
Abstract
Recent studies have shown that approximately 70% of patients with severe atopic dermatitis (AD) develop asthma. Development of AD in infancy and subsequent other atopic diseases such as asthma in childhood is referred to as atopic march. However, a causal link between the diseases of atopic march has remained largely unaddressed, possibly due to lack of a proper animal model. Recently, we developed an AD rat model showing chronically relapsing dermatitis and scratching behaviors induced by neonatal capsaicin treatment. Here, we investigated whether our model also showed asthmatic changes, with the aim of expanding our AD model into an atopic march model. First, we confirmed that capsaicin treatment (50 mg/kg within 24 h after birth) induced dermatitis and scratching behaviors until 6 weeks of age. After that, the mRNA expression of Th1 and Th2 cytokines, such as IFN-γ and TNF-α, and IL-4, IL-5, and IL-13, respectively, was quantified with quantitative real-time polymerase chain reaction in the skin and the lungs. The number of total cells and eosinophils was counted in bronchoalveolar lavage (BAL) fluid. The levels of IgE in the serum and BAL fluid were determined with enzyme-linked immunosorbent assay. Paraffin-embedded sections (4 μm) were stained with hematoxylin/eosin to analyze the morphology of the lung and the airway. Airway responsiveness was measured in terms of airway resistance and compliance using the flexiVent system. In the capsaicin-treated rats, persistent dermatitis developed, and scratching behaviors increased over several weeks. The levels of IgE in the serum and BAL fluid as well as the mRNA expression of Th2 cytokines, including IL-4, IL-5, and IL-13, in both the skin and the lungs were elevated, and the number of eosinophils in the BAL fluid was also increased in the capsaicin-treated rats compared to control rats. Morphological analysis of the airway revealed smooth muscle hypertrophy and extensive mucus plug in the capsaicin-treated rats. Functional studies demonstrated an increment of the airway resistance and a decrement of lung compliance in the capsaicin-treated rats compared to control rats. Taken together, our findings suggested that neonatal capsaicin treatment induced asthma-like airway inflammation and responses in juvenile rats.
Introduction
Atopic dermatitis (AD) in infants frequently progresses to asthma and allergic rhinitis in childhood. This progression is known as atopic march. In fact, more than 50% of patients with moderate-to-severe AD develop asthma, while 8% of the people without AD suffer from asthma.Citation1,Citation2 The development of various atopic diseases shares complex and heterogeneous, but common pathogenesis including increment in IgE production, enhanced Th2 allergic reactions, and impaired barrier function.Citation2 Barrier dysfunction and subsequent early life exposure to allergen are considered as triggers for atopic march.Citation2,Citation3 However, we still lack a detailed picture of the progression of atopic diseases from AD to asthma.
One of the major problems with investigating the link among atopic diseases is probably the lack of a proper animal model. Some research groups have developed animal models induced by repeated allergic sensitization of ovalbumin through impaired skin barriers, followed by intranasal exposure, for the purpose of studying the progression of atopic diseases.Citation4,Citation5
Recently, we developed a rat model of AD with chronically relapsing dermatitis and scratching behaviors induced by neonatal capsaicin treatment. In the previous study, we showed how neonatal capsaicin treatment interfered with epithelial barrier function, which possibly resulted in the development of chronic pruritic cutaneous lesions.Citation6 Thus, we could infer a link between AD and asthma from epithelial barrier dysfunction. In fact, Long et alCitation7 reported that chronic expsoure of SO2 simutaneously with neonatal capsaicin treatment altered airway responsiveness in adult rats.
Here, we proposed that the AD model produced by neonatal capsaicin treatment also showed asthmatic changes such as Th2 allergic inflammation in the airway, airway remodeling, and airway hyperresponsiveness (AHR).
Materials and methods
Animals
All experiments were approved by the Institutional Animal Care and Use Committee in College of Medicine, Korea University (KOREA-2016-0024), and animals were maintained in the facility compliant with the Korean Ministry of Food and Drug Safety Standard Guideline based on Guide for the Care and Use of Laboratory Animals (8th edition). Pregnant Sprague Dawley rats (Samtako, South Korea) were acquired 2 or 3 days before parturition. Newborns were weaned at postnatal week 3, and only the males were used. All animals were reared in a room maintained at a 12-h light/dark cycle (light on at 7:00 AM) at 22–25°C and had ad libitum access to food and water.
Neonatal capsaicin treatment
As previously described,Citation6 neonatal rats were injected subcutaneously with capsaicin (capsaicin-treated group, 50 mg/kg; Sigma) or vehicle (control group, saline containing 10% Tween 80 and 10% ethanol) at a volume of 10 ml/g body weight within 24 h of birth.
Evaluation of pruritus
Rats were placed in separate plastic chambers (20×30×20 cm) containing a mirror for prevention of a blinded view. The scratching behaviors were recorded for an hour using a digital video camera (HDR-CX380; Sony, Tokyo, Japan) every week for 6 weeks. The video files were played back, and the number of scratches was assessed by blind experimenters unaware of the allocation of treatments. A bout of consecutive scratching strokes was regarded as one scratch (Video S1).
Evaluation of cutaneous lesions
Cutaneous lesions were carefully appraised using the score systemCitation8 with modifications. The skin lesions in three different regions (the ears, face, and back) were assessed every week for 6 weeks to evaluate the severity and extent of lesions. The unit size for the measurement of cutaneous lesions was 0.25 cm2, and the dermatitis score was determined by the unit size multiplied by the severity index, which is defined in .
Table 1 Severity index for the dermatitis score
Sampling of the tissues
After behavioral testing for 6 weeks, rats were sacrificed using CO2 gas. After that, lesional skin in the middle of the back of the neck (or the nearest lesional part) was acquired from the capsaicin-treated rats. The non-lesional skin in the same region was obtained from the control rats. Blood was collected with cardiac puncture using a 1 ml syringe, and then both the lobes of the lung were acquired for histological analysis and quantitative real-time polymerase chain reaction (RT-PCR). All samples were stored at −70°C and used within 2 weeks of collection.
Enzyme-linked immunosorbent assay (ELISA)
The IgE levels of serum and bronchoalveolar lavage (BAL) fluid were determined using a rat IgE ELISA Kit (CUSA-BIO), according to the manufacturer’s protocol.
Quantitative RT-PCR
The tissues were homogenized, and RNA was obtained using TRIzol Reagent (Thermo Fisher Scientific). Synthesis of cDNA was performed using M-MLV reverse transcriptase (Thermo Fisher Scientific). Quantitative PCR (qPCR) was performed in duplicate using the GoTaq qPCR Master Mix (Promega) on a Roche LightCycler 96 (Roche). Primers are described in .
Table 2 Primers used in quantitative real-time polymerase chain reaction
Bronchoalveolar lavage
After the rats in both the groups were sacrificed, the airway was cannulated with a 16-gauge needle and lavaged once with 5 ml of phosphate-buffered saline (PBS). The fluid collected from airway lavage was defined as BAL fluid, and was centrifuged, and the supernatant was used for ELISA. The cell pellet was treated with red blood cell lysis buffer (eBioscience) and washed in PBS. The resultant cells were fixed with methyl alcohol and stained with hematoxylin and eosin. The number of total cells was counted in duplicate with a hemocytometer. More than 250 cells were used to determine the differential cell count.
Lung histology
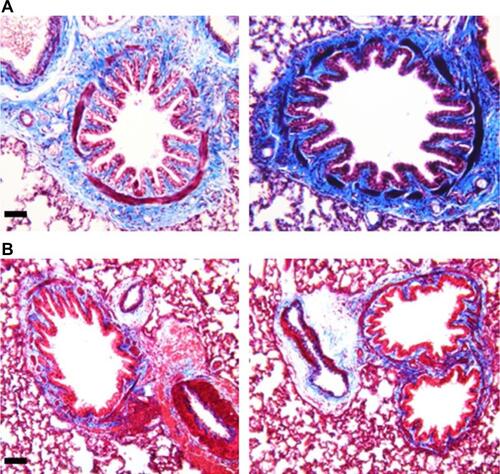
The lungs were removed, washed in PBS, and then fixed in 10% neutral buffered formalin solution (Sigma). After fixation, they were embedded in paraffin, and lung sections were cut into 4-μm samples. Slides were stained either with hematoxylin and eosin or Masson’s trichrome (Trichrome III Blue Staining Kit; Ventana Medical Systems), and more than three different sections of each lung were evaluated under light microscopy. Only the airways whose lumen diameter was from 50 to 200 μm were included in the evaluation.
For evaluating airway remodeling, we measured the amount of mucus plug and the extent of smooth muscle hypertrophy. First, to assess the amount of mucus plug in the airways, we adopted a scoring system, and the size of the area was measured using Image J (NIH). The sum of the scores from each airway of all the slides was taken as the mucus plug score in arbitrary units, which is shown in . Next, to calculate smooth muscle hypertrophy, we considered (A/C)*(B/C) as smooth muscle thickness ratio (). The largest width of smooth muscle in the section was considered as the smooth muscle thickness (A). The major axis of the airway lumen (C) and the minor axis of the airway lumen (B) were used. We divided A by C to correct the bias from the size of the lumen, and (B/C) was multiplied to compensate the bias from the tilt angle of the section. The sum of the smooth muscle thickness ratio from each airway was divided by the number of airways examined, and stated in arbitrary units. If B/C was smaller than 2/3, the ratio was dropped from the calculation to exclude angled sections.
Figure 1 Schematic representation of the airway lumen and surrounding smooth muscle.
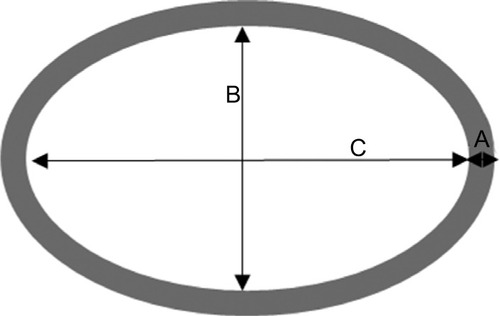
Table 3 Scoring system for measuring the amount of mucus plug in the airway
Measurement of airway responsiveness
AHR to methacholine (acetyl-b-methylcholine chloride; Sigma) was evaluated in rats of 6 weeks of age using a computer-controlled small-animal ventilator (flexiVent; SCIREQ). Rats were anesthetized with sodium pentobarbital (50 mg/kg) via intraperitoneal injection. The rats were intubated with a 16-gauge metallic needle and connected to flexiVent machine. Lung mechanics (airway resistance and compliance) were measured in every rat after nebulization of PBS for the baseline using an ultrasonic nebulizer (Aeroneb; SCIREQ). After measurement of the baseline, a methacholine challenge test was conducted. Rats were challenged with methacholine at the following doses: 6.25, 12.5, 25, and 50 mg/ml. Then, the rats were ventilated in a constant-flow inflation mode with a tidal volume of 10 ml/kg, an inspiratory–expiratory ratio of 2:3, a respiratory rate of 120 breaths/min, and maximum pressure of 30 cm H2O. Positive end-expiratory pressure was maintained by submerging the expiratory limb in a water trap.
Statistical analysis
Results were expressed as mean ± standard error of the mean. Data were analyzed using the Student’s t-test or the Mann–Whitney rank-sum test, depending on results from the normality test.
Results
AD-like symptoms and amplification of Th2-dominant immune response
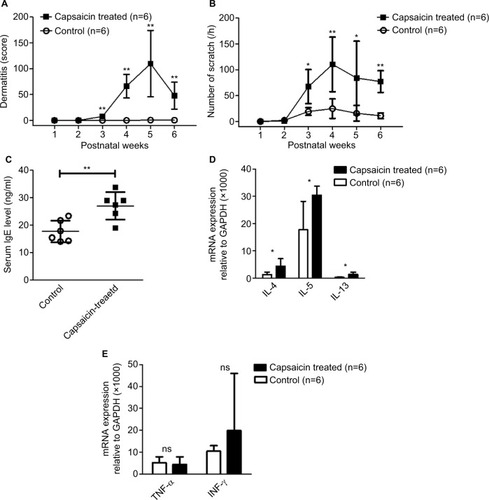
To confirm whether neonatal capsaicin treatment caused AD-like symptoms in rats, we examined cutaneous lesions and scratching behaviors for 6 weeks. Dermatitis first appeared at 3 weeks of age and became aggravated over the following weeks in the capsaicin-treated rats unlike the control rats (). At 3 weeks of age, the number of scratches also started to increase significantly in the capsaicin-treated rats compared to the control rats and persisted until 6 weeks of age (). Serum IgE level was remarkably higher in the capsaicin-treated rats than in the control rats at 6 weeks of age (). To determine whether the skin inflammation was attributed to enhanced Th2 allergic response, we investigated cytokine production in the skin. At 6 weeks of age, mRNA expression of Th2 cytokines such as IL-4, IL-5, and IL-13 was markedly elevated in the skin of the capsaicin-treated rats compared to the control rats (), but mRNA expression of Th1 cytokines such as TNF-α and IFN-γ showed no significant alteration ().
Figure 2 Atopic dermatitis-like skin inflammation, pruritus, and amplification of Th2 inflammatory reactions.
Eosinophil infiltration in the airway and increment of mRNA expression of Th2 cytokines in the lung
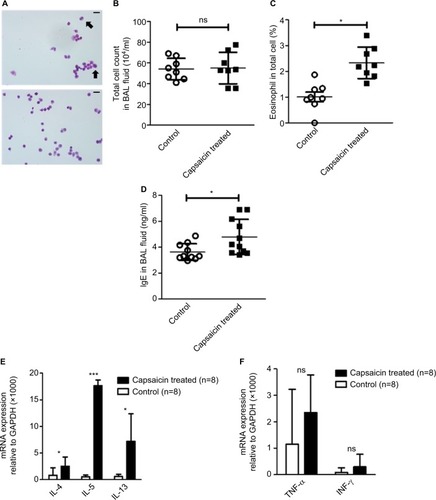
To investigate asthmatic progression in the airway and the lungs, we examined the total cell count, differential cell count, and IgE levels in BAL fluid. Although no significant difference in the number of leukocytes was observed between the capsaicin-treated rats and the control rats (), the percentage of eosinophils in the total cell count was significantly higher in the capsaicin-treated rats compared to control rats (). The IgE level in BAL fluid was also remarkably higher in the capsaicin-treated rats than in control rats (). To investigate whether Th2 allergic inflammation was as enhanced in the lung as in the skin, the mRNA expression of Th2 and Th1 cytokines was quantified. Similar to the skin, mRNA expression of Th2 cytokines such as IL-4, IL-5, and IL-13 was higher in the lung of the capsaicin-treated rats compared to control rats (), even though no remarkable difference was recorded for the mRNA expression of Th1 cytokines such as TNF-α and IFN-γ ().
Figure 3 Eosinophil infiltration and elevated expression of Th2 cytokines.
Abbreviations: BAL, bronchoalveolar lavage; ns, non significant.
Airway remodeling and AHR
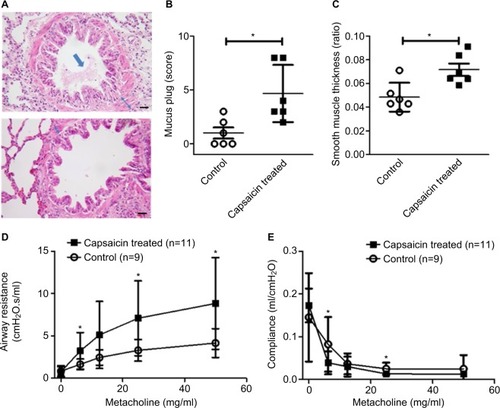
To determine concurrent airway remodeling, we analyzed the histological features of the airway. Morphological analysis revealed extensive mucus plug and smooth muscle hypertrophy (). The amount of mucus plug expressed was markedly higher in the capsaicin-treated rats than in the control rats (). The ratio of smooth muscle thickness, indicative of smooth muscle hypertrophy, was significantly higher in capsaicin-treated rats compared to control rats (). Furthermore, collagen deposition around airways was also increased in capsaicin-treated rats (Figure S1A) compared to control rats (Figure S1B). Airway resistance in response to methacholine was remarkably higher in the capsaicin-treated rats than in the control rats in a dose-dependent fashion, with the exception of rats exposed to 12.5 mg/ml methacholine (). Lung compliance in response to methacholine was also much lower in the capsaicin-treated rats than in the control rats at doses of 6.25 and 25 mg/ml, but not with 12.5 and 50 mg/ml (). Furthermore, even at the doses showing a lack of statistical significance, alteration of airway resistance and compliance demonstrated the same tendency.
Figure 4 Airway remodeling and airway hyperresponsiveness.
Discussion
We developed a rat model of AD induced by neonatal capsaicin treatment.Citation6 In the present study, the model showed not only AD-like symptoms but also asthmatic changes such as elevated levels of IgE, Th2 allergic inflammation, airway remodeling, and AHR in adolescent period. Taken together, this model could be useful for studying the progression of atopic diseases.
As previously described,Citation6 the capsaicin-treated rats developed chronically relapsing dermatitis and pruritus starting at 3 weeks of age. The expression pattern of inflammatory cytokines in supports the idea that allergic reactions of Th2, but not Th1, played a pivotal role in the clinical manifestation of AD-like symptoms in the capsaicin-treated rats. Elevated serum IgE level in the capsaicin-treated rats was also consistent with the feature of atopic diseases as AD and asthma are almost invariably accompanied by elevated levels of IgE.Citation9
In accordance with the results in , human studies provided evidence that the number of eosinophils increased in patients with asthma, while the total cell count of asthma patients did not differ from controls.Citation10,Citation11 In addition, IgE production and Th2-associated immune responses are features of asthmatics.Citation12,Citation13 Crimi et alCitation14 reported that total IgE/K+ ratio in bronchial lavage was higher in asthmatics than in controls, which was consistent with the results in . Inflammatory Th2 cytokines such as IL-4 and IL-13 released from eosinophils contributed to asthmatic symptoms.Citation15 Furthermore, Th2 cells promoted allergic reactions in asthmatics by amplifying production of cytokines including IL-4, IL-5, and IL-13, which resulted in IgE production, along with AHR and tissue remodeling.Citation16,Citation17 Thus, the elevated mRNA expression of Th2 cytokines shown in possibly contributed to the airway remodeling and asthmatic symptoms in the capsaicin-treated rats.
The complex interplay between aberrant immune response and chronic inflammation leads to the progression of airway remodeling in asthmatics.Citation18,Citation19 Airway smooth muscle hypertrophy has been accepted as a distinctive feature of asthmatic remodeling, though there are controversial reports regarding the degree of hypertrophy.Citation20,Citation21 Furthermore, the airway of asthma patients is characterized by hyperproduction of mucus as well as persistent inflammation and eosinophil infiltration.Citation22–Citation24 Histological analysis of the airway of the capsaicin-treated rats revealed asthmatic features such as smooth muscle hypertrophy and increased mucus plug in agreement with previous reports. AHR is defined as the aggravation of airway obstruction in response to bronchoconstrictors and is a diagnostic symptom of asthma. Progressive increment of airway resistance at increasing doses of methacholine is regarded as AHR in rodents. Moreover, when significant airflow obstruction occurs, compliance decreases substantially during tidal breath in asthma.Citation25 Increased airway resistance and decreased compliance, shown in , run parallel to the characteristic airway responsiveness of asthma.
AD and asthma share a common pathogenesis and immunologic features including elevated IgE, overproduction of Th2 cytokines, and barrier dysfunction.Citation2 Several studies provided evidence that sensitization through a disrupted skin barrier enhanced the Th2 cytokine expressionCitation26 and subsequent migration of Th2 cells through the circulatory system promoted an allergic response in the airways.Citation27 We previously demonstrated the impairment of skin barrier function in our model,Citation6 which was plausibly attributed to inflammatory reactions due to neuronal cell death by neonatal capsaicin treatment.Citation28 Overexpression of inflammatory mediators including IL-13 contributed to the induction of atopic diseases such as AD in rodents even without deliberate allergen exposure.Citation29,Citation30 Thus, such barrier dysfunction and inflammatory response possibly facilitated spontaneous early-life allergen sensitization, which contributed to the development of asthma as well as AD.Citation31 However, the role of inflammation or neuronal cell death in the pathogenesis of allergic diseases through a damaged barrier in our model needs to be confirmed.
In summary, our results suggest that our model induced by neonatal capsaicin treatment could be suitable for the study of atopic march from the perspective of clinical manifestation and intrinsic pathogenesis.
Acknowledgments
This research was supported by the Public Welfare & Safety research program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (NRF-2014M3C8A5030612), and supported by the Chung Yang, Cha Young Sun, M.D. and Jang Hi Joo Yeu Sa Memorial Fund.
Supplementary materials
Figure S1 Increased collagen deposition in the airways.
Note: Representative slides revealed relative abundance of collagen deposition in the airways from the capsaicin-treated rats (A) compared to control rats (B) (scale bar=50 μm).
Disclosure
The authors report no conflicts of interest in this work.
References
- HahnELBacharierLBThe atopic march: the pattern of allergic disease development in childhoodImmunol Allergy Clin North Am2005252231246v15878453
- SpergelJMPallerASAtopic dermatitis and the atopic marchJ Allergy Clin Immunol20031126 SupplS118S12714657842
- Biagini MyersJMKhurana HersheyGKEczema in early life: genetics, the skin barrier, and lessons learned from birth cohort studiesJ Pediatr2010157570471420739029
- Leyva-CastilloJMHenerPJiangHLiMTSLP produced by kerati-nocytes promotes allergen sensitization through skin and thereby triggers atopic march in miceJ Invest Dermatol2013133115416322832486
- SpergelJMMizoguchiEBrewerJPMartinTRBhanAKGehaRSEpicutaneous sensitization with protein antigen induces localized allergic dermatitis and hyperresponsiveness to methacholine after single exposure to aerosolized antigen in miceJ Clin Invest19981018161416229541491
- BackSKJeongKYLiCLeeJLeeSBNaHSChronically relapsing pruritic dermatitis in the rats treated as neonate with capsaicin; a potential rat model of human atopic dermatitisJ Dermatol Sci201267211111922721998
- LongNCMartinJGPantanoRShoreSAAirway hyperresponsiveness in a rat model of chronic bronchitis: role of C fibersAm J Respir Crit Care Med19971554122212299105058
- MiharaKKurataniKMatsuiTNakamuraMYokotaKVital role of the itch-scratch response in development of spontaneous dermatitis in NC/Nga miceBr J Dermatol2004151233534515327540
- OettgenHCGehaRSIgE in asthma and atopy: cellular and molecular connectionsJ Clin Invest1999104782983510510320
- KellyCWardCStentonCSBirdGHendrickDJWaltersEHNumber and activity of inflammatory cells in bronchoalveolar lavage fluid in asthma and their relation to airway responsivenessThorax19884396846923194874
- SmithDLDeshazoRDBronchoalveolar lavage in asthma. An update and perspectiveAm Rev Respir Dis199314825235328342920
- WilsonDRMerrettTGVargaEMIncreases in allergen-specific IgE in BAL after segmental allergen challenge in atopic asthmaticsAm J Respir Crit Care Med20021651222611779725
- MartinezFDVercelliDAsthmaLancet201338299011360137224041942
- CrimiEScordamagliaACrimiPZupoSBarocciSTotal and specific IgE in serum, bronchial lavage and bronchoalveolar lavage of asthmatic patientsAllergy19833885535596660434
- FrigasEGleichGJThe eosinophil and the pathophysiology of asthmaJ Allergy Clin Immunol19867745275373514730
- McGeeHSAgrawalDKTH2 cells in the pathogenesis of airway remodeling: regulatory T cells a plausible panacea for asthmaImmunol Res200635321923217172648
- DaviesDEWicksJPowellRMPuddicombeSMHolgateSTAirway remodeling in asthma: new insightsJ Allergy Clin Immunol20031112215225 quiz 22612589337
- LarchéMRobinsonDSKayABThe role of T lymphocytes in the pathogenesis of asthmaJ Allergy Clin Immunol20031113450463 quiz 46412642820
- HomerRJEliasJAAirway remodeling in asthma: therapeutic implications of mechanismsPhysiology (Bethesda)200520283515653837
- BenayounLDruilheADombretMCAubierMPretolaniMAirway structural alterations selectively associated with severe asthmaAm J Respir Crit Care Med2003167101360136812531777
- WoodruffPGDolganovGMFerrandoREHyperplasia of smooth muscle in mild to moderate asthma without changes in cell size or gene expressionAm J Respir Crit Care Med200416991001100614726423
- HolgateSTWilsonJRHowarthPHNew insights into airway inflammation by endobronchial biopsyAm Rev Respir Dis19921452 Pt 2S2S61734808
- KimKCMcCrackenKLeeBCAirway goblet cell mucin: its structure and regulation of secretionEur Respir J19971011264426499426108
- KupermanDAHuangXKothLLDirect effects of interleukin-13 on epithelial cells cause airway hyperreactivity and mucus overproduction in asthmaNat Med20028888588912091879
- BarnesPJDrazenJMRennardSIThomsonNCAsthma and COPD: Basic Mechanisms and Clinical Management2nd edSan DiegoAcademic Press2009
- KondoHIchikawaYImokawaGPercutaneous sensitization with allergens through barrier-disrupted skin elicits a Th2-dominant cytokine responseEur J Immunol19982837697799541570
- LambrechtBNCarro-MuinoIVermaelenKPauwelsRAAllergen-induced changes in bone-marrow progenitor and airway dendritic cells in sensitized ratsAm J Respir Cell Mol Biol19992061165117410340935
- SugimotoTXiaoCIchikawaHNeonatal primary neuronal death induced by capsaicin and axotomy involves an apoptotic mechanismBrain Res19988071–21471549757021
- ZhengTOhMHOhSYSchroederJTGlickABZhuZTransgenic expression of interleukin-13 in the skin induces a pruritic dermatitis and skin remodelingJ Invest Dermatol2009129374275118830273
- KimNBaeKBKimMOOverexpression of cathepsin S induces chronic atopic dermatitis in miceJ Invest Dermatol201213241169117622170489
- PatelKRAvenLShaoFMast cell-derived neurotrophin 4 mediates allergen-induced airway hyperinnervation in early lifeMucosal Immunol2016961466147626860818