutics.
In addition to C1C2 coupling, therapeutics may also be nonspecifically bound to exosomes. This was recently demonstrated by Sun et al, who showed that mixing curcumin with exosomes enhanced its bioavailability, stability, and solubility, and improved its anti-inflammatory activity in an in vivo lipopolysaccharide-induced septic shock model compared with curcumin alone.Citation61 Furthermore, intranasally administered mouse lymphoma exosomes facilitated curcumin and stat3 inhibitor delivery to brain microglia, inducing anti-inflammatory and antitumor effects, respectively.Citation62
The examples mentioned above all use endogenous exosomes in their full complexity (or with only minimal modifications to their natural content) to deliver therapeutic cargo. This may offer a range of advantages over conventional drug delivery systems, such as viral or synthetic (nonviral) nanoparticles. Virus-based drug delivery systems have some of the advantages of natural viruses, including their excellent capacity to invade host cells and incorporate their viral load into the host genome.Citation63 However, their potential for insertional mutagenesis or oncogenesis, high production costs, and risk of immunogenicity limit their clinical use.Citation64 Nonviral drug delivery vehicles, such as cationic polyplexes and lipoplexes, are generally considered to be less immunogenic and mutagenic, but cytotoxicity and low transfection efficiencies in vivo are still major challenges to overcome.Citation64,Citation65
Exosome-based drug delivery systems may provide unique advantages over other systems, including limited or no undesired immunogenicity when self-derived exosomes are used, greater stability in the blood due to evasion of complement and coagulation factors,Citation66 efficient delivery of cargo into the cytosol of the target cell, and possibly fewer off-target effects due to the natural tendency of exosomes to act on specific target cells.
Despite these advantages, there are still some major obstacles and challenges to overcome before endogenous exosomes may be used in a clinical setting. Naturally derived exosomes are complex structures which are difficult to characterize pharmaceutically. In addition, they have complicated roles in health and disease, which are still poorly understood.Citation27,Citation67,Citation68 Therefore, they may induce adverse effects when used in pharmaceutical preparations. Thorough characterization of exosome content and function is required to avoid such issues. Furthermore, biological fluids contain a mixture of extracellular vesicle populations from various cell types. The purification of single populations of cell-specific vesicles (eg, cancer cell-derived exosomes) still provides a barrier which hinders translation to the clinic. Highly purified populations of exosomes may be obtained from exosome-secreting cell lines, but such exosomes may lack “self ” signals and elicit undesired immune responses. In addition, nanotechnological approaches for scalable production and efficient loading of exosomes are lacking, and remain an area of investigation.Citation69
A viable alternative for biotechnologically tailored exosome-based drug delivery systems is synthetic exosome mimetics. It is well possible that not all components in natural exosomes are required for specific and efficient delivery of cargo to the target cell. By extensive characterization of the lipid, protein, and nucleic acid content of exosomes, only functional components could be used for selective incorporation in exosome mimetics. Given that natural exosomes exist as spherical lipid bilayer structures, liposomes would provide a logical basis for the generation of exosome mimetics. Similar to exosomes, liposomes are bilayered phospholipid structures with (adjustable) diameters around 100 nm, which can be loaded with a variety of proteins, nucleic acids, or drug molecules.Citation70,Citation71 Liposomes have been shown to be valuable tools in drug delivery; several liposome-based drug delivery systems are currently in preclinical development and clinical trials, while others have been approved for clinical development.Citation71 The tailoring of liposomes to mimic exosomes could therefore provide a springboard for a novel class of nonviral drug delivery systems. Such drug delivery systems would have the benefits of endogenous exosomes (eg, limited immunogenicity, efficient cargo delivery, enhanced stability in body fluids), but could limit some of the problems anticipated with biotechnologically engineered exosomes. Production of exosome mimetics is more easily scalable for use in preclinical or clinical settings. In addition, the assembly process of exosome mimetics is controllable and results in the formation of “clean”, well characterized drug delivery systems with high pharmaceutical acceptability. Moreover, the use of exosome mimetics allows us to study the effect of each component separately. However, the components which are likely to be required for proper functioning of exosome mimetics as drug delivery systems are not yet well defined in the literature. Therefore, this review aims to describe potential candidate components for the assembly of functional exosome mimetics.
Lipids
In 2009, an initiative was undertaken by Mathivanan and Simpson to create an online database of all studies on exosomal protein and nucleic acid content, named ExoCarta.Citation72 This allows scientists to catalogue exosome-specific proteomic and genomic data and makes them available to other scientists in the field. ExoCarta has grown steadily since its development, and now contains 134 studies covering data of 4049 proteins, 1639 mRNAs, and 764 miRNAs. Because it is increasingly understood that not only proteins and nucleic acids contribute to exosomal function, the feature to include data on exosomal lipids in the database has also recently been added.Citation73 At this moment, only a handful of lipids has been registered in ExoCarta, including four prostaglandins (E2, F2, J2, and D2) and the conical lipid lysobisphosphatidic acid. Exosome-bound prostaglandins are involved in specific intracellular signaling pathways of the target cellsCitation74 and thus do not appear to be essential for exosomal stability and delivery.
Likewise, it is unlikely that lysobisphosphatidic acid plays a functional role in circulating exosomes. Exosomes derived from rat mast cells, human dendritic cells, and B cells have been found to contain only minor amounts of this lipid compared with their parent cells.Citation75,Citation76 Rather, the lipid may be involved in exosome biogenesis at the multivesicular body-limiting membrane, where its abundant presence has been shown.Citation77,Citation78 It has been postulated that lysobisphosphatidic acid could contribute to vesicle budding at this membrane, without being incorporated in the newly formed exosomes itself.Citation79 In addition, the lipid may be involved in fusion of endocytosed vesicles with lysobisphosphatidic acid-containing endosomal membranes. It has been demonstrated that some infectious particles, including vesicular stomatitis and dengue viruses, fuse with late endosomes through interactions with lysobisphosphatidic acid, resulting in the release of their contents into the cytosol of the target cell.Citation80–Citation82 Thus, the activity of lysobisphosphatidic acid appears to be limited to intracellular fusion and budding processes.
The lipid bilayer of circulating exosomes appears to be mainly constituted of plasma membrane lipids, including sphingomyelin, phosphatidylcholine, phosphatidyl-ethanolamine, phosphatidylserine, ganglioside GM3, and phosphatidylinositol.Citation79 The ratios of these lipids vary among exosomes from different cell types. For instance, almost half of the lipids in exosomes derived from reticulocytes constitute phosphatidylcholine,Citation83 whereas this lipid forms less than one third of total lipid content of mast cell-derived and dendritic cell-derived exosomes.Citation75 In general, exosomes appear to be enriched in sphingomyelin, cholesterol, GM3, and phosphatidylserine compared with their parent cells.Citation79 These lipids are not commonly used in liposomal drug delivery systems, but their incorporation can be advantageous. Sphingomyelin and cholesterol are thought to form hydrogen bonds, resulting in tight packing of sphingomyelin/cholesterol bilayers and low water permeability.Citation84,Citation85 In addition, they provide detergent resistance.Citation86 Therefore, incorporation of these lipids in exosome mimetics may increase their rigidity and stability. Indeed, liposomes containing sphingomyelin/cholesterol have longer circulation times and show decreased insertion of plasma proteins into their membrane.Citation87 Liposomes containing sphingomyelin/cholesterol and loaded with vincristine have shown promising results in Phase II clinical trials, being tolerated at twice the dose of nonencapsulated vincristine in patients with aggressive non-Hodgkin lymphoma and showing a favorable side effect profile.Citation88,Citation89
GM3 may act as a stabilizer of the exosomal wall and may shield the vesicle from interactions with blood components. Gangliosides, sphingomyelin, and cholesterol were demonstrated to act synergistically to decrease uptake of liposomes by the reticuloendothelial system both in vitro and in vivo.Citation90,Citation91 However, Yokoyama et al demonstrated with DPPG/GM3 liposomes that high molar concentrations (>15 mol%) of GM3 may induce membrane segregation and leakage from vesicles.Citation92 Therefore, GM3 may be beneficial in exosome mimetics when used in low concentrations only.
The structural function of phosphatidylserine in biological membranes is less well studied, but it is known to play a role as a signaling molecule in a large variety of biological processes. The biological role of phosphatidylserine may differ among vesicle populations and cellular sources. For instance, circulating erythrocytes gradually externalize phosphatidylserine during aging, providing an “eat-me” signal for the reticuloendothelial system.Citation93 Allen et al showed that low concentrations of phosphatidylserine (>2 mol%) in the membrane of erythrocyte-resembling liposomes were sufficient to induce clearance by this system dramatically, without affecting membrane lipid organization.Citation94 The “eat-me” signaling function of phosphatidylserine is nature’s way of clearing phosphatidylserine-exposing apoptotic cells by phagocytosis, reducing leakage of the cellular content to the extracellular matrix.Citation95 The lipid may also serve as a docking station for factors of the coagulation cascadeCitation96 and has been suggested to play roles in other physiological processes, such as muscle formation and anti-inflammatory responses.Citation97,Citation98
Whilst these characteristics appear to be unfavorable for exosome mimetics, for which longer circulation times are often desired, incorporation of phosphatidylserine in exosome mimetics may be beneficial nonetheless. The natural conical shape of phosphatidylserine may aid in the assembly of the curved exosome mimetic membrane.Citation99 In addition, conically shaped lipids facilitate fusion and fission of biological membranes.Citation100–Citation102 Phosphatidylserine could thus enhance fusion of exosome mimetics with target cell membranes and promote intracellular release of cargo. Indeed, it has been shown in a number of studies that inhibition of phosphatidylserine on microvesicle membranes by Annexin-V or Diannexin reduced fusion of microvesicles with the plasma membranes of their target cells,Citation36,Citation38,Citation103,Citation104 indicating that phosphatidylserine is an important mediator of vesicle fusion. Moreover, it has been postulated that the negatively charged phosphatidylserine enhances the stability of cell membranes by electrostatic interactions with (cationic) skeletal proteins.Citation105 This effect may be exploited in exosome mimetics loaded with cationic compounds, increasing the retention of encapsulated cargo. However, given that phosphatidylserine displayed on the outer layer of exosome mimetics may dramatically decrease their circulation time, incorporation of the lipid in exosome mimetics has to be carried out with caution.
Regardless of the exact lipid composition of extracellular vesicles, the lipid bilayer of exosomes appears to be adapted to their target environment. Extracellular pH may differ among target tissues, and rigidity of the vesicle needs to be maintained in these environments to assure optimal function. Parolini et al showed that exosomes released in an acidic microenvironment contained higher concentrations of sphingomyelin and GM3 compared with exosomes from the same cellular origin but released in a buffered environment.Citation106 These lipids were suggested to increase exosomal rigidity upon interaction with cholesterol.Citation107 Moreover, they demonstrated enhanced fusion of acidic exosomes with target cells compared with exosomes secreted in buffered conditions. Given that the target cells (metastatic melanoma) are surrounded by an acidic extracellular matrix under physiological conditions,Citation106 this suggests that the lipid composition of exosomes may be tailored to their target microenvironment. The influence of extravesicular pH on exosome behavior was also demonstrated by Laulagnier et al, who showed that exosomal rigidity increased from pH 5 to pH 7, a common transition during release of exosomes from acidic multivesicular bodies.Citation75 The rigid bilayer at physiological pH may enhance membrane fusion with target cells, but this was not investigated in this study.
Proteins
Although lipids are increasingly accepted to play important roles in exosomal function, exosome-associated proteins are indispensable. Such proteins can actively participate in regulatory processes and trigger cellular responses, but are also irrefutably involved in functional aspects of exosomes, such as assembly, (preventing) interaction with the extracellular matrix, and binding and fusion with target cells. For instance, it was demonstrated that cross-linking of proteins on exosomes by paraformaldehyde decreased exosomal fusion with parental cells by approximately 20%.Citation106 Furthermore, when these exosomes were solubilized with octylglucoside and reconstructed by dialysis (removing membrane proteins), they showed a dramatically reduced ability to fuse with target cells compared with untreated exosomes. The fusion efficiency of the protein-depleted exosomes was comparable with the fusion efficiency of large unilamellar vesicles with a lipid composition similar to that of the natural exosomes, confirming the importance of exosomal proteins in fusion events.Citation106
According to a variety of proteomic studies (catalogued in ExoCarta), a number of proteins and protein families are abundantly detected in exosomes.Citation30 Many of these, such as heat shock proteins, annexins, and proteins of the Rab family, are mainly involved in intracellular assembly and trafficking of exosomes and may not be required further after the vesicles are secreted. Thus, the inclusion of such proteins in exosome mimetics would probably not be beneficial for drug delivery purposes. However, several other exosomal proteins or protein families may be exploited to enhance the delivery properties of exosome mimetics. It should be noted that incorporation of proteins in exosome mimetics is still a challenging task, especially when simultaneous incorporation of multiple components is desired. However, work in this area is progressing and has yielded promising results.Citation108–Citation112 For instance, the functional reconstitution of a voltage-gated potassium channel and αbIIβ3 integrin in giant unilamellar vesicles was recently described.Citation108,Citation110 The potential advantages of the incorporation of several exosomal proteins in exosome mimetics are discussed below.
Tetraspanins
Tetraspanins are a family of transmembrane proteins commonly detected in exosomes. Among them are CD9, CD63, CD81, and CD82, which are often used as exosome markers.Citation30,Citation113 Some tetraspanins are selectively enriched in exosomes compared with their parent cells. An example is CD9, which was found to be more than 10-fold enriched in dendritic cell-derived exosomes.Citation114 CD37, CD63, CD81, and CD82 were abundantly detected in exosomes derived from B lymphocytes.Citation115 Atay et al demonstrated that CD81 was selectively enriched in exosomes from trophoblast cells, but could not detect CD63 (considered to be a canonical exosomal protein) in either parent cells or exosomes.Citation116 Tetraspanin enrichment patterns in exosomes thus vary among parent cells.
Tetraspanins have been relatively understudied due to their limited ligand-receptor interaction and their small size, which in some cases may prevent biochemical or immunological detection.Citation117 However, their functions as mediators of fusion, cell migration, cell-cell adhesion, and signaling events designate them as interesting targets in the field of drug discovery.Citation118 Moreover, their functions in exosomes (albeit largely unknown) may be exploited to enhance the properties of exosome mimetics as drug delivery systems.
The tetraspanin, CD9, has been shown to mediate fusion processes in a variety of cell types. Miyado et al demonstrated that egg cells from CD9 knockout mice (CD9−/−) failed to fuse with sperm cells from wild-type males, although binding between the two cell types was unaltered. Normal fusion events were observed between sperm cells and CD+/+ eggs, suggestive of an involvement of CD9 in the fusion process. Moreover, when CD9 on eggs was blocked with anti-CD9 antibodies, fusion was significantly inhibited.Citation119 A role of CD9 in fusion processes was also evidenced by Tachibana and Hemler, who showed that blocking CD9 and CD81 on myoblast cells inhibited cell fusion to syncytia, and upregulation of the two tetraspanins resulted in enhanced cell fusion.Citation120 It has been suggested that tetraspanin-mediated cell fusion is exploited by various viruses, allowing them to spread while avoiding exposure to the humoral immune system.Citation121,Citation122 However, the fusogenic properties of CD9 and CD81 were contradicted by studies of the fusion behavior of mononuclear phagocytes. Incubation of these cells with antibodies against CD9 and CD81 resulted in enhanced formation of multinucleated giant cells, and CD9−/− and CD81−/− alveolar macrophages formed 3–4-fold more multinucleated giant cells than wild-type cells.Citation123 These results were supported by Parthasarathy et al, who additionally showed that, in contrast with CD9 and CD81, CD63 in fact promoted monocyte fusion.Citation124 Thus, the fusogenic properties of tetraspanins appear to be dependent on cell type.
Interestingly, the tetraspanins which are often found to be enriched in exosomes (CD9, CD63, CD81, and CD82)Citation30,Citation113 have been shown to be involved in the migration of dendritic cells. In a chemotaxis assay, antibodies against the single tetraspanins increased migration of these cells by 50%–70%, while 100% increased migration was observed when these antibodies were combined.Citation125 The increase in migration was attributed to decreased binding of the cells to integrins expressed in the extracellular matrix. These results suggest that tetraspanins in exosomal membranes may facilitate their binding to target cells.
Although fusogenic and matrix-binding properties of tetraspanins in cell membranes have been described, not much is known about the functions of tetraspanins in exosomes. However, tetraspanins, Tspan8, CD49d, and CD106, appeared to be involved in binding and uptake of pancreatic adenocarcinoma-derived exosomes by fibroblasts and endothelial cells.Citation126 It has also been suggested that CD9 is involved in fusion of exosomes with their target cells,Citation52 indicating that tetraspanins may have similar functions in exosomes and cell membranes.
It is important to consider that tetraspanins probably exert most of their functions in unison with other membrane proteins. In cell membranes, tetraspanins are laterally organized in tetraspanin-enriched microdomains, which are resistant to mild detergent conditions. Tetraspanin-enriched microdomains generally contain integrins, signaling molecules, and other tetraspanins, and are involved in a variety of cell fusion, cell adhesion strengthening, and signaling events, as excellently reviewed previously.Citation117,Citation118,Citation127,Citation128 The effects of single tetraspanins, particularly in extracellular vesicles, are still poorly understood. Incorporation of tetraspanins in exosome mimetics may therefore not only enhance exosome mimetic functioning (ie, binding and uptake in target cells), but may also provide important clues on tetraspanin functioning in exosomes.
Adhesion molecules
According to the ExoCarta database, integrins are among the most abundant proteins detected in exosomes derived from cancer and immune cells.Citation129 Integrins are membrane-spanning proteins which exist as heterodimers of α and β subunits. A total of 24 different heterodimers have been identified in vertebrates, of which β1, β2, and αv integrins comprise the largest group. They function primarily as adhesion molecules and establish cell binding to the extracellular matrix, although other functions have been described.Citation130,Citation131 In exosomes, integrins are most likely involved in addressing the vesicles to their target cells.Citation31 Therefore, incorporation of integrins in exosome mimetics may potentiate their interactions with the extracellular matrix, increasing their delivery potential.
Given that integrins are abundantly detected in a range of exosomal preparations (especially those derived from cancer cells),Citation132,Citation133 it is surprising that only a limited number of studies have addressed the functional role of these adhesion molecules in exosomes. In seminal work by Rieu et al, it was shown that integrin α4β1 is present on the surface of young reticulocytes and its surface levels decrease during maturation.Citation134 The adhesion molecules of mature reticulocytes were detected in the exosomal fraction, suggesting that the cells use exosomes to dispose of integrins. Functional examination of the exosomes showed that α4β1 integrin mediated binding of the exosomes to fibronectin. This binding was dependent on divalent cations (Mg2+, Mn2+) and was significantly inhibited by the α4β1-binding domain of fibronectin and anti-α4 antibodies. In similar work, it was found that exosomes from fibroblasts and epithelial cells contained β1 integrins and that exosomes from human B cells expressed both β1 and β2 integrins.Citation135 B cell-derived exosomes bound collagen I and fibronectin in a cation-dependent manner, and binding was inhibited by antibodies against α4 and β1 integrins. Moreover, it was shown that these exosomes strongly adhered to tumor necrosis factor alpha-stimulated fibroblasts through an integrin-intercellular adhesion molecule (ICAM-1) interaction. Integrin β1 and CD9 were also detected on the surface of retroviral particles, and it was suggested that these contribute to adhesion of viruses to host cells.Citation136 Taken together, these results suggest that exosomal integrins play a crucial role in establishing exosomal adhesion to the extracellular matrix. Given that binding to target cells is the first step in intracellular drug delivery, incorporation of integrins into exosome mimetics may thus greatly enhance their drug delivery potential. This strategy has not been implemented in drug delivery systems yet, but integrin-ligand interactions have been successfully exploited. For instance, the fibronectin-derived RGD peptide is commonly used to target drug delivery systems to αvβ3 integrin, which is overexpressed in tumor vasculature.Citation137 Integrin incorporation in exosome mimetics may work in a similar fashion to deliver therapeutics to specific tissues. It has been shown that integrin αIIbβ3 can be incorporated into liposomal membranes without losing its ability to bind to its ligands.Citation109,Citation138 However, the affinity of this integrin for its ligand, cyclo(RGDfV), was higher in cell membranes than in liposomes,Citation139 suggesting that other endogenous membrane molecules (eg, tetraspanins, lipids) contribute to integrin- mediated adhesion. Indeed, such a synergistic effect was demonstrated in a study by Zheng et al,Citation111 who incorporated integrin α5β1 into phosphatidylcholine/cholesterol liposomes containing increasing concentrations of ganglioside GM3 (0–10 nmol/55 μg phosphatidylcholine). The binding of the liposomes to fibronectin-coated plates differed significantly among varying concentrations of GM3, and was optimal at GM3 concentrations of 0.2–0.4 nmol (0.22–0.44 μg)/55 μg phosphatidylcholine. In addition, mammary carcinoma cells mutated to contain high concentrations of GM3 bound better to these plates than their parent cells, further evidencing the synergistic effect of other membrane components to integrin function.Citation111 Given that the membranes of exosome mimetics resemble those of endogenous exosomes, integrin incorporation may be a powerful strategy to enhance the adhesive properties of exosome mimetics.
In addition to integrins, other adhesion molecules which may be potentially exploited in exosome mimetics have been identified in exosomes. Comprehensive proteomic studies have identified thrombospondin 1 in exosomes in the urine, plasma, and saliva of healthy volunteers.Citation140–Citation143 In addition, thrombospondin 1 and thrombospondin 2 were detected in exosomes from patients diagnosed with various types of cancer, and in exosomes derived from a colon cancer cell line.Citation144,Citation145 Thrombospondin 1 contains a variety of adhesive domains, including an RGD sequence that binds αvβ3 integrins. Citation146 Nucera et al demonstrated that the protein plays an important role in cancer metastasis.Citation147 They showed that knockdown of the thrombospondin 1 gene in papillary thyroid carcinoma cells decreased their adhesion and migration in vitro and in vivo, and that treatment with a thrombospondin 1 inhibitor decreased metastasis and tumor growth in an orthotopic mouse model. In an attempt to use the adhesive properties of thrombospondins for targeted drug delivery, Rivera-Fillat et al developed aspartimide analogs of thrombospondin 1 and 2 and conjugated them to liposomes loaded with doxorubicin.Citation148 Both peptides adhered to endothelial cells and colon carcinoma cells in vitro. In addition, it was demonstrated that the thrombospondin 1 analog enhanced the antitumor effect of doxorubicin-loaded liposomes in mice bearing a human colon carcinoma; tumor growth was inhibited and mouse survival was prolonged compared with the nontargeted doxorubicin liposomes. This effect was most likely due to enhanced delivery of doxorubicin to target cells, given that targeted liposomes without doxorubicin did not exert antitumoral activity. In addition to conferring adhesion properties, thrombospondins also serve as signaling molecules with antiangiogenic and antitumoral effects. Therefore, they may have therapeutic potential on their own, and have been reviewed elsewhere.Citation149,Citation150 Thrombospondins may thus be valuable tools in tumor-targeted exosome mimetics; they may promote adhesion to target cells and induce antiangiogenic signaling pathways.
A third interesting class of adhesion molecules found associated with exosomes is the family of ICAMs. ICAM-1 and ICAM-3 have been detected in immune cell-derived exosomesCitation41,Citation115,Citation151–Citation153 and may function as important mediators in immune responses. ICAM-1 is a ligand for integrin αLβ2 (LFA-1) and Mac-1, and promotes leukocyte adhesion,Citation154 while ICAM-3 binds DC-SIGN on dendritic cells.Citation155 Their functions in exosomes are still under investigation, although some interesting results have been published. Segura et al showed that exosomes from mature dendritic cells contained markedly more ICAM-1 compared with those from immature dendritic cells.Citation156 Mature exosomes induced T cell activation and proliferation in vitro, and enhanced T cell activation and skin graft rejection in vivo compared with immature exosomes. The authors showed that ICAM-1 expression was an important contributor to this process, and suggested that ICAM-1 mediates exosome adhesion to target antigen-presenting cells, thereby inducing the immune response.Citation156 In a follow-up study, the group showed that ICAM-1-exposing exosomes induced significantly stronger T cell responses than ICAM-1−/− exosomes in vitro and in vivo, and that this effect was due to loss of exosome-cell adhesion mediated by ICAM-1 and LFA-1.Citation157 These results demonstrate the importance of ICAM-1 in the adhesion of exosomes to LFA- 1-expressing immune cells. When targeting the immune system (eg, in vaccine preparations), the incorporation of ICAM-1 into exosome mimetics may be considered.
Other membrane proteins
Apart from tetraspanins and adhesion molecules, other proteins which also have the potential to enhance the drug delivery properties of exosome mimetics have been identified in exosomes. Two of these, CD55 and CD59, have been found in exosomes derived from various sources, including B cells, dendritic cells, colorectal cancer cells, saliva, and bronchial epithelial cells.Citation66,Citation141,Citation145,Citation153,Citation158,Citation159 Both factors protect cells from complement-mediated lysis; CD55 accelerates the decay of autologous complement factors,Citation160 whereas CD59 prevents activation of the membrane attack complex by inhibition of C9 incorporation in C5b-9.Citation161 In an interesting study by Clayton et al, it was demonstrated that inhibition of exosomal CD55 resulted in increased membrane disposition of C3b and exosome lysis.Citation66 Blockade of CD59 also enhanced complement-mediated exosome lysis, and inhibition of both CD55 and CD59 increased lysis even further. These proteins thus appear to protect exosomes from complement-mediated lysis. Given that liposomes are prone to lysis mediated by complement factors,Citation162,Citation163 CD55 and CD59 may be used to improve the stability and circulation time of exosome mimetics.
Another exosomal protein with possible therapeutic applications is lactadherin (also known as EGF-factor VIII or MFG-E8).Citation114 This protein is commonly found in exosomes derived from immune cellsCitation114,Citation164,Citation165 and fibroblasts.Citation166 Lactadherin consists of two EGF-like domains and two lectin type C domains (C1 and C2) with high homology to coagulation factors V and VIII.Citation167 The EGF-like domains mediate binding to integrins αvβ3 and αvβ5 via an RGD motif, while the C2 domain promotes binding to phosphatidylserine in biological membranes.Citation168 Thus, it acts as a scaffold protein between the surface of phosphatidylserine-exposing apoptotic cells or exosomes and target cells. It has been reported that lactadherin promotes adhesion and uptake of exosomes by dendritic cells,Citation169 but also interactions with other phagocytotic cells expressing integrins αvβ3/5, such as macrophages and endothelial cells, are likely.Citation114 To demonstrate that angiogenic endothelial cells are capable of phagocytosis, Fens et al prepared phosphatidylserine-exposing liposomes and opsonized them with lactadherin.Citation170 Marked uptake of the opsonized liposomes in human umbilical vein endothelial cells was observed, whereas uptake was several-fold lower in the absence of lactadherin. Moreover, membrane vesicles containing egg phosphatidyl glycerol, which is a negatively charged phospholipid that lacks a lactadherin binding site, were barely taken up by human umbilical vein endothelial cells in the presence of the opsonin. These results show that lactadherin may be used to target exosome mimetics to specific integrin-expressing cells, such as angiogenic endothelial cells and dendritic cells.
Furthermore, it has been shown that fusion proteins containing the lactadherin C1C2 domain can be exploited to induce immune responses to tumor antigens.Citation56–Citation58 In such strategies, the phosphatidylserine-binding domain of the opsonin is used to coat exosomes and exploit the excellent delivery potential of these vesicles to deliver the antigen to antigen-presenting cells, resulting in immune responses. These studies illustrate the versatility of the lactadherin protein and its application in exosome mimetic-mediated drug delivery. An advantage of lactadherin is that it does not need to be incorporated in the membrane of exosome mimetics. Rather, it is possible to produce the protein recombinantlyCitation171 and attach it to phosphatidylserine-exposing exosome mimetics after vesicle formation. Additionally, fusion proteins of lactadherin and functional domains of other proteins can be engineered, conferring unique properties to exosome mimetics coated with these constructs. For instance, by fusing the C1C2 domain of lactadherin to targeting ligands, exosome mimetics may be targeted to specific organs and cell types. An additional advantage of lactadherin coating of phosphatidylserine-exposing exosome mimetics is that recognition of the lipid by phosphatidylserine receptors may be blocked. This way, the “eat-me” signaling function of phosphatidylserine would be inhibited, resulting in prolonged circulation time of the vesicles.
Decoration of exosome mimetics with specific proteins may also be achieved by the use of exosomal membrane anchors. In contrast with lactadherin, which is coated on the surface of exosome mimetics, such anchors may be plugged into the exosome mimetic membrane during exosome mimetic assembly and provide a stable scaffold for fused proteins. An example is Lamp2b, which was used by Alvarez-Erviti et al to anchor the neuron-specific RVG peptide to the exosomal membrane.Citation53 Although this study exploited the endogenous exosomal pathway to incorporate the anchor into exosomes, it may be possible to reconstitute the protein into an artificial membrane (ie, exosome mimetic membrane). Thus, linkage of exosome mimetics to proteins which are not naturally expressed in exosomes may be achieved, circumventing possible expression issues in biotechnologically engineered exosomes. Such issues were clearly demonstrated by Shen et al, who attempted to target a highly oligomeric cytoplasmic protein (TyA-GFP) to secreted vesicles by fusing the protein to a variety of exosomal membrane proteins, including CD4, CD38, CD43, CD69, and CD83.Citation172 After transfection of Jurkat T cells with expression vectors encoding the fusion proteins, only CD43-TyA-GFP could be weakly detected in the secreted vesicles. The authors suggested that endoplasmic reticulum-associated degradation could be responsible for the undetectable exosomal expression of some of the other constructs. A small number of constructs was degraded nor secreted, but was found to be retained in the endoplasmic reticulum instead. In this regard, the synthetic construction of exosome mimetics can offer a solution. A promising novel technique to incorporate membrane proteins into liposomes is the cell-free expression of such proteins in the presence of liposomes.Citation173–Citation175 Using this technique, integral membrane proteins are inserted into the liposomal membrane during their expression, resulting in functional and stable protein incorporation. Successful cell-free expression in liposomes has been demonstrated for a number of membrane proteins, such as membrane channel proteins.Citation175–Citation177
Therapeutic cargo
Exosome mimetics provide an opportunity to deliver therapeutic cargo directly into the cytoplasm of target cells. It is possible to load exosome mimetics with synthetic drugs, as is commonly done in other liposomal formulations,Citation178,Citation179 but the unique exosome-resembling characteristics of exosome mimetics allow them to excel in the delivery of therapeutic proteins and nucleic acids. An interesting class of molecules which could benefit from exosome mimetic-mediated delivery are miRNAs. These short noncoding endogenous RNA molecules recruit cellular proteins and bind to target mRNA sequences via Watson-Crick base-pairing, resulting in cleavage or translation repression of the target strand (RNA interference). This way, they regulate protein expression and cellular functioning.Citation180 Given the discovery that miRNAs are not only involved in the regulation of cellular processes in the miRNA-producing cell, but that they also regulate processes in other cells by exosomal transfer,Citation41 the therapeutic applications of miRNAs have gained increasing attention.
Intracellular levels of miRNAs have been extensively mapped in a number of diseases. Among the most studied diseases in this regard is cancer, in which specific miRNAs are often overexpressed or underexpressed compared with healthy cells.Citation181 Low levels of cancer-associated miRNAs can be compensated by miRNA-based therapeutics (so-called miRNA restoration or replacement), resulting in improvement of the disease outcomes. Successful restoration of miRNA to treat various types of cancer has been described in a number of recent studies and was reviewed recently by Henry et al.Citation182 For instance, in 2009, Kota et al showed that the expression of the miRNA, miR-26a, in liver tumors was markedly decreased compared with healthy liver tissue.Citation183 To test whether restoration of miR-26a could have antitumorigenic effects, an adeno-associated virus vector packed with miR-26a was intravenously administered to liver tumorbearing mice. Three weeks after treatment, disease progression was dramatically decreased, cancer cell proliferation was inhibited, and tumor cell-specific apoptosis was increased compared with mice treated with empty vectors. The therapeutic effect of miRNA restoration was also shown for miR-34a, which was found to be underexpressed in prostate cancer cells.Citation184 Upon repetitive intravenous administration of miR-34a complexed with a lipid-based delivery agent to mice with prostate cancer xenografts, tumor growth and metastasis were inhibited and mouse survival was prolonged. These effects were likely due to miR-34a- mediated downregulation of the adhesion molecule CD44 in tumor cells.Citation184
While some miRNAs appear to be downregulated in various disease states, other miRNAs have been shown to be overexpressed. This phenomenon has supported the emergence of anti-miRNA-based therapeutics.Citation185 These are antisense oligonucleotides with sequences complementary to those of the targeted miRNA, which disrupt miRNA biogenesis and functioning at multiple fronts (eg, miRNA processing and interactions with the target mRNA).Citation186 Several interesting studies have effectively employed anti-miRNA strategies to induce cancer cell death and inhibit disease progression.Citation187–Citation191
These exciting results illustrate the potential of miRNAs and anti-miRNAs as a novel class of endogenous therapeutics. However, the intracellular delivery of (anti-)miRNAs is generally poor, and multiple (intratumoral) injections are often required for an effect.Citation192,Citation193 Given that exosomes are natural carriers for miRNAs, exosome mimetics would be excellent delivery systems for such molecules. Because their membrane resembles that of natural exosomes, the stability of encapsulated miRNAs could be increased. Furthermore, exosome mimetics may allow for nontoxic and nonimmunogenic delivery of their cargo to target cells, strongly potentiating the effects of (anti-)miRNAs.
Although miRNAs are promising therapeutic targets, it is important to note that they do not require full (100%) binding to their target mRNA sequences for inhibiting effects.Citation194 This allows them to act synergistically on various targets within signaling pathways, but also increases their chance for offtarget effects. These can be minimized by extensive characterization of possible target sequences and optimization of the administered miRNA doses. Alternatively, siRNA, which is known for its excellent sequence specificity,Citation195 may be used to silence the expression of specific target genes. Similar to miRNA, the properties of siRNA (eg, their anionic charge, size, and rapid clearance) call for packaging in adequate delivery systems. Given that biotechnologically engineered exosomes have previously been successfully applied for delivery of siRNA,Citation53 exosome mimetics hold great potential for siRNA-delivery.
Conclusion and perspectives
The safe and effective delivery of drug molecules to their target site is a field which has increasingly gained attention in drug design and development. In recent decades, the focus has shifted from synthetic drug compounds to the delivery of biological drugs (ie, proteins and nucleic acids), which are very prone to immune effects and degradation. In this regard, exosome mimetics are promising candidate delivery vehicles, given that they mimic nature’s delivery vehicles of biologicals, but are not as complex as their biological counterparts. These characteristics may allow them to deliver biologicals in an effective and safe manner, with high pharmaceutical acceptability due to their well characterized components. However, before the full potential of exosome mimetics can be exploited, a number of challenges still need to be overcome. The assembly of liposomes with varying sizes and lipid compositions is commonly performed and well understood, but the incorporation of functional proteins and nucleic acids with reasonable efficiency is a field on its own. Membrane proteins have been successfully incorporated in liposomes without losing their functionality,Citation108–Citation112 but the incorporation of multiple proteins is a time-consuming and complicated process. Their membrane localization may require a specific lipid composition and chaperone proteins. Moreover, the (recombinant) production of membrane proteins is a challenging task due to their high hydrophobicity and cytotoxicity, tendency to form aggregates, and low expression levels. The use of promising cell-free expression systems may aid to overcome these production issues.Citation173–Citation175
For assembly of functional exosome mimetics, it is also important to take into account the structural and biological functions of the lipids to be incorporated. Lipids come in various sizes and shapes, which influence the behavior of the liposome in biological environments (eg, fusion and stability). In this regard, it may be wise to borrow a leaf from nature’s book and mimic the lipid composition of exosomes in the intended target environment for optimal functionality. Increasingly powerful analytical methods in the field of lipidomics may aid in elucidating both function and composition of lipids in natural exosomes. These may provide valuable clues for drug delivery with exosome mimetics.
An additional challenge governing the assembly of functional exosome mimetics is the loading of therapeutic cargo. Although work is progressing in this area, high-efficiency loading of siRNA (and likewise miRNA) in lipid vesicles is still rarely achieved.Citation196,Citation197 Given that exosomes are loaded with multiple components that probably contribute to the outstanding delivery capacities of these vesicles, efficient loading of a combination of nucleic acids and proteins in exosome mimetics merits investigation. Furthermore, it is important to note that crucial components of exosomes are still largely unknown, and probably differ among exosomes with varying functions and target cells. The rapidly expanding field of exosomics allows us to increase our understanding of the composition and function of these intriguing vesicles, but is still in its infancy. Extensive proteomic studies have identified more than 4000 proteins and more than 1500 miRNAs in exosomes from various sources, but integrative studies are required to elucidate the biological functions of these components. The answer to “What makes natural exosomes such effective delivery vehicles?”, is the first step towards the design and assembly of functional exosome mimetics.
Acknowledgments
The work of SAAK, PV, SMvD, and RMS on cell-derived membrane vesicles is supported by a European Research Council starting grant (260627) “MINDS” in the FP7 ideas program of the European Union.
Disclosure
The authors report no conflicts of interest in this work.
References
- WolfPThe nature and significance of platelet products in human plasmaBr J Haematol19671332692886025241
- CrawfordNThe presence of contractile proteins in platelet microparticles isolated from human and animal platelet-free plasmaBr J Haematol197121153694254312
- DoonanFCotterTGMorphological assessment of apoptosisMethods200844320020418314050
- HristovMErlWLinderSWeberPCApoptotic bodies from endothelial cells enhance the number and initiate the differentiation of human endothelial progenitor cells in vitroBlood200410492761276615242875
- XieYBaiOYuanJTumor apoptotic bodies inhibit CTL responses and antitumor immunity via membrane-bound transforming growth factor-beta1 inducing CD8+ T-cell anergy and CD4+ Tr1 cell responsesCancer Res200969197756776619789353
- ClineAMRadicMZApoptosis, subcellular particles, and autoimmunityClin Immunol2004112217518215240161
- TheryCOstrowskiMSeguraEMembrane vesicles as conveyors of immune responsesNat Rev Immunol20099858159319498381
- SchillerMBekeredjian-DingIHeyderPBlankNHoADLorenzHMAutoantigens are translocated into small apoptotic bodies during early stages of apoptosisCell Death Differ200815118319117932498
- GregoryCDPoundJDMicroenvironmental influences of apoptosis in vivo and in vitroApoptosis20101591029104920237956
- Muralidharan-ChariVClancyJWSedgwickAD’Souza-SchoreyCMicrovesicles: mediators of extracellular communication during cancer progressionJ Cell Sci2010123Pt 101603161120445011
- PilzerDGasserOMoskovichOSchifferliJAFishelsonZEmission of membrane vesicles: roles in complement resistance, immunity and cancerSpringer Semin Immunopathol200527337538716189651
- SheddenKXieXTChandaroyPChangYTRosaniaGRExpulsion of small molecules in vesicles shed by cancer cells: association with gene expression and chemosensitivity profilesCancer Res200363154331433712907600
- SekulaMJanawaGStankiewiczEStepienEEndothelial microparticle formation in moderate concentrations of homocysteine and methionine in vitroCell Mol Biol Lett2011161697821161416
- ShantsilaEKamphuisenPWLipGYCirculating microparticles in cardiovascular disease: implications for atherogenesis and atherothrombosisJ Thromb Haemost20108112358236820695980
- Antwi-BaffourSKholiaSAryeeYKHuman plasma membrane-derived vesicles inhibit the phagocytosis of apoptotic cells – possible role in SLEBiochem Biophys Res Commun2010398227828320599722
- ObregonCRothen-RutishauserBGitahiSKGehrPNicodLPExovesicles from human activated dendritic cells fuse with resting dendritic cells, allowing them to present alloantigensAm J Pathol200616962127213617148675
- HugelBMartinezMCKunzelmannCFreyssinetJMMembrane microparticles: two sides of the coinPhysiology (Bethesda)200520222715653836
- BuckiRBachelot-LozaCZachowskiAGiraudFSulpiceJCCalcium induces phospholipid redistribution and microvesicle release in human erythrocyte membranes by independent pathwaysBiochemistry1998374415383153919799499
- GonzalezLJGibbonsEBaileyRWThe influence of membrane physical properties on microvesicle release in human erythrocytesPMC Biophys200921719703298
- SalzerUHinterdorferPHungerUBorkenCProhaskaRCa(++)-dependent vesicle release from erythrocytes involves stomatin-specific lipid rafts, synexin (annexin VII), and sorcinBlood20029972569257711895795
- CrawfordSDiamondDBrustolonLPenarretaREffect of increased extracellular ca on microvesicle production and tumor spheroid formationCancer Microenviron2010419310321505564
- AharonATamariTBrennerBMonocyte-derived microparticles and exosomes induce procoagulant and apoptotic effects on endothelial cellsThromb Haemost2008100587888518989533
- ThomasLMSalterRDActivation of macrophages by P2X7-induced microvesicles from myeloid cells is mediated by phospholipids and is partially dependent on TLR4J Immunol201018563740374920709956
- WileyJSSluyterRGuBJStokesLFullerSJThe human P2X7 receptor and its role in innate immunityTissue Antigens201178532133221988719
- RecordMSubraCSilvente-PoirotSPoirotMExosomes as intercellular signalosomes and pharmacological effectorsBiochem Pharmacol201181101171118221371441
- SimonsMRaposoGExosomes – vesicular carriers for intercellular communicationCurr Opin Cell Biol200921457558119442504
- TaylorDDGercel-TaylorCExosomes/microvesicles: mediators of cancer-associated immunosuppressive microenvironmentsSemin Immunopathol201133544145421688197
- SahooSKlychkoEThorneTExosomes from human CD34(+) stem cells mediate their proangiogenic paracrine activityCirc Res2011109772472821835908
- SokolovaVLudwigAKHornungSCharacterisation of exosomes derived from human cells by nanoparticle tracking analysis and scanning electron microscopyColloids Surf B Biointerfaces201187114615021640565
- SimpsonRJJensenSSLimJWProteomic profiling of exosomes: current perspectivesProteomics20088194083409918780348
- TheryCDubanLSeguraEVeronPLantzOAmigorenaSIndirect activation of naive CD4+ T cells by dendritic cell-derived exosomesNat Immunol20023121156116212426563
- MartinezMCAndriantsitohainaRMicroparticles in angiogenesis: therapeutic potentialCirc Res2011109111011921700952
- AharonABrennerBMicroparticles, thrombosis and cancerBest Pract Res Clin Haematol2009221616919285273
- ChoJAParkHLimEHExosomes from ovarian cancer cells induce adipose tissue-derived mesenchymal stem cells to acquire the physical and functional characteristics of tumor-supporting myofibroblastsGynecol Oncol2011123237938621903249
- Baj-KrzyworzekaMMajkaMPraticoDPlatelet-derived microparticles stimulate proliferation, survival, adhesion, and chemotaxis of hematopoietic cellsExp Hematol200230545045912031651
- Al-NedawiKMeehanBKerbelRSAllisonACRakJEndothelial expression of autocrine VEGF upon the uptake of tumor-derived microvesicles containing oncogenic EGFRProc Natl Acad Sci U S A2009106103794379919234131
- RatajczakJWysoczynskiMHayekFJanowska-WieczorekARatajczakMZMembrane-derived microvesicles: important and underappreciated mediators of cell-to-cell communicationLeukemia20062091487149516791265
- Del CondeIShrimptonCNThiagarajanPLopezJATissue-factor-bearing microvesicles arise from lipid rafts and fuse with activated platelets to initiate coagulationBlood200510651604161115741221
- TianTWangYWangHZhuZXiaoZVisualizing of the cellular uptake and intracellular trafficking of exosomes by live-cell microscopyJ Cell Biochem2010111248849620533300
- SarkarAMitraSMehtaSRaicesRWewersMDMonocyte derived microvesicles deliver a cell death message via encapsulated caspase-1PLoS One200949e714019779610
- ValadiHEkstromKBossiosASjostrandMLeeJJLotvallJOExosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cellsNat Cell Biol20079665465917486113
- MontecalvoALarreginaATShufeskyWJMechanism of transfer of functional microRNAs between mouse dendritic cells via exosomesBlood2012119375676622031862
- ArroyoJDChevilletJRKrohEMArgonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasmaProc Natl Acad Sci U S A2011108125003500821383194
- TurchinovichAWeizLLangheinzABurwinkelBCharacterization of extracellular circulating microRNANucleic Acids Res201139167223723321609964
- GuesciniMGenedaniSStocchiVAgnatiLFAstrocytes and glioblastoma cells release exosomes carrying mtDNAJ Neural Transm201011711419680595
- GuesciniMGuidolinDValloraniLC2C12 myoblasts release micro-vesicles containing mtDNA and proteins involved in signal transductionExp Cell Res2010316121977198420399774
- MitchellPSParkinRKKrohEMCirculating microRNAs as stable blood-based markers for cancer detectionProc Natl Acad Sci U S A200810530105131051818663219
- SkogJWurdingerTvan RijnSGlioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkersNat Cell Biol200810121470147619011622
- PegtelDMCosmopoulosKThorley-LawsonDAFunctional delivery of viral miRNAs via exosomesProc Natl Acad Sci U S A2010107146328633320304794
- CastanottoDRossiJJThe promises and pitfalls of RNA-interference-based therapeuticsNature2009457722842643319158789
- BaigudeHRanaTMDelivery of therapeutic RNAi by nanovehiclesChembiochem200910152449245419688785
- van den BoornJGSchleeMCochCHartmannGSiRNA delivery with exosome nanoparticlesNat Biotechnol201129432532621478846
- Alvarez-ErvitiLSeowYYinHBettsCLakhalSWoodMJDelivery of siRNA to the mouse brain by systemic injection of targeted exosomesNat Biotechnol201129434134521423189
- DelcayreAEstellesASperindeJExosome display technology: applications to the development of new diagnostics and therapeuticsBlood Cells Mol Dis200535215816816087368
- OtzenDEBlansKWangHGilbertGERasmussenJTLactadherin binds to phosphatidylserine-containing vesicles in a two-step mechanism sensitive to vesicle size and compositionBiochim Biophys Acta2012181841019102721920348
- ZeelenbergISOstrowskiMKrumeichSTargeting tumor antigens to secreted membrane vesicles in vivo induces efficient antitumor immune responsesCancer Res20086841228123518281500
- HartmanZCWeiJGlassOKIncreasing vaccine potency through exosome antigen targetingVaccine201129509361936722001882
- RountreeRBMandlSJNachtweyJMExosome targeting of tumor antigens expressed by cancer vaccines can improve antigen immunogenicity and therapeutic efficacyCancer Res201171155235524421670078
- EscudierBDorvalTChaputNVaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived-exosomes: results of the first phase I clinical trialJ Transl Med2005311015740633
- MorseMAGarstJOsadaTA phase I study of dexosome immunotherapy in patients with advanced non-small cell lung cancerJ Transl Med200531915723705
- SunDZhuangXXiangXA novel nanoparticle drug delivery system: the anti-inflammatory activity of curcumin is enhanced when encapsulated in exosomesMol Ther20101891606161420571541
- ZhuangXXiangXGrizzleWTreatment of brain inflammatory diseases by delivering exosome encapsulated anti-inflammatory drugs from the nasal region to the brainMol Ther201119101769177921915101
- OgawaraKRotsMGKokRJA novel strategy to modify adenovirus tropism and enhance transgene delivery to activated vascular endothelial cells in vitro and in vivoHum Gene Ther200415543344315144574
- KimWJKimSWEfficient siRNA delivery with non-viral polymeric vehiclesPharm Res200926365766619015957
- SinghaKNamgungRKimWJPolymers in small-interfering RNA deliveryNucleic Acid Ther201121313314721749290
- ClaytonAHarrisCLCourtJMasonMDMorganBPAntigen-presenting cell exosomes are protected from complement-mediated lysis by expression of CD55 and CD59Eur J Immunol200333252253112645951
- MeckesDGJrRaab-TraubNMicrovesicles and viral infectionJ Virol20118524128441285421976651
- CamussiGDeregibusMCBrunoSGrangeCFonsatoVTettaCExosome/microvesicle-mediated epigenetic reprogramming of cellsAm J Cancer Res2011119811021969178
- ChenTSArslanFYinYEnabling a robust scalable manufacturing process for therapeutic exosomes through oncogenic immortalization of human ESC-derived MSCsJ Transl Med201194721513579
- PuriALoomisKSmithBLipid-based nanoparticles as pharmaceutical drug carriers: from concepts to clinicCrit Rev Ther Drug Carrier Syst200926652358020402623
- FenskeDBCullisPRLiposomal nanomedicinesExpert Opin Drug Deliv200851254418095927
- MathivananSSimpsonRJExoCarta: a compendium of exosomal proteins and RNAProteomics20099214997500019810033
- MathivananSFahnerCJReidGESimpsonRJExoCarta 2012: database of exosomal proteins, RNA and lipidsNucleic Acids Res201240Database issueD1241124421989406
- SubraCGrandDLaulagnierKExosomes account for vesicle-mediated transcellular transport of activatable phospholipases and prostaglandinsJ Lipid Res20105182105212020424270
- LaulagnierKMottaCHamdiSMast cell- and dendritic cell-derived exosomes display a specific lipid composition and an unusual membrane organizationBiochem J2004380Pt 116117114965343
- WubboltsRLeckieRSVeenhuizenPTProteomic and biochemical analyses of human B cell-derived exosomes. Potential implications for their function and multivesicular body formationJ Biol Chem200327813109631097212519789
- AlonsoRMazzeoCRodriguezMCDiacylglycerol kinase alpha regulates the formation and polarisation of mature multivesicular bodies involved in the secretion of Fas ligand-containing exosomes in T lymphocytesCell Death Differ20111871161117321252909
- WhiteIJBaileyLMAghakhaniMRMossSEFutterCEEGF stimulates annexin 1-dependent inward vesiculation in a multivesicular endosome subpopulationEMBO J200625111216052208
- SubraCLaulagnierKPerretBRecordMExosome lipidomics unravels lipid sorting at the level of multivesicular bodiesBiochimie200789220521217157973
- RothSLWhittakerGRPromotion of vesicular stomatitis virus fusion by the endosome-specific phospholipid bis(monoacylglycero)phosphate (BMP)FEBS Lett2011585686586921333650
- ZaitsevaEYangSTMelikovKPourmalSChernomordikLVDengue virus ensures its fusion in late endosomes using compartment-specific lipidsPLoS Pathog2010610e100113120949067
- Le BlancILuyetPPPonsVEndosome-to-cytosol transport of viral nucleocapsidsNat Cell Biol20057765366415951806
- VidalMSainte-MarieJPhilippotJRBienvenueAAsymmetric distribution of phospholipids in the membrane of vesicles released during in vitro maturation of guinea pig reticulocytes: evidence precluding a role for “aminophospholipid translocase”J Cell Physiol198914034554622777884
- NeedhamDNunnRSElastic deformation and failure of lipid bilayer membranes containing cholesterolBiophys J199058499710092249000
- RamstedtBSlotteJPMembrane properties of sphingomyelinsFEBS Lett20025311333712401199
- Abi-RizkGBessonFInteractions of Triton X-100 with sphingomyelin and phosphatidylcholine monolayers: influence of the cholesterol contentColloids Surf B Biointerfaces200866216316718644701
- DrummondDCMeyerOHongKKirpotinDBPapahadjopoulosDOptimizing liposomes for delivery of chemotherapeutic agents to solid tumorsPharmacol Rev199951469174310581328
- RodriguezMAPytlikRKozakTVincristine sulfate liposomes injection (Marqibo) in heavily pretreated patients with refractory aggressive non-Hodgkin lymphoma: report of the pivotal phase 2 studyCancer2009115153475348219536896
- BoehlkeLWinterJNSphingomyelin/cholesterol liposomal vincristine: a new formulation for an old drugExpert Opin Biol Ther20066440941516548767
- AllenTMAustinGAChonnALinLLeeKCUptake of liposomes by cultured mouse bone marrow macrophages: influence of liposome composition and sizeBiochim Biophys Acta19911061156641995057
- AllenTMChonnALarge unilamellar liposomes with low uptake into the reticuloendothelial systemFEBS Lett1987223142463666140
- YokoyamaSTakedaTTsunodaTOhtaYImuraTAbeMMembrane properties of mixed dipalmitoylphosphatidylglycerol/ganglioside GM3 liposomes in the presence of bovine serum albuminColloids Surf B Biointerfaces2003272–3141146
- SchuttersKReutelingspergerCPhosphatidylserine targeting for diagnosis and treatment of human diseasesApoptosis20101591072108220440562
- AllenTMWilliamsonPSchlegelRAPhosphatidylserine as a determinant of reticuloendothelial recognition of liposome models of the erythrocyte surfaceProc Natl Acad Sci U S A19888521806780713186707
- PittoniVValesiniGThe clearance of apoptotic cells: implications for autoimmunityAutoimmun Rev20021315416112849009
- MorelOJeselLFreyssinetJMTotiFCellular mechanisms underlying the formation of circulating microparticlesArterioscler Thromb Vasc Biol2011311152621160064
- JeongJConboyIMPhosphatidylserine directly and positively regulates fusion of myoblasts into myotubesBiochem Biophys Res Commun2011414191321910971
- WuZNakanishiHPhosphatidylserine-containing liposomes: potential pharmacological interventions against inflammatory and immune diseases through the production of prostaglandin E(2) after uptake by myeloid derived phagocytesArch Immunol Ther Exp (Warsz)201159319520121479802
- MaxfieldFRMcGrawTEEndocytic recyclingNat Rev Mol Cell Biol20045212113215040445
- ChernomordikLVKozlovMMProtein-lipid interplay in fusion and fission of biological membranesAnnu Rev Biochem20037217520714527322
- PapahadjopoulosDNirSDuzgunesNMolecular mechanisms of calcium-induced membrane fusionJ Bioenerg Biomembr19902221571792139437
- HafezIMCullisPRRoles of lipid polymorphism in intracellular deliveryAdv Drug Deliv Rev2001472–313914811311989
- Al-NedawiKMeehanBMicallefJIntercellular transfer of the oncogenic receptor EGFRvIII by microvesicles derived from tumour cellsNat Cell Biol200810561962418425114
- MeckesDGJrShairKHMarquitzARKungCPEdwardsRHRaab-TraubNHuman tumor virus utilizes exosomes for intercellular communicationProc Natl Acad Sci U S A201010747203702037521059916
- MannoSTakakuwaYMohandasNIdentification of a functional role for lipid asymmetry in biological membranes: Phosphatidylserine-skeletal protein interactions modulate membrane stabilityProc Natl Acad Sci U S A20029941943194811830646
- ParoliniIFedericiCRaggiCMicroenvironmental pH is a key factor for exosome traffic in tumor cellsJ Biol Chem200928449342113422219801663
- NiemelaPSOllilaSHyvonenMTKarttunenMVattulainenIAssessing the nature of lipid raft membranesPLoS Comput Biol200732e3417319738
- AimonSManziJSchmidtDPoveda LarrosaJABassereauPToombesGEFunctional reconstitution of a voltage-gated potassium channel in giant unilamellar vesiclesPLoS One2011610e2552921998666
- ErbEMTangemannKBohrmannBMullerBEngelJIntegrin alphaIIb beta3 reconstituted into lipid bilayers is nonclustered in its activated state but clusters after fibrinogen bindingBiochemistry19973624739574029200686
- StreicherPNassoyPBarmannMIntegrin reconstituted in GUVs: a biomimetic system to study initial steps of cell spreadingBiochim Biophys Acta20091788102291230019665445
- ZhengMFangHTsuruokaTTsujiTSasakiTHakomoriSRegulatory role of GM3 ganglioside in alpha 5 beta 1 integrin receptor for fibronectin-mediated adhesion of FUA169 cellsJ Biol Chem19932683221722228420989
- VarnierAKermarrecFBlesneacIA simple method for the reconstitution of membrane proteins into giant unilamellar vesiclesJ Membr Biol20102331–3859220135103
- RaimondoFMorosiLChinelloCMagniFPittoMAdvances in membranous vesicle and exosome proteomics improving biological understanding and biomarker discoveryProteomics201111470972021241021
- TheryCRegnaultAGarinJMolecular characterization of dendritic cell-derived exosomes. Selective accumulation of the heat shock protein hsc73J Cell Biol1999147359961010545503
- EscolaJMKleijmeerMJStoorvogelWGriffithJMYoshieOGeuzeHJSelective enrichment of tetraspan proteins on the internal vesicles of multivesicular endosomes and on exosomes secreted by human B-lymphocytesJ Biol Chem19982733220121201279685355
- AtaySGercel-TaylorCKesimerMTaylorDDMorphologic and proteomic characterization of exosomes released by cultured extravillous trophoblast cellsExp Cell Res201131781192120221276792
- HemlerMETetraspanin functions and associated microdomainsNat Rev Mol Cell Biol200561080181116314869
- HemlerMETargeting of tetraspanin proteins – potential benefits and strategiesNat Rev Drug Discov20087974775818758472
- MiyadoKYamadaGYamadaSRequirement of CD9 on the egg plasma membrane for fertilizationScience2000287545132132410634791
- TachibanaIHemlerMERole of transmembrane 4 superfamily (TM4SF) proteins CD9 and CD81 in muscle cell fusion and myotube maintenanceJ Cell Biol1999146489390410459022
- MartinFRothDMJansDATetraspanins in viral infections: a fundamental role in viral biology?J Virol20057917108391085116103137
- Gordon-AlonsoMYanez-MoMBarreiroOTetraspanins CD9 and CD81 modulate HIV-1-induced membrane fusionJ Immunol200617785129513717015697
- TakedaYTachibanaIMiyadoKTetraspanins CD9 and CD81 function to prevent the fusion of mononuclear phagocytesJ Cell Biol2003161594595612796480
- ParthasarathyVMartinFHigginbottomADistinct roles for tetraspanins CD9, CD63 and CD81 in the formation of multinucleated giant cellsImmunology2009127223724819489128
- MantegazzaARBarrioMMMoutelSCD63 tetraspanin slows down cell migration and translocates to the endosomal-lysosomal-MIICs route after extracellular stimuli in human immature dendritic cellsBlood200410441183119015130945
- NazarenkoIRanaSBaumannACell surface tetraspanin Tspan8 contributes to molecular pathways of exosome-induced endothelial cell activationCancer Res20107041668167820124479
- HemlerMETetraspanin proteins mediate cellular penetration, invasion, and fusion events and define a novel type of membrane microdomainAnnu Rev Cell Dev Biol20031939742214570575
- ZollerMTetraspanins: push and pull in suppressing and promoting metastasisNat Rev Cancer200991405519078974
- van DommelenSMVaderPLakhalSMicrovesicles and exosomes: opportunities for cell-derived membrane vesicles in drug deliveryJ Control Release11252011 [Epub ahead of print.]
- BarczykMCarracedoSGullbergDIntegrinsCell Tissue Res2010339126928019693543
- CampbellIDHumphriesMJIntegrin structure, activation, and interactionsCold Spring Harb Perspect Biol201133a00499421421922
- HakulinenJSankkilaLSugiyamaNLehtiKKeski-OjaJSecretion of active membrane type 1 matrix metalloproteinase (MMP-14) into extracellular space in microvesicular exosomesJ Cell Biochem200810551211121818802920
- WeltonJLKhannaSGilesPJProteomics analysis of bladder cancer exosomesMol Cell Proteomics2010961324133820224111
- RieuSGeminardCRabesandratanaHSainte-MarieJVidalMExosomes released during reticulocyte maturation bind to fibronectin via integrin alpha4beta1Eur J Biochem2000267258359010632729
- ClaytonATurkesADewittSSteadmanRMasonMDHallettMBAdhesion and signaling by B cell-derived exosomes: the role of integrinsFASEB J200418997797915059973
- SeguraMMGarnierADi FalcoMRIdentification of host proteins associated with retroviral vector particles by proteomic analysis of highly purified vector preparationsJ Virol20088231107111718032515
- TemmingKSchiffelersRMMolemaGKokRJRGD-based strategies for selective delivery of therapeutics and imaging agents to the tumour vasculatureDrug Resist Updat20058638140216309948
- MeineckeRMeyerBDetermination of the binding specificity of an integral membrane protein by saturation transfer difference NMR: RGD peptide ligands binding to integrin alphaIIbbeta3J Med Chem200144193059306511543674
- ClaasenBAxmannMMeineckeRMeyerBDirect observation of ligand binding to membrane proteins in living cells by a saturation transfer double difference (STDD) NMR spectroscopy method shows a significantly higher affinity of integrin alpha(IIb)beta3 in native platelets than in liposomesJ Am Chem Soc2005127391691915656629
- GonzalesPAPisitkunTHoffertJDLarge-scale proteomics and phosphoproteomics of urinary exosomesJ Am Soc Nephrol200920236337919056867
- Gonzalez-BegneMLuBHanXProteomic analysis of human parotid gland exosomes by multidimensional protein identification technology (MudPIT)J Proteome Res2009831304131419199708
- PisitkunTShenRFKnepperMAIdentification and proteomic profiling of exosomes in human urineProc Natl Acad Sci U S A200410136133681337315326289
- LoozeCYuiDLeungLProteomic profiling of human plasma exosomes identifies PPARgamma as an exosome-associated proteinBiochem Biophys Res Commun2009378343343819028452
- BardMPHegmansJPHemmesAProteomic analysis of exosomes isolated from human malignant pleural effusionsAm J Respir Cell Mol Biol200431111412114975938
- MathivananSLimJWTauroBJJiHMoritzRLSimpsonRJProteomics analysis of A33 immunoaffinity-purified exosomes released from the human colon tumor cell line LIM1215 reveals a tissue-specific protein signatureMol Cell Proteomics20109219720819837982
- JohnASRothmanVLTuszynskiGPThrombospondin-1 (TSP-1) stimulates expression of integrin alpha6 in human breast carcinoma cells: a downstream modulator of TSP-1-induced cellular adhesionJ Oncol2010201064537620631908
- NuceraCPorrelloAAntonelloZAB-Raf(V600E) and thrombospondin-1 promote thyroid cancer progressionProc Natl Acad Sci U S A201010723106491065420498063
- Rivera-FillatMPReigFMartinezEMGrau-OlieteMRImproved therapeutic responses for liposomal doxorubicin targeted via thrombospondin peptidomimetics versus untargeted doxorubicinJ Pept Sci201016731532120552567
- KazerounianSYeeKOLawlerJThrombospondins in cancerCell Mol Life Sci200865570071218193162
- ZhangXLawlerJThrombospondin-based antiangiogenic therapyMicrovasc Res2007742–3909917559888
- SkokosDLe PanseSVillaIMast cell-dependent B and T lymphocyte activation is mediated by the secretion of immunologically active exosomesJ Immunol2001166286887611145662
- SkokosDLe PanseSVillaINonspecific B and T cell-stimulatory activity mediated by mast cells is associated with exosomesInt Arch Allergy Immunol20011241–313313611306949
- BuschowSIvan BalkomBWAalbertsMHeckAJWaubenMStoorvogelWMHC class II-associated proteins in B-cell exosomes and potential functional implications for exosome biogenesisImmunol Cell Biol201088885185620458337
- LongEOICAM-1: getting a grip on leukocyte adhesionJ Immunol201118695021502321505213
- SvajgerUAnderluhMJerasMObermajerNC-type lectin DC-SIGN: an adhesion, signalling and antigen-uptake molecule that guides dendritic cells in immunityCell Signal201022101397140520363321
- SeguraENiccoCLombardBICAM-1 on exosomes from mature dendritic cells is critical for efficient naive T-cell primingBlood2005106121622315790784
- SeguraEGuerinCHoggNAmigorenaSTheryCCD8+ dendritic cells use LFA-1 to capture MHC-peptide complexes from exosomes in vivoJ Immunol200717931489149617641014
- ChoiDSLeeJMParkGWProteomic analysis of microvesicles derived from human colorectal cancer cellsJ Proteome Res20076124646465517956143
- KesimerMScullMBrightonBCharacterization of exosome-like vesicles released from human tracheobronchial ciliated epithelium: a possible role in innate defenseFASEB J20092361858186819190083
- BrodbeckWGMoldCAtkinsonJPMedofMECooperation between decay-accelerating factor and membrane cofactor protein in protecting cells from autologous complement attackJ Immunol200016573999400611034410
- RollinsSASimsPJThe complement-inhibitory activity of CD59 resides in its capacity to block incorporation of C9 into membrane C5b-9J Immunol19901449347834831691760
- LiuSIshidaTKiwadaHCharacterization of bovine serum factor triggering the lysis of liposomes via complement activationBiol Pharm Bull19982143903979586579
- BradleyAJDevineDVAnsellSMJanzenJBrooksDEInhibition of liposome-induced complement activation by incorporated poly(ethylene glycol)-lipidsArch Biochem Biophys199835721851949735159
- FevrierBViletteDArcherFCells release prions in association with exosomesProc Natl Acad Sci U S A2004101269683968815210972
- PotolicchioICarvenGJXuXProteomic analysis of microglia-derived exosomes: metabolic role of the aminopeptidase CD13 in neuropeptide catabolismJ Immunol200517542237224316081791
- JiHErfaniNTauroBJDifference gel electrophoresis analysis of Ras-transformed fibroblast cell-derived exosomesElectrophoresis200829122660267118494037
- AndersenMHGraversenHFedosovSNPetersenTERasmussenJTFunctional analyses of two cellular binding domains of bovine lactadherinBiochemistry200039206200620610821695
- LinLHuaiQHuangMFurieBFurieBCCrystal structure of the bovine lactadherin C2 domain, a membrane binding motif, shows similarity to the C2 domains of factor V and factor VIIIJ Mol Biol2007371371772417583728
- MorelliAELarreginaATShufeskyWJEndocytosis, intracellular sorting, and processing of exosomes by dendritic cellsBlood2004104103257326615284116
- FensMHMastrobattistaEde GraaffAMAngiogenic endothelium shows lactadherin-dependent phagocytosis of aged erythrocytes and apoptotic cellsBlood200811194542455018292292
- QiangXLiJWuRExpression and characterization of recombinant human milk fat globule-EGF factor VIIIInt J Mol Med20112861071107621874225
- ShenBWuNYangJMGouldSJProtein targeting to exosomes/microvesicles by plasma membrane anchorsJ Biol Chem201128616143831439521300796
- KlammtCSchwarzDLohrFSchneiderBDotschVBernhardFCell-free expression as an emerging technique for the large scale production of integral membrane proteinFEBS J2006273184141415316930130
- ReckelSSobhanifarSDurstFStrategies for the cell-free expression of membrane proteinsMethods Mol Biol201060718721220204858
- SchwarzDDotschVBernhardFProduction of membrane proteins using cell-free expression systemsProteomics20088193933394618763710
- HovijitraNTWuuJJPeakerBSwartzJRCell-free synthesis of functional aquaporin Z in synthetic liposomesBiotechnol Bioeng20091041404919557835
- KanedaMNomuraSMIchinoseSDirect formation of proteoliposomes by in vitro synthesis and cellular cytosolic delivery with connexin-expressing liposomesBiomaterials20093023–243971397719423159
- MalamYLoizidouMSeifalianAMLiposomes and nanoparticles: nanosized vehicles for drug delivery in cancerTrends Pharmacol Sci2009301159259919837467
- TarahovskyYS“Smart” liposomal nanocontainers in biology and medicineBiochemistry (Mosc)201075781182420673204
- WangHXKolesnikovaTVDenisonCGygiSPHemlerMEThe C-terminal tail of tetraspanin protein CD9 contributes to its function and molecular organizationJ Cell Sci2011124Pt 162702271021771881
- BartelsCLTsongalisGJMicroRNAs: novel biomarkers for human cancerClin Chem200955462363119246618
- HenryJCAzevedo-PoulyACSchmittgenTDmicroRNA replacement therapy for cancerPharm Res201128123030304221879389
- KotaJChivukulaRRO’DonnellKATherapeutic microRNA delivery suppresses tumorigenesis in a murine liver cancer modelCell200913761005101719524505
- LiuCKelnarKLiuBThe microRNA miR-34a inhibits prostate cancer stem cells and metastasis by directly repressing CD44Nat Med201117221121521240262
- GambariRFabbriEBorgattiMTargeting microRNAs involved in human diseases: a novel approach for modification of gene expression and drug developmentBiochem Pharmacol201182101416142921864506
- KotaSKBalasubramanianSCancer therapy via modulation of micro RNA levels: a promising futureDrug Discov Today20101517–1873374020692360
- MatsubaraHTakeuchiTNishikawaEApoptosis induction by antisense oligonucleotides against miR-17-15p and miR-20a in lung cancers overexpressing miR-17–92Oncogene200726416099610517384677
- SeguraMFHannifordDMenendezSAberrant miR-182 expression promotes melanoma metastasis by repressing FOXO3 and microphthalmia-associated transcription factorProc Natl Acad Sci U S A200910661814181919188590
- SiMLZhuSWuHLuZWuFMoYYmiR-21-mediated tumor growthOncogene200726192799280317072344
- SylvestreYDe GuireVQueridoEAn E2F/miR-20a autoregulatory feedback loopJ Biol Chem200728242135214317135249
- LuYRoySNuovoGAnti-microRNA-222 (anti-miR-222) and -181B suppress growth of tamoxifen-resistant xenografts in mouse by targeting TIMP3 protein and modulating mitogenic signalJ Biol Chem201128649422924230222009755
- YangYPChienYChiouGYInhibition of cancer stem cell-like properties and reduced chemoradioresistance of glioblastoma using microRNA145 with cationic polyurethane-short branch PEIBiomaterials20123351462147622098779
- XuDTakeshitaFHinoYmiR-22 represses cancer progression by inducing cellular senescenceJ Cell Biol2011193240942421502362
- KimDHRossiJJStrategies for silencing human disease using RNA interferenceNat Rev Genet20078317318417304245
- GuoPCobanOSneadNMEngineering RNA for targeted siRNA delivery and medical applicationAdv Drug Deliv Rev201062665066620230868
- SzokaFMolecular biology. The art of assemblyScience2008319586357857918239114
- LakhalSWoodMJExosome nanotechnology: an emerging paradigm shift in drug delivery: exploitation of exosome nanovesicles for systemic in vivo delivery of RNAi heralds new horizons for drug delivery across biological barriersBioessays2011331073774121932222