
Abstract
Allergic rhinitis (AR) is a chronic allergic disease of the upper respiratory system that affects approximately 10–40% of the global population. Due to the large number of plant pollen allergens with obvious seasonal variations, AR is common in China. AR is primarily caused by the abnormal regulation of the immune system. Its pathophysiological mechanism involves a series of immune cells and immune mediators, including cytokines. The present review summarizes the common allergens in China and the complex pathophysiological mechanism of AR. Additionally, host allergen contact, signal transduction, immune cell activation, cytokine release, and a series of inflammatory reactions are described according to their sequence of occurrence.
Introduction
Allergic rhinitis (AR), a chronic upper respiratory tract disease, is one of the most common allergic diseases resulting from exposure to specific allergens.Citation1,Citation2 The pathophysiology of AR involves IgE-mediated inflammation of the nasal mucosa.Citation3 In recent years, the incidence and prevalence of AR in China have increased, negatively impacting the economy and people’s quality of life.Citation4–6 AR, also known as hay fever, is characterized by a blocked nose, runny nose, itching, sneezing, and postnasal drip syndrome (PND), which involves eye itching, redness, and tearing.Citation3,Citation7 Dust mites, animal furs, and mold spores are the primary triggers for perennial allergic reactions,Citation8 whereas pollen and weed powder are the primary causes of seasonal allergies.Citation9 AR occurs when its corresponding allergen activates mast cells or basophils to release various immune mediators, including cytokines that mediate T helper 2 (Th2) cell maturation and mucosal inflammation.Citation10–12
The pathophysiological mechanism of AR is complex and involves the activation of effector cells and the release of immune mediators, such as leukotrienes (LTs), prostaglandins (PGD), and platelet-activating factors.Citation13 The nasal mucosa is the primary site of allergen exposure, and inflammatory reactions lead to AR symptom development.Citation14 The symptoms are predominantly triggered by the interactions between specific mast cells, IgE antibodies, and airborne allergens.Citation15 Allergen exposure induces inflammatory mediator release, causing the specific symptoms and the inflammatory morphology seen in AR.Citation15,Citation16
This review highlights the common allergens responsible for AR in China. As the pathogenesis of AR is caused by an individual’s immune response, this response and its underlying mechanisms are also comprehensively reviewed. It also summarizes the immune regulatory mechanisms that occur in host cells following allergen exposure.
AR Allergens in China
Regional Differences of Rhinitis Allergens in China
The prevalence of allergic diseases is closely related to various environmental allergens and direct contact, inhalation, or ingestion exposure levels.Citation17,Citation18 China is a large country with a vast geography and diverse climates and lifestyles, which leads to significant regional differences in AR distribution.Citation19 The most common pattern of sensitization in Southern and Eastern China is house dust mites sensitization, while the prevalence of low pollen sensitization in Western and Northern China is extremely high.Citation20 In China, over 38,000 adults have self-reported AR, and its prevalence varies widely across regions, ranging from 8.7% in Beijing in North China to 24.1% in Urumqi in Northwest China.Citation21 A cross-sectional survey was conducted in eight cities in China and revealed significant geographical differences in the prevalence of AR among Chinese children.Citation22 Therefore, identifying the specific allergens prevalent in particular regions that affect populations is important for the early diagnosis, treatment, and prevention of respiratory allergic diseases, including AR.Citation18 The incidence of allergic diseases has increased in China, particularly rhinitis caused by house dust mites and weed pollen (). Several environmental factors, such as cold air, fog, and tobacco smoke, play important roles in the etiology of AR and other respiratory allergic diseases.Citation23,Citation24 However, the sensitivity to these allergens differs among populations and different regions of China. In one study, the three most common self-reported allergens among individuals with AR in Northern China were Blattella germanica (16.6%), Dermatophagoides farinae (14.6%), and Dermatophagoides pteronyssinus (13.9%).Citation25 A multicenter study reported the most common allergens in Eastern China to be D. farinae (74.9%) and D. pteronyssinus (74.3%), while in the West, they were mugwort (60.0%) and marguerite (56.6%).Citation26 The most common allergens in South China are house dust mites (28.1%), cockroaches (24.3%), shrimp (19.2%), crabs (15.5%), and egg whites (9.9%).Citation27 An analysis of allergen-specific IgE in serum samples from individuals based in Guangzhou, Southern China, revealed that D. pteronyssinus (85.0%) had the highest proportion of specific IgE, followed by D. farinae (83.5%), egg whites (19.3%), milk (14.6%), German cockroaches (14.2%) and ox-eye daisy (12.6%).Citation28
Table 1 Partial Data for Rhinitis Patients in China
China is a vast territory that spans approximately 50 latitudes. Therefore, a great diversity in vegetation is seen across the country.Citation29 The main difference in allergens between different regions lies in the species and richness of the sensitized pollen present. Understanding the aerobiological data regarding the atmospheric pollen linked to micrometeorological indices can help determine the correlation between air pollen concentration and AR, thereby establishing the appropriate etiological diagnosis and treatment plan.Citation30,Citation31 In a study that was conducted over 2 years, the main air allergens in Guangxi were found to be Artemisia spores, providing basic information on seasonal allergies in Southern China and Southeast Asia.Citation32 During spring in Wuhan, airborne pollen primarily comes from Artemisia, willow, Acanthaceae, Lauraceae, and pine, while in autumn, it is primarily derived from Artemisia and Cucurbitaceae.Citation33 In Zhongshan City in Southern China, Gramineae is associated with the highest annual pollen levels, with other common pollen contributors being pine, alfalfa, water chestnut, and Casuarinaceae, which have peak seasons in February and March I.Citation34
Northern China is dry and windy, with abundant pollen in the air during the spring and autumn. Studies have examined the aerobiological data similarity matrices of eight selected pollen monitoring stations at 45–50°N for four different air allergens: birch (Betula platyphylla), grass (Gramineae), Artemisia (Artemisia), and ragweed (Ambrosia).Citation35 In Northern China, trees and weeds produce the main pollen types and are responsible for the two main allergic pollen seasons.Citation36 One season occurs in spring and is caused by tree pollen, including Fraxinus mandshurica, Betula platyphylla, and poplar trees. The other occurs in late summer and autumn and is caused by weed pollens such as Artemisia, amaranth, gourd, tall bran, and Chenopodium.Citation36–38
In Western Inner Mongolia, the typical peak pollen seasons occur in summer and autumn and involves primarily weeds and grass pollen, while in the east, there are two peak seasons in spring (involving mainly tree pollen) and summer and autumn (involving mainly weeds and grass pollen).Citation39 In Qingdao, China, children with AR are highly sensitive to ragweed and pollen.Citation18 Compared with South China, the main pollen allergens in North China are birch, Artemisia, poplar, and ragweed.
The prevalence of AR is higher in developed countries and currently affects 10–40% of adults and 2–25% of children worldwide. With the rapid economic development, the prevalence of AR has also increased in developing countries in recent decades.Citation40 Persistent AR prevalence is higher in Northern Europe and northern China compared to southern regions, while Western Europe has a higher prevalence of AR than Eastern Europe.Citation41 Different allergens exist in different regions, for example birch and grass pollen are the most common allergens in Denmark, while Japanese cedar pollen is the most common in Japan.Citation42
Since AR is a chronic disease caused by a specific allergen, it is important for patients to try to identify allergens and environmental factors that may trigger AR. Avoiding or minimizing exposure to disease-causing allergens should be the first management step for AR. Therefore, exploring the differences of allergens in China can help to identify and avoid allergens in specific regions and prevent AR.
Rhinitis Allergens
Dust mites, summer and autumn pollen, weed pollen, Neurospora crassa, and Blattella germanica are common allergens that cause AR in China ().Citation43 Dust mites mainly reside in mattresses, clothing, blankets, carpets, plush toys, and pets. Sensitization to house dust mites (HDM) is highest in South China, while the three most common seasonal airborne allergens in Northwest China are mugwort, ragweed, and dandelion pollen.Citation44 Neurospora crassa is a saprophyte that exists vegetatively as an incompletely septate syncytium.Citation45 Blattella germanica normally resides in human habitats, in warm, moist, and dark hidden places.Citation46 As B. germanica contains several symbiotic microorganisms,Citation47 insect excretions, and an exfoliated epidermis, it also carries allergens that may be transmitted to humans and result in AR.
Figure 1 Common allergens and pathogenesis of AR. (A). The main allergens causing AR in northern China (dust mites, summer and autumn pollen, weed pollen, Neurospora crassa, and Blattella germanica). (B). Immune response to allergens. Specific IgE binds to the high-affinity IgE Fc receptor on the surface of mast cells and induces the release of PAF, LTs, histamine, and other inflammatory mediators. Histamine binds its receptor, whereas IL-4, IL-5, TNF, and other cytokines stimulate eosinophils, and Neutrophils stimulate respiratory tract contraction, causing AR.
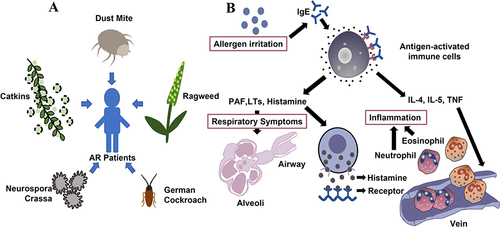
Sensitivity to common allergens among hospitalized patients in different geographical areas and who belong to different age groups varies considerably. Many factors, such as climate and plants, affect AR incidence.Citation48 Certain foods, such as mussels, shrimp, carp, eggs, and milk, increase AR incidence.Citation18 Some drugs, such as aspirin and nonsteroidal anti-inflammatory drugs, can also cause AR,Citation49 while physical factors, such as morning hours, winter, and cold air, are common AR triggers.Citation24
Direct contact with specific allergens is the first step in AR occurrence. This causes the immune system to be sensitized and activated, and immune response signals are subsequently transmitted between cells. Allergen-specific IgE binds to high-affinity IgE Fc receptors on the surface of mast cells, or the allergen directly binds to the sensitized target cell’s IgE,Citation50 thereby inducing the secretion of histamine, kininogenases, LTs, cytokines, and other mediators.Citation14 This causes the airway smooth muscles and telangiectasia to contract, increasing permeability and leading to excessive glandular secretion.Citation51 This process culminates in sneezing, runny nose, nasal itching, congestion, and other symptoms.Citation52 AR is an allergic disease caused by abnormal immune system regulation. As allergic reactions primarily involve immune cells, signaling molecules, and mediators, research on AR pathogenesis has focused on host immune responses to allergens. The following sections summarize the existing findings on the immune response to allergens.
Host Immune Responses to Allergens
Antigen Presentation and Sensitization
Antigen-presenting cells (APCs) are immune cells that uptake, process, and present antigens to the T and B lymphocytes.Citation57–59 Upon binding to antigens, water-soluble proteins are engulfed and processed by APCs (dendritic cells [DCs] and macrophages) and then processed by naïve CD4+ T lymphocytes (Th0) in the lymph nodes via major histocompatibility complex II molecules (MHCII) expressed on APC surfaces.Citation13,Citation60 The activation of CD4+ lymphocytes involves direct contact with MHCII molecules on APCs via specific T cell surface receptors and costimulatory receptors CD80 and CD86 on APCs, which connect to the CD28 receptors on T cells.Citation60 T cells from patients with AR are primarily transformed into allergen-specific Th2 cells.Citation61 These cells release cytokines, including interleukin (IL)-4, IL-5, and IL-13, to initiate the inflammatory immune response.Citation13 IL-4 stimulates the differentiation of specific B cells into antibody-producing plasma cells. IL-4 and IL-13 send the first signal from T cells.Citation62 IL-4, IL-5, and IL-13 interact with B cell surface receptors, inducing ε germline transcription to produce immunoglobulins.Citation63 The second signal is produced by the costimulatory interaction between CD154 (CD40 ligand) on the surface of activated T cells and CD40 molecules on the surface of B cells; this interaction activates B cells and stimulates class-switch recombination, inducing IgE production.Citation63,Citation64 This, in turn, stimulates cytokine secretion and triggers the inflammatory response.
Although IgE antibodies only account for a small proportion of the entire antibody family,Citation13 they induce strong inflammation by binding to specific cell surface receptors and crosslinking with antigens. Allergen-specific IgE antibodies bind strongly to high-affinity receptors (FcεRI) expressed on mast cells and basophils.Citation65 Upon subsequent exposure, IgE antibodies bound to FcεRI receptors recognize specific allergens,Citation66,Citation67 resulting in the rapid degranulation of mast cells and basic granulocyte vesicles and the release of histamine, platelet-activating factor, and tryptase.Citation13,Citation68 Activated mast cells release membrane-stored arachidonic acid and participate in the production of cysteinyl LTs (LTC4, LTD4, and LTE4) and prostaglandins (mainly PGD2).Citation69 These reactions induce the contraction of the smooth muscle in the respiratory tract and vasodilation and enhance permeability and excessive glandular secretion ().
Prophase Immune Response
Nasal Mucosal Receptor Signal Transduction
The surface of the nasal mucosa is covered with receptors and is innervated by a group of sensory nerve fibers, including A, δ, and unmyelinated C fibers, and sympathetic and parasympathetic nerves.Citation14,Citation70 When the temperature difference is high during the turn of seasons in China, catkins and ragweed pollen allergens are blown into the air. For patients with rhinitis that occurs following exposure to specific allergens, direct contact with the respective antigens activates the immune system and increases its sensitivity. In this process, the specific IgE binds to high-affinity IgE Fc receptors on mast cell surfaces.Citation50,Citation71 The FcεRI receptor is a member of the polysubunit immunoreactive receptor (multisubunit immune response receptor [MIRR]) family.Citation72 FcεRI is mainly involved in type I hypersensitivity and mediates mast cell degranulation and the release of inflammatory mediators. FcεRI aggregation and crosslinking activate the Lyn PTK and src families; activated Lyn phosphorylates the β-chain, γ-chain, and adjacent tyrosine proteins. The latter process, in turn, activates Lyn and Syk.Citation72 The activated (ie, depolarized) sensory nerves transmit signals to the central nervous system, resulting in motor reflexes, such as sneezing, watery nose, nasal itching, and nasal congestion.
Histamine Release
Histamine is a biogenic amine secreted by mast cells during allergic reactions.Citation73 It has strong and diverse biological functions, including in AR pathophysiology.Citation74 Lung tissue contains four histamine receptors: H1, H2, H3, and H4.Citation75–78 The pulmonary artery has a biphasic response to histamine due to H1 receptor-mediated vasoconstriction and H2 receptor-mediated vasodilation.Citation79 Histamine causes plasma leakage from the microcapillary venules by affecting the bronchial microcirculatory system. Additionally, histamine accelerates chloride ion transport in the respiratory epithelial cells, which is closely related to the water transfer process within the respiratory tract.Citation80 When an inhaled allergen comes into contact with the bronchi of patients with northern rhinitis, it triggers the release of histamine to the surface of the respiratory tract, which can then be identified in the bronchoalveolar lavage fluid (BALF).Citation79 The mast cells release other inflammatory mediators, such as 9 α, 11 β-prostaglandins F2-α and trypsin.
The early allergen response can be attributed to the allergen-induced degranulation of nasal mast cells. This phenomenon causes IgE-mediated histamine release into the nasal mucosaCitation81 and upregulates proinflammatory cytokine expression, mediating the early immune response,Citation70 including allergic reactions and inflammation.Citation82–85 The release of histamine from mast cells promotes H1 receptor activation.Citation86,Citation87 The activated sensory nerves transmit the signals to the central nervous system, which causes motor reflexes, such as itching and sneezing.Citation88,Citation89 Histamine stimulates watery secretions in the mucosa, leading to rhinorrhea and the expression of H1 and H2 receptors in the nasal vessels, increasing vascular permeability and vasodilation, which causes nasal mucosal congestion.Citation90–92
Sensory nerve stimulation induces the release of acetylcholine from the parasympathetic nerves. Cytokines are increased on both sides of the central reflex, which upregulates nasal mucus secretion. H1R binds to the G protein GQ/11, activating phospholipase C and stimulating the production of 5-inositol triphosphate; this induces calcium and diacylglycerol mobilization and activates protein kinase C (PKC).Citation93,Citation94 PKC activates the transcription factor NF-kappa B, which enhances the transcription of proinflammatory genes, while calcium promotes nasal vasodilation. These processes augment the pericellular permeability and the antigen capture and processing ability of APCs.Citation95,Citation96 The histamine H2 receptor causes nasal congestion by binding to the GS protein. Hypo 2R activates adenylyl cyclase (AC) by binding to the GS protein. In turn, AC stimulates cyclic adenosine monophosphate (cAMP) and induces the secretion of the cAMP response element-binding protein.Citation97–99 CAMP-dependent protein kinase signaling inhibits LT synthesis in neutrophils.Citation100 The histamine H3 receptor inhibits the release of substance P from nasal nociceptive sensory nervesCitation101–103 and promotes the IgE-mediated activation of mast cell degranulation.Citation104 Numerous antihistamine drugs, such as loratadine, and methods are currently available to treat AR. Effective AR prevention and treatment strongly relies on the regulation of histamine release.
Late Immune Response
Primary Effector Cells Release Cytokines and Chemokines
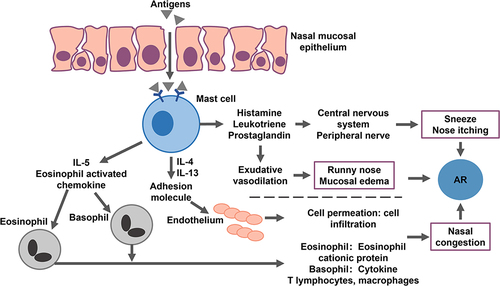
Mast cells are the primary effector cells in the early allergen response. Mast cells release cytokines and attract eosinophils, Th2 cells, and neutrophils to the nasal mucosa.Citation63 The late response in AR involves the release of newly secreted cytokines, such as LTs, and the recruitment of numerous effector cells from the lymphoid tissue and general circulation.Citation57 These responses sustain inflammation and prolong allergic reactions.Citation57,Citation105 Although nasal congestion and sneezing can last for a significant period, persistent nasal congestion indicates late-stage allergic reactions.Citation16
Mast cells release cytokines, such as IL-4, IL-13, and tumor necrosis factor-α. In turn, the cytokines activate the endothelial cells and upregulate the expression of adhesion molecules, such as intercellular adhesion molecule-1 and vascular cell adhesion molecule-1, inducing the migration of eosinophils, T cells, basophils, and neutrophils.Citation105–107 IL-5 participates in the early activation, terminal differentiation, growth, and survival of eosinophils and inflammation. In AR, IL-5 signal transduction involves JAK-STAT-p38MAPK-NFkB activation, extracellular matrix remodeling, epithelial-mesenchymal transition, and immune response mediation. Allergens increase the expression of IL-5 receptor α on the eosinophil surfaces, which leads to increased IL-5 levels.Citation108 It transmits signals via the IL-5 receptor α-chain and β-chain complexes.Citation109 IL-18 participates in the immune response and plays a vital role in the inflammatory pathway, being involved in the production, transformation, and maturation of pathogenic eosinophils (CD101+ and CD274+). In a previous study, mice models showed that IL-18 promotes ovalbumin immunity and the migration of eosinophils to the respiratory tract following ragweed pollen exposure.Citation110 Additionally, IL-18 stimulates human invariant natural killer T cells and endothelial cells in vitro and induces the eosinophil-active cytokines IL-5 and IL-13.Citation110 The mediators released by mast cells, such as LTs, prostaglandins, and platelet-activating factors, lead to the observed reactions and immune chemotaxis,Citation13 which further helps maintain inflammation. After the release of histamine, various cytokines and chemokines released by different cell types interact with each other and mediate AR pathogenesis.
Inflammatory Reaction
Eosinophils and neutrophils migrate to the respiratory tract of patients with AR, maintaining the inflammatory state and prolonging its related reactions by releasing corresponding cytokines and inflammatory mediators.Citation111–113 A recent nasal cytological study on 468 patients with AR indicated that neutrophils mediated major inflammatory cell responses in 14.32% of patients, whereas a mixture of eosinophils and neutrophils was observed in 23.93% of patients.Citation114 Late mucosal inflammation in patients with HDM-specific AR activates neutrophil circulation and increases neutrophil migration into the nasal mucosa.Citation112 T cells play a major role in allergic respiratory inflammation. Th1 cell differentiation from CD_4 T cells (TH0) is mediated by IL-12 and interferon-γ. The number of Th1 cells and the level of Th1-derived interferon-γ are relatively lower in the nasal lavage fluid of patients with AR than in individuals without AR.Citation115 Activated T cells promote allergic inflammation by releasing IL-5. Eosinophils are granulated inflammatory cells involved in the innate immune system which possess multiple inflammatory functions.Citation116 In allergic inflammation, these cells promote Th2 polarization by releasing IL-4, IL-25, and indole-2,3 dioxygenase. Eosinophils promote B cell proliferation and antibody induction and stimulate the secretion of several chemokines, including CCL17/CCL22, which in turn recruits Th2 cells to interact with DCs.Citation117,Citation118 In pollen-sensitized AR patients, nasal mucosal eosinophils increase within a few days of an allergen challenge.Citation119 Immune cells mediate allergic inflammatory reactions in the respiratory tract by inducing the release of the corresponding cytokines.
Inflammation persists in patients with rhinitis, during which activated eosinophils flow into the nasal mucosa. Nasal epithelium injury exposes the nerve fibers, and overactivity in the nasal cavity triggers AR reactions. Epithelial damage is caused by the toxic effect of superoxide anions, the production of hydrogen peroxide, and the release of granule products, such as eosinophil cationic proteins, eosinophil-derived neurotoxins, and the basic protein released by eosinophils.Citation120 Eosinophils secrete IL-5, which promotes the activation and survival of eosinophils in an autocrine manner.Citation121 Additionally, T cells and mast cells promote the survival of eosinophils in the nasal mucosa by releasing granulocyte-macrophage colony-stimulating factor and IL-5.Citation122,Citation123 These proinflammatory molecules are chemoattractants that enhance the Th2 response and participate in the recruitment of eosinophils, basophils, and T cells to the nasal mucosa (). Cytokines and other mediators induce immune cells to continuously increase and maintain the number of proinflammatory molecules in the nasal mucosa, thereby sustaining the inflammatory response and mediating AR pathogenesis.
Figure 2 Allergens trigger the body’s AR immune response process.
Understanding the mechanisms that trigger AR is crucial for the treatment of AR. At present, the identification of novel biomarkers of AR phenotype or endotype patients is an important research direction, which can be used to predict the therapeutic effect and develop treatment strategies. Further elucidating the novel molecular mechanisms involved in the allergen specific reaction in the nasal mucosa, as well as the transformation and integration of biochemical methods such as genomics in the treatment of AR are also relatively frontier research areas.
Conclusions
AR is one of the common allergic diseases associated with the immune response and is mediated by IgE. AR is closely related to host immunity. Dominant allergens vary across the different vegetation environments in China. Compared with traditional allergens, AR caused by specific allergens places a higher burden on the immune system. Upon exposure, the allergens are engulfed, processed, and presented by APCs to the T cells. The T cells then differentiate and proliferate into Th2 cells, which then secrete cytokines and induce IgE on B cells and stimulate B cell differentiation into specific antibody-producing plasma cells. The IgE secreted by the plasma cells binds to the receptor on mast cell surfaces. Degranulated mast cells secrete more chemical mediators, including histamine, which induce AR. Understanding the China-specific allergens of AR and the host regulatory mechanisms that cause AR are important for the development of therapeutic methods for AR.
Data Sharing Statement
The data presented in this study are available in the article.
Author Contributions
Qirong Li, Xinyi Zhang and Dongxu Wang wrote the manuscript. Qirong Li, Xinyi Zhang, Qiang Feng, Hengzong Zhou, Chaoyang Ma, Chao Lin, Dongxu Wang, Jianmei Yin searched PubMed and Web of Science for citations and prepared figures. All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
Acknowledgments
Appreciate all participants for their contributions.
Additional information
Funding
References
- Tohidinik HR, Mallah N, Takkouche B. History of allergic rhinitis and risk of asthma; a systematic review and meta-analysis. World Allergy Organ J. 2019;12(10):100069. doi:10.1016/j.waojou.2019.100069
- Bousquet J, Khaltaev N, Cruz AA, et al. Allergic Rhinitis and its Impact on Asthma (ARIA) 2008 update (in collaboration with the World Health Organization, GA(2)LEN and AllerGen). Allergy. 2008;63(Suppl 86):8–160. doi:10.1111/j.1398-9995.2007.01620.x
- Rahim NA, Jantan I, Said MM, Jalil J, Abd Razak AF, Husain K. Anti-allergic rhinitis effects of medicinal plants and their bioactive metabolites via suppression of the immune system: a mechanistic review. Front Pharmacol. 2021;12:660083. doi:10.3389/fphar.2021.660083
- Wang XD, Zheng M, Lou HF, et al. An increased prevalence of self-reported allergic rhinitis in major Chinese cities from 2005 to 2011. Allergy. 2016;71(8):1170–1180. doi:10.1111/all.12874
- Meltzer EO. Allergic rhinitis: burden of illness, quality of life, comorbidities, and control. Immunol Allergy Clin North Am. 2016;36(2):235–248. doi:10.1016/j.iac.2015.12.002
- Leynaert B, Neukirch C, Liard R, Bousquet J, Neukirch F. Quality of life in allergic rhinitis and asthma. A population-based study of young adults. Am J Respir Crit Care Med. 2000;162(4 Pt 1):1391–1396. doi:10.1164/ajrccm.162.4.9912033
- Skoner DP. Allergic rhinitis: definition, epidemiology, pathophysiology, detection, and diagnosis. J Allergy Clin Immunol. 2001;108(1 Suppl):S2–S8. doi:10.1067/mai.2001.115569
- Pomes A, Chapman MD, Wunschmann S. Indoor allergens and allergic respiratory disease. Curr Allergy Asthma Rep. 2016;16(6):43. doi:10.1007/s11882-016-0622-9
- Wakamiya S, Matsune S, Okubo K, Aramaki E. Causal relationships among pollen counts, tweet numbers, and patient numbers for seasonal allergic rhinitis surveillance: retrospective analysis. J Med Internet Res. 2019;21(2):10450.
- Wheatley LM, Togias A, Solomon CG. Clinical practice. Allergic rhinitis. N Engl J Med. 2015;372(5):456–463. doi:10.1056/NEJMcp1412282
- Barnes PJ. Pathophysiology of allergic inflammation. Immunol Rev. 2011;242(1):31–50. doi:10.1111/j.1600-065X.2011.01020.x
- Georas SN, Guo J, De Fanis U, Casolaro V. T-helper cell type-2 regulation in allergic disease. Eur Respir J. 2005;26(6):1119–1137. doi:10.1183/09031936.05.00006005
- Bernstein DI, Schwartz G, Bernstein JA. Allergic Rhinitis: mechanisms and Treatment. Immunol Allergy Clin North Am. 2016;36(2):261–278. doi:10.1016/j.iac.2015.12.004
- Watts AM, Cripps AW, West NP, Cox AJ. Modulation of allergic inflammation in the nasal mucosa of allergic rhinitis sufferers with topical pharmaceutical agents. Front Pharmacol. 2019;10:294. doi:10.3389/fphar.2019.00294
- Galli SJ, Tsai M. IgE and mast cells in allergic disease. Nat Med. 2012;18(5):693–704. doi:10.1038/nm.2755
- Bousquet J, Van Cauwenberge P, Khaltaev N. Allergic rhinitis and its impact on asthma. J Allergy Clin Immunol. 2001;108(5 Suppl):S147–S334. doi:10.1067/mai.2001.118891
- Sun BQ, Chen DH, Zheng PY, et al. Allergy-related evidences in relation to serum IgE: data from the China state key laboratory of respiratory disease, 2008–2013. Biomed Environ Sci. 2014;27(7):495–505. doi:10.3967/bes2014.081
- Lin H, Lin R, Li N. Sensitization rates for various allergens in children with allergic rhinitis in Qingdao, China. Int J Environ Res Public Health. 2015;12(9):10984–10994. doi:10.3390/ijerph120910984
- Luo WT, Wang DD, Zhang T, et al. Prevalence patterns of allergen sensitization by region, gender, age, and season among patients with allergic symptoms in Mainland China: a Four-year Multicenter Study. Allergy. 2021;76(2):589–593. doi:10.1111/all.14597
- Hou XQ, Luo WT, Wu LT, et al. Associations of Four sensitization patterns revealed by Latent Class Analysis with Clinical symptoms: a multi-center study of China. Eclinicalmedicine. 2022;46. doi:10.1016/j.eclinm.2022.101349
- Zhang L, Han D, Huang D, et al. Prevalence of self-reported allergic rhinitis in eleven major cities in China. Int Arch Allergy Immunol. 2009;149(1):47–57. doi:10.1159/000176306
- Li F, Zhou Y, Li S, et al. Prevalence and risk factors of childhood allergic diseases in eight metropolitan cities in China: a multicenter study. BMC Public Health. 2011;11:437. doi:10.1186/1471-2458-11-437
- Feleszko W, Ruszczyński M, Jaworska J, Strzelak A, Zalewski BM, Kulus M. Environmental tobacco smoke exposure and risk of allergic sensitisation in children: a systematic review and meta-analysis. Arch Dis Child. 2014;99(11):985–992. doi:10.1136/archdischild-2013-305444
- Li CW, Chen DD, Zhong JT, et al. Epidemiological characterization and risk factors of allergic rhinitis in the general population in Guangzhou City in China. PLoS One. 2014;9(12):e114950. doi:10.1371/journal.pone.0114950
- Huang Y, Zhang Y, Zhang L. Prevalence of allergic and nonallergic rhinitis in a rural area of northern China based on sensitization to specific aeroallergens. Allergy Asthma Clin Immunol. 2018;14:77. doi:10.1186/s13223-018-0299-9
- Wu L, Luo W, Hu H, et al. A multicenter study assessing risk factors and aeroallergens sensitization characteristics in children with self-reported allergic rhinitis in China. J Asthma Allergy. 2021;14:1453–1462. doi:10.2147/jaa.S342495
- Luo WT, Hu HS, Tang WB, et al. Allergen sensitization pattern of allergic adults and children in southern China: a survey based on real life data. Allergy Asthma Cl Im. 2019;15. doi:10.1186/s13223-019-0357-y
- Liao CX, Huang DM, Hu HS, et al. 广东地区儿童呼吸道过敏性疾病患者过敏原特异性IgE抗体检测的研究 [Analysis of serum allergen-specific IgE in children with respiratory allergic disease in Guangdong]. Zhonghua Yu Fang Yi Xue Za Zhi. 2021;55(5):613–619. Chinese. doi:10.3760/cma.j.cn112150-20201217-01467
- Cheng YY, Ruan B. Species and distribution of airborne pollen plants in major cities of China. Chin J Allergy Clin Immunol. 2015;2015:136–141.
- Pfaar O, Karatzas K, Bastl K, et al. Pollen season is reflected on symptom load for grass and birch pollen-induced allergic rhinitis in different geographic areas-an EAACI Task Force Report. Allergy. 2020;75(5):1099–1106. doi:10.1111/all.14111
- Geller-Bernstein C, Portnoy JM. The clinical utility of pollen counts. Clin Rev Allergy Immunol. 2019;57(3):340–349. doi:10.1007/s12016-018-8698-8
- Chen K, Liao YF, Zhang JT. The major aeroallergens in Guangxi, China. Clin Allergy. 1988;18(6):589–596. doi:10.1111/j.1365-2222.1988.tb02910.x
- Wang Y, Gao Z, Lv H, Xu Y. Online public attention toward allergic rhinitis in Wuhan, China: infodemiology study using Baidu index and meteorological data. Front Public Health. 2022;10:971525. doi:10.3389/fpubh.2022.971525
- Wang Y, Zhan Z. Airborne pollens in Zhongshan city. Chin J Allergy Clin Immunol. 2012;6:261–265.
- Bastl K, Bastl M, Bergmann KC, Berger U. How to do a clinical trial? Recommendations from the aerobiological point of view. World Allergy Organ J. 2019;12(4):100020. doi:10.1016/j.waojou.2019.100020
- Xu Y, Guan K, Sha L, et al. Sensitization Profiles of Timothy Grass Pollen in Northern China. J Asthma Allergy. 2021;14:1431–1439. doi:10.2147/jaa.S334183
- Ouyang YH, Zhang DS, Fan EZ, Li Y, Zhang L. 夏秋季花粉症症状与花粉播散的相关性分析 [Correlation between symptoms of pollen allergic rhinitis and pollen grain spreading in summer and autumn]. Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2012;47(8):623–627. Chinese.
- Hu WN, Zhu L, Xie LF, et al. 北京市三年内每日花粉浓度与变应性鼻炎患者就诊比例的关联分析 [Correlation between the visiting rate of patients with allergic rhinitis and airborne pollen concentrations in Beijing in recent 3 years]. Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2017;52(1):31–36. Chinese. doi:10.3760/cma.j.issn.1673-0860.2017.01.006
- Wang XY, Ma TT, Wang XY, et al. Prevalence of pollen-induced allergic rhinitis with high pollen exposure in grasslands of northern China. Allergy. 2018;73(6):1232–1243. doi:10.1111/all.13388
- Zhang Y, Zhang L. Increasing prevalence of allergic rhinitis in China. Allergy Asthma Immun. 2019;11(2):156–169. doi:10.4168/aair.2019.11.2.156
- Wise SK, Damask C, Roland LT, et al. International consensus statement on allergy and rhinology: allergic rhinitis-2023. Int Forum Allergy Rh. 2023;13(4):293–859. doi:10.1002/alr.23090
- Meng YF, Wang CS, Zhang L. Advances and novel developments in allergic rhinitis. Allergy. 2020;75(12):3069–3076. doi:10.1111/all.14586
- Yang Y, Zhao Y, Wang CS, Wang XD, Zhang L. 变应性鼻炎患者10030例吸入性变应原谱分析 [Prevalence of sensitization to aeroallergens in 10 030 patients with allergic rhinitis]. Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2011;46(11):914–920. Chinese.
- Lou H, Ma S, Zhao Y, et al. Sensitization patterns and minimum screening panels for aeroallergens in self-reported allergic rhinitis in China. Sci Rep. 2017;7(1):9286. doi:10.1038/s41598-017-10111-9
- Dunlap JC, Borkovich KA, Henn MR, et al. Enabling a community to dissect an organism: overview of the Neurospora functional genomics project. Adv Genet. 2007;57:49–96. doi:10.1016/s0065-2660(06)57002-6
- Nazari M, Motlagh BA, Nasirian H. Toxicity of cypermethrin and chlorpyrifos against German cockroach [Blattella germanica (Blattaria: blattellidae)] strains from Hamadan, Iran. Pak J Biol Sci. 2016;19(6):259–264. doi:10.3923/pjbs.2016.259.264
- Pan X, Wang X, Zhang F. New insights into cockroach control: using functional diversity of blattella germanica symbionts. Insects. 2020;11(10):696.
- Heinzerling LM, Burbach GJ, Edenharter G, et al. GA(2)LEN skin test study I: GA(2)LEN harmonization of skin prick testing: novel sensitization patterns for inhalant allergens in Europe. Allergy. 2009;64(10):1498–1506. doi:10.1111/j.1398-9995.2009.02093.x
- Liva GA, Karatzanis AD, Prokopakis EP. Review of rhinitis: classification, types, pathophysiology. J Clin Med. 2021;10(14):3183.
- Gould HJ, Sutton BJ. IgE in allergy and asthma today. Nat Rev Immunol. 2008;8(3):205–217. doi:10.1038/nri2273
- Togias A. Rhinitis and asthma: evidence for respiratory system integration. J Allergy Clin Immunol. 2003;111(6):1171–83; quiz 1184. doi:10.1067/mai.2003.1592
- Rondón C, Fernández J, López S, et al. Nasal inflammatory mediators and specific IgE production after nasal challenge with grass pollen in local allergic rhinitis. J Allergy Clin Immunol. 2009;124(5):1005–11.e1. doi:10.1016/j.jaci.2009.07.018
- Wang X, Guo M, Wang H, Wang X. Pollen allergen sensitization feature of seasonal allergic rhinitis in children and adolescents in northern China. J Clin Otorhinolaryngol Head Neck Surg. 2020;34(11):2096–7993. doi:10.13201/j.issn
- Cheng L, Chen J, Fu Q, et al. Chinese Society of Allergy Guidelines for diagnosis and treatment of allergic rhinitis. Allergy Asthma Immunol Res. 2018;10(4):300–353. doi:10.4168/aair.2018.10.4.300
- Yang L, Zhang Q, Zhang P. Analysis of HLA-DRB1 allele polymorphism for patients with allergic rhinitis. Chin J Otorhinolaryngol. 1999;34(3):0412–3948.
- Li L, Guan K. Quality of life in 164 allergic rhinitis patients caused by different aeroallergens. J Clin Otorhinolaryngol Head Neck Surg. 2015;29(3):1101–1781. doi:10.13201/j.issn
- Mandhane SN, Shah JH, Thennati R. Allergic rhinitis: an update on disease, present treatments and future prospects. Int Immunopharmacol. 2011;11(11):1646–1662. doi:10.1016/j.intimp.2011.07.005
- Takhar P, Smurthwaite L, Coker HA, et al. Allergen drives class switching to IgE in the nasal mucosa in allergic rhinitis. J Immunol. 2005;174(8):5024–5032. doi:10.4049/jimmunol.174.8.5024
- Lambrecht BN. Allergen uptake and presentation by dendritic cells. Curr Opin Allergy Clin Immunol. 2001;1(1):51–59. doi:10.1097/01.all.0000010985.57414.74
- KleinJan A, Willart M, van Rijt LS, et al. An essential role for dendritic cells in human and experimental allergic rhinitis. J Allergy Clin Immunol. 2006;118(5):1117–1125. doi:10.1016/j.jaci.2006.05.030
- Van Overtvelt L, Wambre E, Maillere B, et al. Assessment of Bet v 1-specific CD4+ T cell responses in allergic and nonallergic individuals using MHC class II peptide tetramers. J Immunol. 2008;180(7):4514–4522. doi:10.4049/jimmunol.180.7.4514
- Stone KD, Prussin C, Metcalfe DD. IgE, mast cells, basophils, and eosinophils. J Allergy Clin Immunol. 2010;125(2 Suppl 2):S73–S80. doi:10.1016/j.jaci.2009.11.017
- Sin B, Togias A. Pathophysiology of allergic and nonallergic rhinitis. Proc Am Thorac Soc. 2011;8(1):106–114. doi:10.1513/pats.201008-057RN
- Janeway CATP, Walport M, Shlomchik M. The Immune System in Health and Disease Immunobiology. NY: Gerald Publishing. 2001:41.
- Kraft S, Kinet JP. New developments in FcepsilonRI regulation, function and inhibition. Nat Rev Immunol. 2007;7(5):365–378. doi:10.1038/nri2072
- Fewtrell C, Metzger H. Larger oligomers of IgE are more effective than dimers in stimulating rat basophilic leukemia cells. J Immunol. 1980;125(2):701–710.
- Knol EF. Requirements for effective IgE cross-linking on mast cells and basophils. Mol Nutr Food Res. 2006;50(7):620–624. doi:10.1002/mnfr.200500272
- Norman PS, Naclerio RM, Creticos PS, Togias A, Lichtenstein LM. Mediator release after allergic and physical nasal challenges. Int Arch Allergy Appl Immunol. 1985;77(1–2):57–63. doi:10.1159/000233753
- Peters-Golden M, Gleason MM, Togias A. Cysteinyl leukotrienes: multi-functional mediators in allergic rhinitis. Clin Exp Allergy. 2006;36(6):689–703. doi:10.1111/j.1365-2222.2006.02498.x
- Wang D, Smitz J, Waterschoot S, Clement P. An approach to the understanding of the nasal early-phase reaction induced by nasal allergen challenge. Allergy. 1997;52(2):162–167. doi:10.1111/j.1398-9995.1997.tb00970.x
- Kalesnikoff J, Galli SJ. Anaphylaxis: mechanisms of mast cell activation. Chem Immunol Allergy. 2010;95:45–66. doi:10.1159/000315937
- Johnson SA, Pleiman CM, Pao L, Schneringer J, Hippen K, Cambier JC. Phosphorylated immunoreceptor signaling motifs (ITAMs) exhibit unique abilities to bind and activate Lyn and Syk tyrosine kinases. J Immunol. 1995;155(10):4596–4603.
- Dale HH, Laidlaw PP. Histamine shock. J Physiol. 1919;52(5):355–390. doi:10.1113/jphysiol.1919.sp001837
- White MV. The role of histamine in allergic diseases. J Allergy Clin Immunol. 1990;86(4 Pt 2):599–605. doi:10.1016/s0091-6749(05)80223-4
- Jones JV. The nature of the pulmonary receptors excited by antihistamines. Br J Pharmacol Chemother. 1952;7(3):450–454. doi:10.1111/j.1476-5381.1952.tb00712.x
- Tucker A, Weir EK, Reeves JT, Grover RF. Histamine H1- and H2-receptors in pulmonary and systemic vasculature of the dog. Am J Physiol. 1975;229(4):1008–1013. doi:10.1152/ajplegacy.1975.229.4.1008
- Ichinose M, Barnes PJ. Inhibitory histamine H3-receptors on cholinergic nerves in human airways. Eur J Pharmacol. 1989;163(2–3):383–386. doi:10.1016/0014-2999(89)90212-4
- Kay LJ, Suvarna SK, Peachell PT. Histamine H4 receptor mediates chemotaxis of human lung mast cells. Eur J Pharmacol. 2018;837:38–44. doi:10.1016/j.ejphar.2018.08.028
- Ahmed T, Mirbahar KB, Oliver W, Eyre P, Wanner A. Characterization of H1- and H2-receptor function in pulmonary and systemic circulations of sheep. J Appl Physiol Respir Environ Exerc Physiol. 1982;53(1):175–184. doi:10.1152/jappl.1982.53.1.175
- Nadel JA, Davis B, Phipps RJ. Control of mucus secretion and ion transport in airways. Annu Rev Physiol. 1979;41:369–381. doi:10.1146/annurev.ph.41.030179.002101
- Taylor-Clark T. Histamine in allergic rhinitis. Adv Exp Med Biol. 2010;709:33–41. doi:10.1007/978-1-4419-8056-4_4
- Huang H, Li Y, Liang J, Finkelman FD. Molecular regulation of histamine synthesis. Front Immunol. 2018;9:1392. doi:10.3389/fimmu.2018.01392
- Schwartz JC, Arrang JM, Garbarg M, Pollard H, Ruat M. Histaminergic transmission in the mammalian brain. Physiol Rev. 1991;71(1):1–51. doi:10.1152/physrev.1991.71.1.1
- Hill SJ. Distribution, properties, and functional characteristics of three classes of histamine receptor. Pharmacol Rev. 1990;42(1):45–83.
- Hill SJ, Ganellin CR, Timmerman H, et al. International union of pharmacology. XIII. Classification of histamine receptors. Pharmacol Rev. 1997;49(3):253–278.
- Doyle WJ, Boehm S, Skoner DP. Physiologic responses to intranasal dose-response challenges with histamine, methacholine, bradykinin, and prostaglandin in adult volunteers with and without nasal allergy. J Allergy Clin Immunol. 1990;86(6 Pt 1):924–935. doi:10.1016/s0091-6749(05)80156-3
- Bachert C. The role of histamine in allergic disease: re-appraisal of its inflammatory potential. Allergy. 2002;57(4):287–296. doi:10.1034/j.1398-9995.2002.1r3542.x
- Schmelz M, Schmidt R, Bickel A, Handwerker HO, Torebjork HE. Specific C-receptors for itch in human skin. J Neurosci. 1997;17(20):8003–8008.
- Andrew D, Craig AD. Spinothalamic lamina I neurons selectively sensitive to histamine: a central neural pathway for itch. Nat Neurosci. 2001;4(1):72–77. doi:10.1038/82924
- Secher C, Kirkegaard J, Borum P, Maansson A, Osterhammel P, Mygind N. Significance of H1 and H2 receptors in the human nose: rationale for topical use of combined antihistamine preparations. J Allergy Clin Immunol. 1982;70(3):211–218. doi:10.1016/0091-6749(82)90044-6
- Wood-Baker R, Lau L, Howarth PH. Histamine and the nasal vasculature: the influence of H1 and H2-histamine receptor antagonism. Clin Otolaryngol Allied Sci. 1996;21(4):348–352. doi:10.1111/j.1365-2273.1996.tb01085.x
- Togias A. H1-receptors: localization and role in airway physiology and in immune functions. J Allergy Clin Immunol. 2003;112(4 Suppl):S60–S68. doi:10.1016/s0091-6749(03)01878-5
- De Backer MD, Gommeren W, Moereels H, et al. Genomic cloning, heterologous expression and pharmacological characterization of a human histamine H1 receptor. Biochem Biophys Res Commun. 1993;197(3):1601–1608. doi:10.1006/bbrc.1993.2662
- Umemori H, Inoue T, Kume S, et al. Activation of the G protein Gq/11 through tyrosine phosphorylation of the alpha subunit. Science. 1997;276(5320):1878–1881. doi:10.1126/science.276.5320.1878
- Flynn AN, Itani OA, Moninger TO, Welsh MJ. Acute regulation of tight junction ion selectivity in human airway epithelia. Proc Natl Acad Sci U S A. 2009;106(9):3591–3596. doi:10.1073/pnas.0813393106
- Georas SN, Rezaee F. Epithelial barrier function: at the front line of asthma immunology and allergic airway inflammation. J Allergy Clin Immunol. 2014;134(3):509–520. doi:10.1016/j.jaci.2014.05.049
- Houston C, Wenzel-Seifert K, Burckstummer T, Seifert R. The human histamine H2-receptor couples more efficiently to Sf9 insect cell Gs-proteins than to insect cell Gq-proteins: limitations of Sf9 cells for the analysis of receptor/Gq-protein coupling. J Neurochem. 2002;80(4):678–696. doi:10.1046/j.0022-3042.2001.00746.x
- Hegyi K, Falus A, Toth S. Elevated CREB activity in embryonic fibroblasts of gene-targeted histamine deficient mice. Inflamm Res. 2007;56(8):339–344. doi:10.1007/s00011-007-7049-7
- Shayo C, Davio C, Brodsky A, et al. Histamine modulates the expression of c-fos through cyclic AMP production via the H2 receptor in the human promonocytic cell line U937. Mol Pharmacol. 1997;51(6):983–990. doi:10.1124/mol.51.6.983
- Flamand N, Plante H, Picard S, Laviolette M, Borgeat P. Histamine-induced inhibition of leukotriene biosynthesis in human neutrophils: involvement of the H2 receptor and cAMP. Br J Pharmacol. 2004;141(4):552–561. doi:10.1038/sj.bjp.0705654
- Bongers G, Bakker RA, Leurs R. Molecular aspects of the histamine H3 receptor. Biochem Pharmacol. 2007;73(8):1195–1204. doi:10.1016/j.bcp.2007.01.008
- Nieto-Alamilla G, Marquez-Gomez R, Garcia-Galvez A-M, Morales-Figueroa G-E, Arias-Montano J-A. The histamine H3 receptor: structure, pharmacology, and function. Mol Pharmacol. 2016;90(5):649–673. doi:10.1124/mol.116.104752
- Wijtmans M, Leurs R, de Esch I. Histamine H 3 receptor ligands break ground in a remarkable plethora of therapeutic areas. Expert Opin Investig Drugs. 2007;16(7):967–985. doi:10.1517/13543784.16.7.967
- Xu H, Gu L-N, Yang Q-Y, Zhao D-Y, Liu F. MiR-221 promotes IgE-mediated activation of mast cells degranulation by PI3K/Akt/PLCγ/Ca2+ pathway. J Bioenerg Biomembr. 2016;48(3):293–299. doi:10.1007/s10863-016-9659-7
- Pawankar R, Mori S, Ozu C, Kimura S. Overview on the pathomechanisms of allergic rhinitis. Asia Pac Allergy. 2011;1(3):157–167. doi:10.5415/apallergy.2011.1.3.157
- Amin K. The role of mast cells in allergic inflammation. Respir Med. 2012;106(1):9–14. doi:10.1016/j.rmed.2011.09.007
- Okano M. Mechanisms and clinical implications of glucocorticosteroids in the treatment of allergic rhinitis. Clin Exp Immunol. 2009;158(2):164–173. doi:10.1111/j.1365-2249.2009.04010.x
- Varricchi G, Bagnasco D, Borriello F, Heffler E, Canonica GW. Interleukin-5 pathway inhibition in the treatment of eosinophilic respiratory disorders: evidence and unmet needs. Curr Opin Allergy Clin Immunol. 2016;16(2):186–200. doi:10.1097/ACI.0000000000000251
- Adachi T, Alam R. The mechanism of IL-5 signal transduction. Am J Physiol. 1998;275(3):C623–C633. doi:10.1152/ajpcell.1998.275.3.C623
- Niranjan R, Rajavelu P, Ventateshaiah SU, et al. Involvement of interleukin-18 in the pathogenesis of human eosinophilic esophagitis. Clin Immunol. 2015;157(2):103–113. doi:10.1016/j.clim.2015.01.007
- Arebro J, Ekstedt S, Hjalmarsson E, Winqvist O, Kumlien Georen S, Cardell LO. A possible role for neutrophils in allergic rhinitis revealed after cellular subclassification. Sci Rep. 2017;7:43568. doi:10.1038/srep43568
- Fransson M, Benson M, Wennergren G, Cardell LO. A role for neutrophils in intermittent allergic rhinitis. Acta Otolaryngol. 2004;124(5):616–620. doi:10.1080/00016480310015173
- Hosoki K, Itazawa T, Boldogh I, Sur S. Neutrophil recruitment by allergens contribute to allergic sensitization and allergic inflammation. Curr Opin Allergy Clin Immunol. 2016;16(1):45–50. doi:10.1097/ACI.0000000000000231
- Chen J, Zhou Y, Zhang L, et al. Individualized treatment of allergic rhinitis according to nasal cytology. Allergy Asthma Immunol Res. 2017;9(5):403–409. doi:10.4168/aair.2017.9.5.403
- Kirmaz C, Ozenturk Kirgiz O, Bayrak P, et al. Effects of allergen-specific immunotherapy on functions of helper and regulatory T cells in patients with seasonal allergic rhinitis. Eur Cytokine Netw. 2011;22(1):15–23. doi:10.1684/ecn.2011.0277
- Jordakieva G, Jensen-Jarolim E. The impact of allergen exposure and specific immunotherapy on circulating blood cells in allergic rhinitis. World Allergy Organ J. 2018;11(1):19. doi:10.1186/s40413-018-0197-0
- Ho J, Bailey M, Zaunders J, et al. Group 2 innate lymphoid cells (ILC2s) are increased in chronic rhinosinusitis with nasal polyps or eosinophilia. Clin Exp Allergy. 2015;45(2):394–403. doi:10.1111/cea.12462
- Nussbaum JC, Van Dyken SJ, von Moltke J, et al. Type 2 innate lymphoid cells control eosinophil homeostasis. Nature. 2013;502(7470):245–248. doi:10.1038/nature12526
- Skrindo I, Scheel C, Johansen FE, Jahnsen FL. Experimentally induced accumulation of Foxp3(+) T cells in upper airway allergy. Clin Exp Allergy. 2011;41(7):954–962. doi:10.1111/j.1365-2222.2011.03710.x
- Ayars GH, Altman LC, McManus MM, et al. Injurious effect of the eosinophil peroxide-hydrogen peroxide-halide system and major basic protein on human nasal epithelium in vitro. Am Rev Respir Dis. 1989;140(1):125–131. doi:10.1164/ajrccm/140.1.125
- Akuthota P, Weller PF. Eosinophils and disease pathogenesis. Semin Hematol. 2012;49(2):113–119. doi:10.1053/j.seminhematol.2012.01.005
- Park CS, Choi YS, Ki SY, et al. Granulocyte macrophage colony-stimulating factor is the main cytokine enhancing survival of eosinophils in asthmatic airways. Eur Respir J. 1998;12(4):872–878. doi:10.1183/09031936.98.12040872
- Yamaguchi Y, Suda T, Ohta S, Tominaga K, Miura Y, Kasahara T. Analysis of the survival of mature human eosinophils: interleukin-5 prevents apoptosis in mature human eosinophils. Blood. 1991;78(10):2542–2547.