Abstract
The transient receptor potential vanilloid 1 (TRPV1) is a thermoreceptor that responds to noxious temperatures, as well as to chemical agonists, such as vanilloids and protons. In addition, its channel activity is notably potentiated by proinflammatory mediators released upon tissue damage. The TRPV1 contribution to sensory neuron sensitization by proalgesic agents has signaled this receptor as a prime target for analgesic and anti-inflammatory drug intervention. However, TRPV1 antagonists have notably failed in clinical and preclinical studies because of their unwanted side effects. Recent reports have unveiled previously unrecognized anti-inflammatory and protective functions of TRPV1 in several diseases. For instance, this channel has been suggested to play an anti-inflammatory role in sepsis. Therefore, the use of potent TRPV1 antagonists as a general strategy to treat inflammation must be cautiously considered, given the deleterious effects that may arise from inhibiting the population of channels that have a protective function. The use of TRPV1 antagonists may be limited to treating those pathologies where enhanced receptor activity contributes to the inflamed state. Alternatively, therapeutic paradigms, such as reduction of inflammatory-mediated increase of receptor expression in the cell surface, may be a better strategy to prevent abrogation of the TRPV1 subpopulation involved in anti-inflammatory and protective processes.
TRPV1 receptor
Transient receptor potential vanilloid 1 (TRPV1), also known as the capsaicin receptor, was first cloned from rat dorsal root ganglion neurons using an expression-cloning screening strategy.Citation1 This newly cloned cDNA was first named VR1, for vanilloid receptor subtype 1. Because this receptor is a member of the transient receptor potential family of cation channels, it was given the name TRPV1 because it represented the first known member of the transient receptor potential vanilloid subfamily of transient receptor potential channels. To date, TRPV1 orthologs have been identified in eukaryotes, including human, rat, guinea pig, rabbit, mouse, dog, and porcine tissues, but not in prokaryotes. The ability of TRPV1 to respond to noxious stimuli and to be functionally sensitized by proinflammatory mediators has signaled it as a “pathological” receptor, having a significant role in the pain transduction pathway, and in the maintenance of inflammatory conditions in a variety of diseases and injury states.
TRPV1 structure and expression
TRPV1 is an 838-amino acid protein with a molecular weight of 95 kDa, consisting of six transmembrane segments, with an amphipathic pore-forming region between the fifth and sixth transmembrane segments, a large N-terminus intracellular domain, and a C-terminal cytosolic region (). Functional TRPV1 channels exist as homomultimers,Citation2 although functional heteroligomers may be formed between TRPV1 and TRPV3Citation3 or between TRPV1 and TRPV2,Citation4,Citation5 which may be responsible, at least in part, for the variable responses to agonists and antagonists. The 432-amino acid N-terminus contains at least six ankyrin repeats,Citation6,Citation7 which are essential for channel functionCitation8,Citation9 and for orchestrating a plethora of protein–protein interactions that govern the assembly of TRPV1-containing signalplexes.Citation10,Citation11 The 145-amino acid C-terminal contains subdomains involved in distinct channel functions. For instance, adjacent to the channel gate,Citation12 a highly conserved region known as the transient receptor potential domain, is involved in the functional coupling of stimuli sensing and gate opening.Citation13,Citation14 Furthermore, the C-terminus contains the molecular determinants for subunit tetramerization,Citation15,Citation16 two nucleotide-binding Walker-type sites,Citation17 as well as consensus sequences for modulation by phosphoinositides and protein kinases.Citation18,Citation19 More notably, this region has been suggested to hold the temperature sensor of the receptor.Citation20
Figure 1 A) Putative membrane topology of a transient receptor potential vanilloid 1 subunit displaying the location of residues involved in ligand-binding, proton activation, and post-translational modifications. The transient receptor potential vanilloid 1 domain, and calmodulin- and phosphatidylinositol-4,5-bisphosphate-binding domains are also depicted. B) Side view of the ribbon structural model of two opposite monomers of the transient receptor potential vanilloid 1 channel inserted into the lipid bilayer, after molecular dynamic simulation. The other two monomers are not shown for clarity.
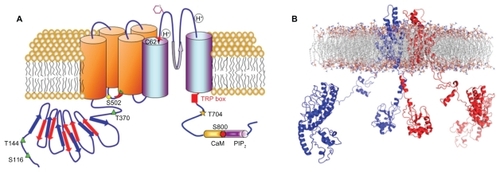
TRPV1 shows a wide tissue distribution. High levels of expression are observed in dorsal root ganglia, trigeminal ganglia, and nodose ganglia.Citation1 TRPV1 is predominantly expressed in small and medium diameter neurons, mainly in the peptidergic ones, that are important in the development of neurogenic pain and inflammation,Citation21 and to a lesser extent in the nonpeptidergic neurons that play a critical role in mediating chronicCitation22 and mechanical pain.Citation23 Although there is still a controversy about the central nervous system distribution of TRPV1,Citation24 several studies have demonstrated the expression of this channel in a wider diversity of brain regions, including the hypothalamus, cerebellum, cerebral cortex, striatum, midbrain, olfactory bulb, medulla, hippocampus, thalamus, and substantia nigra.Citation25 In non-neuronal tissues, TRPV1 expression is detected in keratinocytesCitation26 and melanocytes of the epidermis,Citation27 bladder urothelium,Citation28 smooth muscles,Citation29 glial cells, liver, polymorphonuclear granulocytes,Citation30 mast cells,Citation31 dendritic cells, and macrophages.Citation32
TRPV1 is a nonselective cation channel with near equal selectivity for Na+, K+, Li+, Cs+, and Rb+ ions,Citation1 but moderate selectivity for divalent cations. When activated by capsaicin, the permeability of Mg2+ and Ca2+ relative to Na+ (Px/PNa) is roughly 5 and 10, respectively.Citation1,Citation33,Citation34 Lower Px/PNa values of 3–4 are reported when the channel is activated by heat.Citation35 TRPV1 is also highly permeable to protons and large polyvalent cations, suggesting the existence of a large pore. Several amino acids in the putative pore-forming region between the fifth and sixth transmembrane segment domains are implicated in cation selectivity. Mutation of Glu-648 (E648A) reduces Mg2+ permeability and increases Ca2+ permeability. Mutation of Asp-646 (D646N) reduces Mg2+ permeability and blockade by the cationic dye, ruthenium red.Citation36
The single-channel conductance of capsaicin-activated channels is approximately 90–100 pS at positive potentials. At negative potentials (−60 mV), the conductance is significantly lower, with values of approximately 50 pS.Citation1 TRPV1 currents exhibit significant outward rectification due to a combined effect of voltage on both channel conductance and open probability.Citation37
TRPV1 has different modes of activation
TRPV1 is a polymodal channel, activated by physical and chemical stimuli, including heat, vanilloids, lipids, spider toxins, protons, cations, and voltage.Citation35,Citation38,Citation39 The channel is activated by noxious temperatures with a threshold of approximately 43°C,Citation1 and a temperature-dependent gating characterized by a Q10 ≥ 20 (Q10 is used to estimate the temperature dependence of channel gating).Citation40 The temperature threshold is highly influenced by other ligands that act allosterically and by the receptor phosphorylation state. Thus, when simultaneously activated by other ligands, the threshold may decrease down to 20°C. It has been proposed that temperature regulates TRPV1 by changing the intrinsic voltage sensitivity of the channel.Citation37 The temperature sensitivity of this channel is allosterically linked to chemical and voltage activation.Citation20 Although the mechanisms underlying heat activation remain unclear, a role of the C-terminus and the outer pore region has been proposed.Citation14,Citation41
TRPV1 is activated by capsaicin, the pungent component of hot chili peppers. Capsaicin and related compounds, including resiniferatoxin and olvanil, are highly lipophilic and share a structural similarity to several endogenous fatty acid derivatives that have also been identified as TRPV1 agonists.Citation42,Citation43 These include anandamide (an endocannabinoid), N-arachidonoyl dopamine, oleoyldopamine, 12-hydroperoxyeicosatetraenoic acid (a lipoxygenase product), and 18–20 carbon N-acylethanolamines.Citation44 Vanilloids interact at intracellular regions of TRPV1, as implied by a membrane-impermeable charged capsaicin analog that is only effective when applied cytosolically.Citation45 Consistent with this observation, several intracellular molecular determinants of capsaicin binding have been identified. The amino acid residues, Arg-114 in the N-terminus and Glu-761 in the C-terminal domain, play a key role in ligand binding.Citation46 In addition, Tyr-511 and Ser- 512 located between the second and third transmembrane segments are also critical for vanilloid binding and channel activation,Citation47 and Thr-550 has also been pointed towards as involved in structuring the vanilloid binding site in rat and human TRPV1 channels.Citation48
A pH lower than 5.0 at the extracellular side of the channel activates TRPV1 ion channels. Actually, ligand-, voltage-, and temperature-evoked gating are potentiated by mildly acidic extracellular pH.Citation49 Two glutamate residues located near the extracellular pore-forming region appear critical for proton regulation, ie, Glu-648, at the loop between the fifth and sixth transmembrane segments, is involved in direct activation of the channel by strong pH (pH 4), while Glu-600 located at the end of the fifth transmembrane segment is important for the response of the channel to mildly acidic external conditions (pH 6.5).Citation49 Nevertheless, neutralization of Glu-600 gives rise to a constitutively active channel at 37°C.Citation49
In addition to protons, positively charged compounds are also able to activate TRPV1, suggesting a generalized activation mechanism based primarily on neutralization of Glu-600. External cations tend to enhance agonist-evoked currents, and divalent cations at high (>10 mM)Citation34 or even at physiological concentrationsCitation50 gate the channel directly. Polyvalent cations are even more potent channel regulators. For instance, Gd3+ and the polyamine, spermine,Citation51,Citation52 sensitize and activate TRPV1 at micromolar concentrations. These actions may involve interactions at multiple acidic residues, ie, Glu-600, Glu-648, and Asp-646.
TRPV1 also has a voltage-dependent gating. The channel is activated, at least partially, at strong positive potentials and is deactivated at negative potentials. The sensitivity of voltage-dependent activation and deactivation depends on the recording temperature and on the presence of agonists. In the absence of TRPV1 activators, strong membrane depolarization is required to activate the channel (V0.5 of +150 mV at 21°C), whereas in the presence of agonists, much smaller depolarization suffices to gate the channel, namely V0.5 of 0 mV at 37°C, and +10.6 mV at 21°C in the presence of 50 nM capsaicin.Citation37 Thus, the heat or ligand sensitivity of TRPV1 may reflect a shift in its intrinsic voltage dependence. Consequently, the temperature threshold for TRPV1 activation is not constant, but fluctuates depending on the membrane potential. The voltage sensor remains unknown, although the fourth transmembrane segment has been signaled as a putative candidate to hold it. However, unlike voltage-gated channels, TRPV1 and other transient receptor potential channels lack an array of charged residues in their transmembrane segment domains.
From the aforementioned observations, it appears obvious that various activators of TRPV1 potentiate the effect evoked by others, leading to enhanced activity, suggesting a coupling of their receptor sites. This gating cooperativity of various ligands seems synergistic rather than additiveCitation20 and, given the polymodal and synergistic modes of activation, implies that the TRPV1 ion channel act as an “integrator” of exogenous stimuli.Citation35 In fact, TRPV1 acts similarly in relation to endogenous agents, which makes it of particular relevance in the context of inflammation, given the wide variety of inflammatory agents generated in inflamed conditions.
In the continuous presence of an activating stimulus, TRPV1 undergoes desensitization. This phenomenon can occur rapidly after a prolonged single application of an agonist, or slowly following repeated agonist applications (also known as tachyphylaxia). Receptor desensitization is believed to occur predominantly via a Ca2+-dependent process because it is largely abolished in the absence of Ca2+. However, it should be noted, that some Ca2+-independent desensitization also occurs, especially with heat activation.Citation53 The Ca2+-dependent mechanism arises because of the high TRPV1 Ca2+ permeability, allowing Ca2+ influx to activate an inhibitory process. Indeed, fast desensitization was significantly reduced in a TRPV1 mutant that possesses markedly reduced Ca2+ permeability.Citation33 Furthermore, desensitization is attenuated by inhibitors of calcineurin, a Ca2+-activated phosphatase, thus linking desensitization to a dephosphorylation event.Citation54 In addition, Ca2+ may signal via calmodulin, which interacts with TRPV1 at the N-terminal and C-terminal regions (positions 189–222 and 767–801). Indeed, disruption of the calmodulin C-terminal region partially inhibits fast desensitization.Citation55
Regulation of TRPV1 channel activity
There is increasing evidence that TRPV1 is subjected to complex regulation manifested at several levels, from gene expression to post-translational modification and formation of receptor heteromers, as well as from subcellular compartmentalization and association with regulatory proteins to many second messengers.Citation11
Limited information is available about what controls TRPV1 transcription in nociceptors. Two functional TRPV1 promoter regions and transcription initiation sites have been identified in the rat, ie, a distal promoter region, P1, and a second more proximal promoter region, P2.Citation56 The P1 region containing a classic TATA box and a downstream transcription initiation site directs the strongest promoter activity within the 233-bp core fragment. The proximal promoter region, P2, which lacks a TATA box, contains an associated transcription initiation site that corresponds to the consensus sequence known as the “initiator” element. Alternate use of dual promoters may represent an important aspect of how TRPV1 gene expression can be dynamically regulated. Nerve growth factor induces activation of the GTPase Ras, which is coupled to the activation of both transcription and translation of TRPV1.Citation57 Nerve growth factor positively regulates transcriptional activity of both rat TRPV1 promoters.
A large body of evidence indicates that post-translational modifications of TRPV1, such as phosphorylation mediated by protein kinase A, protein kinase C, and calmodulin-dependent protein kinase, increase its activity. Phosphorylation at Ser-116 in the N-terminus of TRPV1 is pivotal in protein kinase A-mediated downregulation of TRPV1 desensitization.Citation58 In addition, Thr-144, Thr-370, and Ser-502 are important in protein kinase A-mediated phosphorylation/sensitization of the channel. Moreover, protein kinase C-mediated phosphorylation of TRPV1 not only potentiates capsaicin-evoked or proton-evoked responses, but also reduces its temperature threshold, such that receptors are active under physiological conditions (37°C).Citation59 Two serine residues on TRPV1, Ser-502 and Ser-800, have been recognized to be important in protein kinase C-mediated effects. In addition to this direct effect, protein kinase C can also produce phosphatidylinositol 4,5-bisphosphate (PIP2) hydrolysis increasing TRPV1 activity, although PIP2 has been proposed to be involved in sensitization of these channels by proinflammatory agents.Citation59 Calmodulin-dependent protein kinase-mediated phosphorylation of TRPV1 at Ser-502 and Thr-704 plays an important role in channel activation in response to capsaicin application.Citation60 In addition, calcineurin-mediated dephosphorylation at the same sites can produce TRPV1 desensitization.Citation61 Similarly, the nonreceptor cellular tyrosine, c-Src kinase, positively regulates TRPV1 channel activity by tyrosine phosphorylation.Citation61
In addition to phosphorylation, the activity of TRPV1 may be regulated by N-glycosylation,Citation62 given that extracellular Asn-604 has been identified as a glycosylation site.Citation63 Similarly, adenosine 5′ triphosphate may allosterically modulate TRPV1 by direct interaction with the nucleotide-binding Walker-type domains and increasing vanilloid-induced channel activity.Citation9 Modulation of the redox state also impacts the physiological activity of TRPV1, possibly involving the Cys-621 amino acid residue located on the extracellular surface.Citation64
Another essential pathway that influences TRPV1 activity is the formation of signalplexes, or the physical assembly of signaling molecules into discrete macromolecular entities. Citation11 Several signaling proteins have been described as TRPV1-interacting proteins, that could be part of a “TRPV1 receptome” modulating nociceptor activity. As mentioned, TRPV1 associates with intracellular signaling enzymes, including protein kinase A, protein kinase C, Src, inositol 1,4,5-trisphosphate, and calmodulin-dependent protein kinases, and also with calcineurin 2B phosphatase.Citation65 It may also interact with the purinergic P2X3 receptor,Citation66 calmodulin,Citation67 the membrane protein, Pirt,Citation68 the scaffolding protein, AKAP79/150,Citation69 and with cytoskeleton proteins like tubulin.Citation70 Protein kinases modulate channel gating by post-translational modification involving the phosphorylation/dephosphorylation of specific residues that, in turn, lead to a decrease in the temperature threshold of channel activation and a potentiation of its activity, by either destabilizing the closed and desensitized states and/or stabilizing the open state. Other proteins that bind to TRPV1 are snapin and synaptotagmin IX, two components of the SNARE complex that mediates Ca2+-dependent exocytosis.Citation71 Although the precise role of snapin and synaptotagmin IX binding to TRPV1 remains elusive, it could be involved in sorting the receptor into vesicles that will be exocytosed through regulated exocytosis or in promoting channel recruitment to the plasma membrane under inflammatory conditions.
Several proteins that regulate folding (chaperones), protein biosynthesis, surface expression, and channel function have been described to associate with thermotransient receptor potentials. Recently, the γ-aminobutyric A receptor-associated protein, a small cytosolic protein initially described by its ability to interact with the γ subunit of the GABAA receptor,Citation72 was pointed towards as a TRPV1 interacting partner with the cytosolic N-terminal domain of the channel. Citation73 Noteworthy, in heterologous systems, γ-aminobutyric A receptor-associated protein expression significantly augmented the levels of TRPV1 and its targeting to the plasma membrane, where it appears to favor the formation of receptor clusters. Functionally, γ-aminobutyric A receptor-associated protein appears to induce a decrease in channel activity.Citation73
TRPV1 in inflammation
Inflammation is the physiological response to tissue injury caused by pathogens or harmful agents, and is clinically characterized by swelling, redness, heat, pain, and loss of function of the affected tissue or organ. This response is a complex process perfectly orchestrated by several cell types and chemical mediators, which initiate and regulate the necessary mechanisms to remove injurious agents and repair the affected area. The cellular components include circulating monocytes, macrophages, neutrophils, lymphocytes, and dendritic cells, while the humoral components include cytokines and other chemical substances that destroy pathogens or act as mediators for other cells. When tissue damage occurs, resident immune cells, such as macrophages or dendritic cells, are activated and release mediators in order to initiate the inflammatory response. Usually, during acute inflammation, the magnitude of the inflammatory response is locally adjusted to the injurious condition and finally resolved, maintaining homeostasis. However, an imbalance of the regulatory mechanisms is the cause of inflammation as a pathological process and leads to chronic inflammatory states. Regulatory mechanisms of inflammation include mediators of immune, vascular, or neural origin that maintain the inflammatory process within the physiological range. The role of TRPV1, as a major player in the process of neurogenic inflammation, has been traditionally considered to be neuronal. However, the expression of the channel in immune cells also suggests a contribution to the immune response.Citation30–Citation32,Citation74
Inflammatory mediators are released at the site of injury from immune cells, such as chemokines, cytokines, prostaglandins, bradykinin, or growth factors, as well as from sensory neurons that secrete the neuropeptides, substance P and calcitonin gene-related peptide.Citation75 Some of these mediators are able to activate directly local sensory neurons responsible for transducing the painful sensation that, paradoxically, is necessary to react and to avoid or minimize further damage.Citation76–Citation80 In addition, inflammatory agents are responsible for nociceptor sensitization changing the perception of stimuli, which leads to hyperalgesia (exaggerated response to a mild noxious stimulus) and/or to allodynia (response to a non-noxious stimulus), further minimizing additional damage and facilitating tissue repair. In chronic conditions, this process is exacerbated by synaptic changes at the spinal cord, a process known as central sensitization.Citation81 Neuronal sensitization is believed to play a pivotal role in the development and maintenance of chronic pathological pain conditions.Citation82
Inflammatory regulation of TRPV1
Peripheral sensitization of TRPV1 by proinflammatory agents is mediated by different molecular mechanisms, which include long-term upregulation of TRPV1 expression, but also acute functional modification of the channel (). Indeed, increased expression of the channel has been shown in several chronic inflammatory diseases.Citation83–Citation85 This process is also mediated by fast mobilization from a subcellular vesicular reservoir located near the plasma membrane that is recruited by SNARE-dependent exocytosis.Citation11,Citation86
Figure 2 Regulation of transient receptor potential vanilloid 1 function and expression by proinflammatory mediators. A) Acute post-translation modification of transient receptor potential vanilloid 1 function. Activation of phospholipase C/protein kinase C, protein kinase A, calmodulin-dependent protein kinase, and other intracellular signaling cascades increase transient receptor potential vanilloid 1 activity and cytosolic Ca2+ levels. B) Increase of transient receptor potential vanilloid 1 expression by proinflammatory agents. Rapid receptor translocation to the cell surface from the vesicular reservoir (left side). Long-term upregulation of protein levels by transcription/translation process (right side).
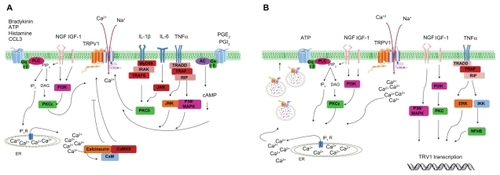
TRPV1 sensitization by nerve growth factor has been well documented, and is a good example of all the inflammatory potentiation strategies described earlier for the modulation of TRPV1 function and expression. Nerve growth factor increases TRPV1 transcription and transport to the peripheral nociceptor terminal, a process mediated by the p38/mitogen-activated protein kinase signaling pathway.Citation87,Citation88 Acute regulation of TRPV1 by nerve growth factor leads to phospholipase C activation and PIP2 hydrolysis. In parallel, nerve growth factor activates phosphatidylinositol 3-kinase-protein kinase C epsilon and calmodulin-dependent protein kinase signaling cascades, increasing TRPV1 opening probability and its translocation to the cell surface from the vesicular pool.Citation89–Citation91 Similar to nerve growth factor, insulin growth factor-1 also enhances TRPV1 membrane currents through phosphatidylinositol 3-kinase and protein kinase C pathways, increasing channel activity and receptor translocation to the cell surface,Citation92 inducing long-term overexpression of TRPV1.Citation93 Both nerve growth factor and insulin growth factor-1 provoke TRPV1 plasma membrane translocation by SNARE-dependent neurosecretion, as was demonstrated by the blockade of Ca2+-dependent neuronal exocytosis with a botulinomimetic peptide which abolished TRPV1 potentiation in dorsal root ganglion neurons.Citation94
Cytokines, such as tumor necrosis factor alpha, interleukin-1β, and interleukin-6, can also regulate TRPV1 function, increasing neuronal excitability.Citation76,Citation95 For instance, the receptors for tumor necrosis factors alpha (ie, TNFR1 and TNFR2) are coexpressed with TRPV1 in sensory neurons, where they can also produce rapid and long-term modification of TRPV1 function. This cytokine increases TRPV1 expression on dorsal root ganglion and trigeminal ganglion neurons via the extracellular signal-regulated kinase pathway.Citation96,Citation97 Additionally, tumor necrosis factor alpha rapidly sensitizes TRPV1 activity and enhances the Ca2+ influx induced by capsaicin. This rapid mechanism seems to be mediated by p38/mitogen-activated protein kinase and the c-jun N-terminal kinase pathway, but not by extracellular signal-regulated kinase.Citation97–Citation99 Although protein kinase C phosphorylation seems also to be implicated, the exact mechanism remains unknown.Citation76,Citation100–Citation102 Tumor necrosis factor alpha can also activate the TRPA1 receptor, which has been implicated in maintaining inflammation-related pain.Citation103 TRPA1 is coexpressed in a subset of TRPV1-expressing nociceptors in trigeminal and dorsal root ganglion neuronsCitation104 and functions to detect products of tissue injury, inflammation, and oxidative stress that cause pain and neurogenic inflammation. Citation105 Under conditions of inflammation or nerve injury, expression of TRPA1 is persistently increased, concurrent with TRPV1.Citation106
Rapid sensitization of TRPV1 currents by interleukin-1β has also been shown to be mediated by protein kinase C activity,Citation107 via a mechanism independent of TRPV1 surface translocation by SNARE-dependent exocytosis.Citation94 Although little is known about the ability of interleukin-6 to sensitize TRPV1,Citation78,Citation108 exposure of dorsal root ganglion cultures to interleukin-6 increases TRPV1 response to heat by a mechanism that involves Janus kinase and protein kinase C.Citation109
Other inflammatory mediators, such as bradykinin, prostaglandin E2, adenosine 5′ triphosphate, and histamine, also sensitize TRPV1. Bradykinin induces excitation and sensitization of TRPV1 to heat via the protein kinase C pathway.Citation110–Citation112 In the same way, TRPV1 potentiation by adenosine 5′ triphosphateCitation113 or by histamineCitation114 is mediated via the phospholipase C/protein kinase C pathway. However, only adenosine 5′ triphosphate has been shown to mobilize TRPV1 to the plasma membrane mediated by the SNARE complex.Citation94 Mechanisms involved in TRPV1 sensitization by prostaglandin E2 and prostaglandin I2 are through phosphorylation by protein kinase A,Citation55 the receptor anchoring for which seems to be mediated by the protein, AKAP150,Citation115 and also by protein kinase C.Citation55
The cellular mechanisms underlying chemokine-induced excitation of sensory neurons include potentiation of TRPV1, in addition to inhibition of K+ conductance.Citation95 CCL3 via the CCR1 receptor enhanced the response of dorsal root ganglion neurons to capsaicin, and decreased the response to hot-plate latency in mice. CCL3-mediated TRPV1 sensitization was reduced by phospholipase C and protein kinase C inhibitors.Citation116 Activation of other chemokine receptors, such as CCR2, also expressed in the dorsal root ganglia, produced sensitization of TRPV1 by phospholipase C/protein kinase C phosphorylation.Citation117
TRPV1 in inflammatory conditions
Besides the direct effect of inflammatory mediators on TRPV1, activation of nociceptors also induces the release of neuropeptides which act both autocrinally on the terminals and paracrinally on target cells, such as mast, immune, and vascular smooth muscle cells.Citation118,Citation119 These peptides contribute to the destruction of the harmful agent and to the repair of damaged tissue. For instance, when the neuropeptides calcitonin gene-related peptide and substance P are released from sensory neurons, their vasodilatory effects facilitate the arrival of more immune cells and proinflammatory mediators at the site of injury, which contributes to plasma extravasation and swelling. In fact, direct activation of sensory nerves is enough to induce an inflammatory response without the presence of pathogens or tissue injury, a process known as neurogenic inflammation or sterile inflammation.Citation120
TRPV1 is expressed in sensory neurons, mainly in peptidergic neurons, found in many tissues close to blood vessels, epithelia, and vascular smooth muscle.Citation121,Citation122 Release of calcitonin gene-related peptide and substance P from sensory neurons is induced by TRPV1 activation via a wide variety of physical and chemical stimuli.Citation123 Sensitization of TRPV1 by inflammatory mediators increases the release of these neuropeptides from a vesicle reservoir.Citation97,Citation124 Due to the proinflammatory effects of these neuropeptides, TRPV1 activation has been long considered as a proinflammatory receptor. However, other neuropeptides with anti-inflammatory properties, such as somatostatin, can also be released as a consequence of Ca2+ influx through the TRPV1 channel.Citation125,Citation126
TRPV1 acts as a transducer of noxious thermal and chemical stimuli in nociceptive sensory neurons, and is vital in mediating enhanced heat sensitivity during inflammation. Preclinical and clinical studies suggest that the TRPV1 receptor is an important component of several disease states, such as pain (inflammatory, visceral, cancer, and neuropathic), airways disease (including chronic cough), inflammatory bowel disease, interstitial cystitis, urinary incontinence, pancreatitis, and migraine.Citation127
Acute and chronic arthritis is characterized by debilitating pain, and by an increment in the levels of neuropeptides in synovial fluid. Due to the role of TRPV1 as an integrator of multiple noxious stimuli as well as its presence in neuropeptide-containing fibers that are present in the knee joint synovium and adjacent bone, this channel has been implicated in the pathological symptoms of acute and chronic arthritis, although the precise mechanism is unclear.Citation128 Keeble et al studied the vascular and hyperalgesic components of joint inflammation in wild-type and TRPV1 knockout mice after intra-articular injection of Freund’s complete adjuvant, and demonstrated that knee swelling and vascular hyperpermeability were significantly lowered in the joints treated with Freund’s complete adjuvant in TRPV1 null mice. Furthermore, intra-articular injection of tumor necrosis factor alpha in these mice produced decreased thermal hyperalgesia and joint swelling, indicating a critical role of tumor necrosis factor alpha and TRPV1 in the pathophysiology of rheumatoid arthritis.Citation129
Cancer pain is a significant clinical problem because it is the first symptom of the disease in approximately 20%–50% of all cancer patients. Bone is the most common site of origin of chronic pain in patients with metastatic lung, prostate, and breast cancers or myeloma.Citation130 There are at least three mechanisms in bone cancer that may contribute to the activation and sensitization of TRPV1 expressed by sensory fibers that innervate the tumor-bearing bone.Citation131 The first is acidosis produced by osteoclasts, the principal bone-resorbing cells, and by lysis of tumor cells that have a lower intracellular pH than normal cells.Citation132 The second is mediated by products released from cancerous tissue, like bradykinin, adenosine 5′ triphosphate, and nerve growth factor, which can modulate TRPV1 function indirectly via activation of second-messenger signaling pathways.Citation130 Because the bone receives a rich sensory innervation by fibers that express TRPV1,Citation133 production of these proalgesic agents may also sensitize TRPV1 channels, thereby generating a state of hyperalgesia and/or allodynia. Finally, the third mechanism is mediated directly by tumor-induced injury to primary afferent neurons.Citation134 In a recent study, it has been shown that activation of TRPV1 was involved in bone cancer pain.Citation135 The investigators found increases in TRPV1 protein levels and in the number of TRPV1-positive neurons in the dorsal root ganglia from a murine model of bone cancer. In support of a role of TPRV1 in bone cancer pain, it has been demonstrated that a receptor antagonist significantly attenuates painful symptoms.Citation130
Itch, a principal symptom in skin diseases, is an important skin manifestation of systemic diseases, and one of the most debilitating symptoms in allergic and atopic dermatitis. Citation136 It can be triggered by localized, systemic, peripheral, or central stimuli, and there are numerous pruritogenic substances, including neuropeptides, cytokines, proteases, and histamine. Citation137 Less is known about pathophysiological specificity among the different diseases, but cross-talk between neuron terminals and dermal mast cells is being recognized as an important mechanism involved in pathogenesis. TRPV1-expressing primary afferents generate responses to pruritogenics via multiple mechanisms, like PLCβ3 activation.Citation138 In addition, keratinocytes express a wide range of mediators and receptors involved in itch, and TRPV1 activation by them results in the release of pruritogenic mediators, as well as cellular proliferation, differentiation, and apoptosis.Citation139 Histamine, the best known pruritogenic agent, induces itch by activating PLA2, lipoxygenase and the TRPV1 signaling pathway, as is shown by the decrease in histamine-induced scratching in TRPV1-deficient mice.Citation140
Protective role of TRPV1 against inflammation
Cumulative evidence suggests that TRPV1 may have an anti-inflammatory action in some pathological conditions. Indeed, the number of diseases in which TRPV1 plays a protective role is expanding. For instance, TRPV1 has been shown to have a protective role against inflammatory conditions in the cardiovascular system, and it has been implicated in protecting against ischemia/reperfusion-induced inflammation of the heart.Citation141 A similar action has been reported for liverCitation142 and kidneyCitation143 pathologies, thus emphasizing a new emerging anti-inflammatory role for TRPV1.
The TRPV1 receptor also plays a critical modulatory role in contact dermatitis, a chronic allergic condition typified by skin inflammation and itching.Citation144 The genetic disruption of TRPV1 channels or blockade of the TRPV1-dependent sensory neurogenic component by resiniferatoxin increased the inflammatory response in an ear murine model of contact dermatitis. This enhancement suggests that capsaicin-sensitive neurons expressing the TRPV1 channel may act to downregulate the hypersensivity, possibly by influencing the immune state of the skin.
Another protective function of TRPV1 against inflammatory conditions has been reported in the pathological condition of colitis,Citation145 one of the disorders under the collective heading of gastrointestinal disturbances referred to as chronic inflammatory bowel diseases. Immunoreactive TRPV1 fibers have been detected on nerve terminals within the myenteric ganglia and the interganglionic fibers throughout the gastrointestinal tract.Citation146 Using a murine model produced by the infusion of 2,4-dinitrobenzene sulfonic acid through the rectum of mice,Citation147 it was reported that the TRPV1 null mice exhibited higher levels of inflammation than wild-type animals, indicating a protective role of TRPV1 channels in the initiation of this inflammatory condition.
Although this protective action of TRPV1 may be surprising, given the widely known proinflammatory activity of this channel, it should be considered that the main mechanisms involved in the development of chronic inflammation can be drastically different in each inflammatory disease. For instance, TRPV1 may mediate the release of both proinflammatory neuropeptides (substance P and calcitonin gene-related peptide) in some conditions, and of anti-inflammatory (somatostatin) peptides in others, depending on which subpopulation of nociceptors is more abundant in the affected tissue. Likewise, the immune cells and inflammatory mediators involved will depend on the specific inflammatory process and the affected tissue. Moreover, this balance usually changes during the development of pathology according to the time course or state of the disease. Therefore, the contribution of TRPV1 should be analyzed in detail for each inflammatory condition along with the mechanism involved in the specific inflammatory process. This evaluation will become essential because abrogation of the anti-inflammatory role of TRPV1 by potent antagonists may lead to aggravation of the disease or to the appearance of side effects.
Inflammatory process of sepsis
Sepsis is defined as the systemic inflammatory response elicited by an infection. The clinical manifestations of sepsis are abnormality in at least two of the following: body temperature (hyperthermia or hypothermia), heart rate (tachycardia), respiratory rate (tachypnea), and white blood cell count (leukocytopenia, leukocytosis or presence of immature forms greater that 10%). Increasingly grave stages of the systemic inflammatory response to an infection are severe sepsis and septic shock. Sepsis is considered when signs of organ dysfunction are present, while septic shock is defined by the presence of hypotension or hypoperfusion which finally leads to multiple organ failure.Citation148,Citation149 Despite the advances in antibiotics and critical care, severe sepsis remains the leading cause of death in intensive care units, in part because antibiotics cannot control systemic inflammation. In fact, the clinical symptoms of severe sepsis are not exclusively due to infection and can be also triggered by trauma, ischemia, severe injury, burns, or pancreatitis, which participate in the pathogenesis of this systemic inflammatory response syndrome.
The pathogenesis of sepsis is characterized by an excessive production of inflammatory mediators, mainly cytokines, chemokines, lipid mediators, and oxygen radicals. In a normal inflammatory response, these molecules are necessary to enhance leukocyte infiltration at the site of infection, to destroy the pathogen and repair tissue damage. However, in sepsis, this uncontrolled production of inflammatory mediators ends up with excessive vasodilatation, capillary leakage, hypotension, tissue injury, and finally lethal multiple organ failure.Citation150–Citation152 Clinical and experimental studies have emphasized the role of apoptosis in sepsis. Programmed cell death constitutes an active process to control cell removal, but also plays an important role in several pathological states. Activation of intracellular cascades finally leads to DNA degradation, a process regulated by a cascade of caspases, which are critical molecules in programmed cell death.Citation153,Citation154 Accelerated apoptosis of lymphocytes has been observed in animal models of sepsis and in autopsies of patients who died from sepsis.Citation155
Different studies have demonstrated that the immune system is not the only system activated in sepsis. For instance, it is well known that a complex interaction exists between inflammation and coagulation in sepsis. The inflammatory response in sepsis skews the balance to a procoagulant state, promoting thrombus and clot formation. Indeed, patients with increased coagulation factors and reduced anticoagulation factors as a result of sepsis are prone to thrombus formation, compromising tissue perfusion and driving towards organ failure.Citation151,Citation156
In addition to activation of the coagulation cascades, extensive bidirectional communication exists between the immune and nervous systems in all inflammatory processes, which involves a huge diversity of molecular mechanisms.Citation156,Citation157 Release of inflammatory molecules can activate or influence sensory nerve function, which, in turn, can stimulate or inhibit the immune system by the release of neurotransmitters. In patients with sepsis, plasma levels of substance P and calcitonin gene-related peptide are significantly increased, providing evidence that sensory nerves are activated in this pathology.Citation158–Citation160 These bioactive neuronal agents are able to induce inflammation by directly acting on immune cells, mast cells, vascular smooth muscle, or other cell types.Citation118,Citation119 Substance P is known to increase vascular permeability,Citation161 and calcitonin gene-related peptide is a potent vasodilator and hypotensive agent.Citation162 Therefore, both neuropeptides have been mainly considered to be involved in the development of inflammation. However, it has also been shown that calcitonin gene-related peptide can mediate anti-inflammatory and immunosuppressive activities. For instance, it modulates cell adhesion and migration, increases some anti-inflammatory mediators, such as interleukin-10 or prostaglandin I2, and inhibits proinflammatory mediators, such as tumor necrosis factor alpha, among others.Citation163–Citation168 Consistent with these findings, administration of calcitonin gene-related peptide attenuated the development of some inflammatory and organ failure models.Citation169,Citation170 Neuropeptides, such as somatostatin, have also been recently reported to be increased in plasma from septic patients,Citation171 but others, such as endothelin-1 or vasoactive intestinal peptide serum levels, seem to remain unmodified during sepsis.Citation172 However, despite these observations, progress towards understanding the potential involvement of sensory nerves in sepsis and defining the exact role of neuropeptides in the development of this pathology is quite limited.
Is TRPV1 involved in sepsis?
Cumulative evidence associates TRPV1 channel activity with a protective effect in experimental models of sepsis. The role of the TRPV1 channel in sepsis was first evidenced by using its known agonist, capsaicin, and its antagonist, capsazepine. Injection of capsaicin was shown to diminish mortality during abdominal sepsis, suggesting an important role for nociceptive system in the host response to infection.Citation173 A small dose of capsaicin reduced the systemic inflammatory response in septic rats by increasing anti-inflammatory cytokines and attenuating proinflammatory cytokines,Citation174 which was consistent with previous in vitro results in lipopolysaccharide-activated peritoneal macrophages.Citation175 In contrast, TRPV1 blockade with capsazepine reduced channel-mediated protection against endotoxin-induced hypotension and mortality in septic rats.Citation176 Later, similar beneficial effects were also shown in a different rat model of sepsis, where capsaicin significantly attenuated systemic inflammation and multiple organ damage caused by sepsis, and protected against mortality.Citation177 Other TRPV1 agonists and antagonists have also been evaluated in different models of sepsis, all showing a consistent decrease in the development of sepsis or a reduction in some of the pathological symptoms when TRPV1 is activated.Citation178–Citation180
Recent elegant experiments performed in TRPV1 knockout mice lend further support to the protective role of the vanilloid channel in the onset of sepsis. In these animals, there was an enhanced development of the pathological features and biomarkers of the systemic inflammatory response. Early onset, decreased body temperature, and enhanced hypotension were shown, together with an increased level of some inflammatory mediators in peritoneal exudates.Citation181,Citation182 At the same time, protective effects were also shown in another septic model in which TRPV1 null mice showed greater infiltration, more histological lesions, bronchial hyperactivity, and increased myeloperoxidase levels in the lungs.Citation183
The neuropeptides involved in the protective effects of TRPV1 in sepsis remain to be exactly defined. Although it was initially suggested that TRPV1 mediated the effects of sepsis via substance P and the NK1 receptor,Citation176 the regulatory role of the channel in sepsis was shown to be independent of substance P in mice lacking TRPV1.Citation181 Alternatively, because calcitonin gene-related peptide, a potent vasodilator, was shown to be elevated in endotoxin-treated rats,Citation184 it has been considered a critical factor in the development of septic shock.Citation185 Moreover, an essential role has been claimed for somatostatin, because somatostatin receptor blockade aggravates sepsis in the lungs of wild-type mice, while the process is attenuated by injection of this neuropeptide in TRPV1 knockout mice.Citation183
It should be noted that these results have been obtained from different models of sepsis in rats or mice, using several research and pharmacological tools which could explain the differences observed in neuropeptides participating in the effects of TRPV1. Moreover, the tissue parameters and/or biomarkers analyzed in each model are different, which could also contribute to the observed differences in the role of neuropeptides. In addition, it should not be forgotten that endotoxin-induced fever was shown to be initiated via the TRPV1 channel.Citation186,Citation187 However, an overall protective role of TRPV1 in sepsis has been proposed in most of these studies, and the increased levels of some neuropeptides in septic patients could be due to compensatory mechanisms of the organism when trying to control the systemic inflammatory process.
Although understanding of the pathogenesis of inflammation and sepsis has improved, this has not translated into clinical benefit. Therapies for severe sepsis are mainly focused on eradication of infection and on maintenance of systemic perfusion. Despite advances in adjuvant treatments, mortality remains high. In past decades, therapeutic attempts have been focused on inflammatory mediators and processes, but they have failed to translate into efficacy in clinical trials, although animal models have shown promising and successful results. The benefit of corticosteroid therapy in severe sepsis and septic shock remains controversial. Activated protein C, one of the coagulation factors, is the only treatment for sepsis approved by the Food and Drug Administration, which is projected to be useful only in a small subset of patients with severe sepsis. The proposed protective role of TRPV1 implies that antiseptic treatments should preserve its channel activity. Thus, until the role of TRPV1 in sepsis is well understood, potent channel antagonists should be used with caution to treat the septic inflammatory process.
Conclusion
It is becoming clear that TRPV1 contributes to the pathophysiology of inflammatory processes. Intriguingly, this channel may have both a proinflammatory and anti-inflammatory action, depending on the disease. Thus, although it was widely accepted that TRPV1 blockers will become a new generation of anti-inflammatory and analgesic drugs to treat a plethora of human diseases, their clinical use must be reconsidered, because of the newly identified protective roles assigned to TRPV1. We are learning that some of the protective anti-inflammatory effects of TRPV1 were most probably ignored or misinterpreted. Therefore, the better we understand how TRPV1 works and how it contributes to human physiology and pathology, the more challenging it will be to find compounds that target pathological proinflammatory TRPV1 channels, without altering physiologically working and anti-inflammatory subpopulations of channels. Perhaps targeting inflammatory expression and/or recruitment of channels may provide a superior therapeutic paradigm to attenuate inflammation.
Acknowledgments
We are grateful to the members of our laboratory for their continuous support and collaboration. This work was supported by grants from the Ministry of Science and Innovation to AF-M, to JMGR, RP-C, from Consolider-Ingenio to AF-M, JMG-R, and RP-C, from La Marató de TV3 to AF-M and RP-C, and from la Generalitat Valenciana Prometeto to AF-M.
Disclosure
The authors report no conflicts of interest in this work.
References
- CaterinaMJSchumacherMATominagaMRosenTALevineJDJuliusDThe capsaicin receptor: A heat-activated ion channel in the pain pathwayNature199738966538168249349813
- KedeiNSzaboTLileJDAnalysis of the native quaternary structure of vanilloid receptor 1J Biol Chem200127630286132861911358970
- SmithGDGunthorpeMJKelsellRETRPV3 is a temperature-sensitive vanilloid receptor-like proteinNature2002418689418619012077606
- LiapiAWoodJNExtensive co-localization and heteromultimer formation of the vanilloid receptor-like protein TRPV2 and the capsaicin receptor TRPV1 in the adult rat cerebral cortexEur J Neurosci200522482583416115206
- RutterARMaQPLeveridgeMBonnertTPHeteromerization and colocalization of TrpV1 and TrpV2 in mammalian cell lines and rat dorsal root gangliaNeuroreport200516161735173916237318
- TominagaMTominagaTStructure and function of TRPV1Pflugers Arch2005451114315015971082
- PhelpsCBProckoELishkoPVWangRRGaudetRInsights into the roles of conserved and divergent residues in the ankyrin repeats of TRPV ion channelsChannels (Austin)20071314815118690026
- PhelpsCBWangRRChooSSGaudetRDifferential regulation of TRPV1, TRPV3, and TRPV4 sensitivity through a conserved binding site on the ankyrin repeat domainJ Biol Chem2010285173174019864432
- LishkoPVProckoEJinXPhelpsCBGaudetRThe ankyrin repeats of TRPV1 bind multiple ligands and modulate channel sensitivityNeuron200754690591817582331
- NagyISanthaPJancsoGUrbanLThe role of the vanilloid (capsaicin) receptor (TRPV1) in physiology and pathologyEur J Pharmacol20045001–335136915464045
- Planells-CasesRValentePFerrer-MontielAQinFSzallasiAComplex regulation of TRPV1 and related thermo-TRPs: Implications for therapeutic interventionAdv Exp Med Biol201170449151521290313
- SalazarHJara-OsegueraAHernandez-GarciaEStructural determinants of gating in the TRPV1 channelNat Struct Mol Biol200916770471019561608
- ValentePGarcia-SanzNGomisAIdentification of molecular determinants of channel gating in the transient receptor potential box of vanilloid receptor IFASEB J20082293298330918559878
- BrauchiSOrioPVoltage sensing in thermo-TRP channelsAdv Exp Med Biol201170451753021290314
- Garcia-SanzNFernandez-CarvajalAMorenilla-PalaoCIdentification of a tetramerization domain in the C terminus of the vanilloid receptorJ Neurosci200424235307531415190102
- ZhangFLiuSYangFZhengJWangKIdentification of a tetrameric assembly domain in the C-terminus of heat-activated TRPV1 channelsJ Biol Chem201128617153081531621357419
- KwakJWangMHHwangSWKimTYLeeSYOhUIntracellular ATP increases capsaicin-activated channel activity by interacting with nucleotide-binding domainsJ Neurosci200020228298830411069936
- VoetsTNiliusBModulation of TRPs by PIPsJ Physiol2007582Pt 393994417395625
- CortrightDNSzallasiABiochemical pharmacology of the vanilloid receptor TRPV1. An updateEur J Biochem2004271101814181915128291
- LatorreRVargasGOrtaGBrauchiSVoltage and temperature gating in thermoTRP channelsLiedtkeWHellerSTRP Ion Channel Function in Sensory Transduction and Cellular Signaling CascadesLondon, UKCRC Taylor and Francis2007
- RichardsonJDVaskoMRCellular mechanisms of neurogenic inflammationJ Pharmacol Exp Ther2002302383984512183638
- FerrariLFBogenOLevineJDNociceptor subpopulations involved in hyperalgesic primingNeuroscience2010165389690119931357
- ChienLYChengJKChuDChengCFTsaurMLReduced expression of A-type potassium channels in primary sensory neurons induces mechanical hypersensitivityJ Neurosci200727379855986517855600
- CavanaughDJCheslerATJacksonACTRPV1 reporter mice reveal highly restricted brain distribution and functional expression in arteriolar smooth muscle cellsJ Neurosci201131135067507721451044
- MezeyETothZECortrightDNDistribution of mRNA for vanilloid receptor subtype 1 (VR1), and VR1-like immunoreactivity, in the central nervous system of the rat and humanProc Natl Acad Sci U S A20009773655366010725386
- SouthallMDLiTGharibovaLSPeiYNicolGDTraversJBActivation of epidermal vanilloid receptor-1 induces release of proinflammatory mediators in human keratinocytesJ Pharmacol Exp Ther2003304121722212490594
- ChoiTYParkSYJoJYEndogenous expression of TRPV1 channel in cultured human melanocytesJ Dermatol Sci200956212813019656659
- LazzeriMVannucchiMGZardoCImmunohistochemical evidence of vanilloid receptor 1 in normal human urinary bladderEur Urol200446679279815548449
- BirderLAKanaiAJde GroatWCVanilloid receptor expression suggests a sensory role for urinary bladder epithelial cellsProc Natl Acad Sci U S A20019823133961340111606761
- HeinerIEisfeldJHalaszovichCRExpression profile of the transient receptor potential (TRP) family in neutrophil granulocytes: Evidence for currents through long TRP channel 2 induced by ADP-ribose and NADBiochem J2003371Pt 31045105312564954
- StanderSMoormannCSchumacherMExpression of vanilloid receptor subtype 1 in cutaneous sensory nerve fibers, mast cells, and epithelial cells of appendage structuresExp Dermatol200413312913914987252
- ChenCWLeeSTWuWTFuWMHoFMLinWWSignal transduction for inhibition of inducible nitric oxide synthase and cyclooxygenase-2 induction by capsaicin and related analogs in macrophagesBr J Pharmacol200314061077108714530214
- MohapatraDPWangSYWangGKNauCA tyrosine residue in TM6 of the vanilloid receptor TRPV1 involved in desensitization and calcium permeability of capsaicin-activated currentsMol Cell Neurosci200323231432412812762
- AhernGPBrooksIMMiyaresRLWangXBExtracellular cations sensitize and gate capsaicin receptor TRPV1 modulating pain signalingJ Neurosci200525215109511615917451
- TominagaMCaterinaMJMalmbergABThe cloned capsaicin receptor integrates multiple pain-producing stimuliNeuron19982135315439768840
- Garcia-MartinezCMorenilla-PalaoCPlanells-CasesRMerinoJMFerrer-MontielAIdentification of an aspartic residue in the P-loop of the vanilloid receptor that modulates pore propertiesJ Biol Chem200027542325523255810931826
- NiliusBTalaveraKOwsianikGPrenenJDroogmansGVoetsTGating of TRP channels: A voltage connection?J Physiol2005567Pt 1354415878939
- CromerBAMcIntyrePPainful toxins acting at TRPV1Toxicon200851216317318061640
- AlawiKKeebleJThe paradoxical role of the transient receptor potential vanilloid 1 receptor in inflammationPharmacol Ther2010125218119519896501
- LiuBHuiKQinFThermodynamics of heat activation of single capsaicin ion channels VR1Biophys J20038552988300614581201
- GrandlJKimSEUzzellVTemperature-induced opening of TRPV1 ion channel is stabilized by the pore domainNat Neurosci201013670871420414199
- De PetrocellisLChuCJMorielloASKellnerJCWalkerJMDi MarzoVActions of two naturally occurring saturated N-acyldopamines on transient receptor potential vanilloid 1 (TRPV1) channelsBr J Pharmacol2004143225125615289293
- RosenbaumTSimonSATRPV1 receptors and signal transductionLiedtkeWHellerSTRP Ion Channel Function in Sensory Transduction and Cellular Signaling CascadesLondon, UKCRC Taylor and Francis2007
- MovahedPJonssonBABirnirBEndogenous unsaturated C18 N-acylethanolamines are vanilloid receptor (TRPV1) agonistsJ Biol Chem200528046384963850416081411
- JungJHwangSWKwakJCapsaicin binds to the intracellular domain of the capsaicin-activated ion channelJ Neurosci19991925295389880573
- JungJLeeSYHwangSWAgonist recognition sites in the cytosolic tails of vanilloid receptor 1J Biol Chem200227746444484445412228246
- JordtSEJuliusDMolecular basis for species-specific sensitivity to “hot” chili peppersCell2002108342143011853675
- GavvaNRKlionskyLQuYMolecular determinants of vanilloid sensitivity in TRPV1J Biol Chem200427919202832029514996838
- JordtSETominagaMJuliusDAcid potentiation of the capsaicin receptor determined by a key extracellular siteProc Natl Acad Sci U S A200097148134813910859346
- PatilMJJeskeNAAkopianANTransient receptor potential V1 regulates activation and modulation of transient receptor potential A1 by Ca2+Neuroscience201017141109111920884333
- TousovaKVyklickyLSusankovaKBenediktJVlachovaVGadolinium activates and sensitizes the vanilloid receptor TRPV1 through the external protonation sitesMol Cell Neurosci200530220721716099171
- AhernGPWangXMiyaresRLPolyamines are potent ligands for the capsaicin receptor TRPV1J Biol Chem2006281138991899516431906
- KoplasPARosenbergRLOxfordGSThe role of calcium in the desensitization of capsaicin responses in rat dorsal root ganglion neuronsJ Neurosci19971710352535379133377
- DochertyRJYeatsJCBevanSBoddekeHWInhibition of calcineurin inhibits the desensitization of capsaicin-evoked currents in cultured dorsal root ganglion neurones from adult ratsPflugers Arch199643168288378927498
- MoriyamaTHigashiTTogashiKSensitization of TRPV1 by EP1 and IP reveals peripheral nociceptive mechanism of prostaglandinsMol Pain20051315813989
- XueQJongBChenTSchumacherMATranscription of rat TRPV1 utilizes a dual promoter system that is positively regulated by nerve growth factorJ Neurochem2007101121222217217411
- BronRKlesseLJShahKParadaLFWinterJActivation of Ras is necessary and sufficient for upregulation of vanilloid receptor type 1 in sensory neurons by neurotrophic factorsMol Cell Neurosci200322111813212595244
- MohapatraDPNauCDesensitization of capsaicin-activated currents in the vanilloid receptor TRPV1 is decreased by the cyclic AMP-dependent protein kinase pathwayJ Biol Chem200327850500805009014506258
- RohacsTThyagarajanBLukacsVPhospholipase C mediated modulation of TRPV1 channelsMol Neurobiol2008372–315316318528787
- RosenbaumTGordon-ShaagAMunariMGordonSECa2+/calmodulin modulates TRPV1 activation by capsaicinJ Gen Physiol20041231536214699077
- BhaveGGereauRWPosttranslational mechanisms of peripheral sensitizationJ Neurobiol20046118810615362155
- WirknerKHognestadHJahnelRHuchoFIllesPCharacterization of rat transient receptor potential vanilloid 1 receptors lacking the N-glycosylation site N604Neuroreport2005169997100115931076
- JahnelRDregerMGillenCBenderOKurreckJHuchoFBiochemical characterization of the vanilloid receptor 1 expressed in a dorsal root ganglia derived cell lineEur J Biochem2001268215489549611683872
- JinXMorsyNWinstonJPasrichaPJGarrettKAkbaraliHIModulation of TRPV1 by nonreceptor tyrosine kinase, c-Src kinaseAm J Physiol Cell Physiol20042872C55856315084474
- Planells-CasesRGarcia-SanzNMorenilla-PalaoCFerrer-MontielAFunctional aspects and mechanisms of TRPV1 involvement in neurogenic inflammation that leads to thermal hyperalgesiaPflugers Arch2005451115115915909179
- StanchevDBlosaMMiliusDCross-inhibition between native and recombinant TRPV1 and P2X(3) receptorsPain20091431–2263619223122
- LukacsVThyagarajanBVarnaiPBallaABallaTRohacsTDual regulation of TRPV1 by phosphoinositidesJ Neurosci200727267070708017596456
- KimAYTangZLiuQPirt, a phosphoinositide-binding protein, functions as a regulatory subunit of TRPV1Cell2008133347548518455988
- ZhangXLiLMcNaughtonPAProinflammatory mediators modulate the heat-activated ion channel TRPV1 via the scaffolding protein AKAP79/150Neuron200859345046118701070
- GoswamiCDregerMJahnelRBogenOGillenCHuchoFIdentification and characterization of a Ca2+-sensitive interaction of the vanilloid receptor TRPV1 with tubulinJ Neurochem20049151092110315569253
- Morenilla-PalaoCPlanells-CasesRGarcia-SanzNFerrer-MontielARegulated exocytosis contributes to protein kinase C potentiation of vanilloid receptor activityJ Biol Chem200427924256652567215066994
- WangHBedfordFKBrandonNJMossSJOlsenRWGABA(A)-receptor-associated protein links GABA(A) receptors and the cytoskeletonNature1999397671469729892355
- LainezSValentePOntoria-OviedoIGABAA receptor associated protein (GABARAP) modulates TRPV1 expression and channel function and desensitizationFASEB J20102461958197020179142
- SaundersCIFassettRGGeraghtyDPUp-regulation of TRPV1 in mononuclear cells of end-stage kidney disease patients increases susceptibility to N-arachidonoyl-dopamine (NADA)-induced cell deathBiochim Biophys Acta20091792101019102619619644
- DrayAInflammatory mediators of painBr J Anaesth19957521251317577246
- SchafersMSorkinLEffect of cytokines on neuronal excitabilityNeurosci Lett2008437318819318420346
- AbbadieCBhangooSDe KoninckYMalcangioMMelik-ParsadaniantzSWhiteFAChemokines and pain mechanismsBrain Res Rev200960112513419146875
- SchaibleHGvon BanchetGSBoettgerMKThe role of proinflammatory cytokines in the generation and maintenance of joint painAnn N Y Acad Sci20101193606920398009
- RenKDubnerRInteractions between the immune and nervous systems in painNat Med201016111267127620948535
- GoldMSGebhartGFNociceptor sensitization in pain pathogenesisNat Med201016111248125720948530
- D’MelloRDickensonAHSpinal cord mechanisms of painBr J Anaesth2008101181618417503
- HillRGMolecular basis for the perception of painNeuroscientist20017428229211488394
- EnglerAAeschlimannASimmenBRExpression of transient receptor potential vanilloid 1 (TRPV1) in synovial fibroblasts from patients with osteoarthritis and rheumatoid arthritisBiochem Biophys Res Commun2007359488488817560936
- ChoWGValtschanoffJGVanilloid receptor TRPV1-positive sensory afferents in the mouse ankle and knee jointsBrain Res20081219596518538749
- AkbarAYiangouYFacerPWaltersJRAnandPGhoshSIncreased capsaicin receptor TRPV1-expressing sensory fibres in irritable bowel syndrome and their correlation with abdominal painGut200857792392918252749
- Planells-CasesRFerrer-MontielATRP channel traffickingLiedtkeWHellerSTRP Ion Channel Function in Sensory Transduction and Cellular Signaling CascadesLondon, UKCRC Taylor and Francis2007
- PabbidiRMCaoDSPariharAPauzaMEPremkumarLSDirect role of streptozotocin in inducing thermal hyperalgesia by enhanced expression of transient receptor potential vanilloid 1 in sensory neuronsMol Pharmacol2008733995100418089839
- JiRRSamadTAJinSXSchmollRWoolfCJp38 MAPK activation by NGF in primary sensory neurons after inflammation increases TRPV1 levels and maintains heat hyperalgesiaNeuron2002361576812367506
- ZhangXHuangJMcNaughtonPANGF rapidly increases membrane expression of TRPV1 heat-gated ion channelsEMBO J200524244211422316319926
- BonningtonJKMcNaughtonPASignalling pathways involved in the sensitisation of mouse nociceptive neurones by nerve growth factorJ Physiol.2003551Pt 243344612815188
- SteinATUfret-VincentyCAHuaLSantanaLFGordonSEPhosphoinositide 3-kinase binds to TRPV1 and mediates NGF-stimulated TRPV1 trafficking to the plasma membraneJ Gen Physiol2006128550952217074976
- Van BurenJJBhatSRotelloRPauzaMEPremkumarLSSensitization and translocation of TRPV1 by insulin and IGF-IMol Pain200511715857517
- LiljaJLaulundFForsbyAInsulin and insulin-like growth factor type-I up-regulate the vanilloid receptor-1 (TRPV1) in stably TRPV1-expressing SH-SY5Y neuroblastoma cellsJ Neurosci Res20078571413141917385724
- Camprubi-RoblesMPlanells-CasesRFerrer-MontielADifferential contribution of SNARE-dependent exocytosis to inflammatory potentiation of TRPV1 in nociceptorsFASEB J200923113722373319584302
- MillerRJJungHBhangooSKWhiteFACytokine and chemokine regulation of sensory neuron functionHandb Exp Pharmacol200919441744919655114
- HensellekSBrellPSchaibleHGBrauerRSegond vonBGThe cytokine TNFalpha increases the proportion of DRG neurones expressing the TRPV1 receptor via the TNFR1 receptor and ERK activationMol Cell Neurosci200736338139117851089
- KhanAADiogenesAJeskeNAHenryMAAkopianAHargreavesKMTumor necrosis factor alpha enhances the sensitivity of rat trigeminal neurons to capsaicinNeuroscience2008155250350918582539
- HagenackerTCzeschikJCSchafersMBusselbergDSensitization of voltage activated calcium channel currents for capsaicin in nociceptive neurons by tumor-necrosis-factor-alphaBrain Res Bull201081115716319818386
- HuYGuQLinRLKryscioRLeeLYCalcium transient evoked by TRPV1 activators is enhanced by tumor necrosis factor-alpha in rat pulmonary sensory neuronsAm J Physiol Lung Cell Mol Physiol20102994L48349220639352
- ConstantinCEMairNSailerCAEndogenous tumor necrosis factor alpha (TNFalpha) requires TNF receptor type 2 to generate heat hyperalgesia in a mouse cancer modelJ Neurosci200828195072508118463260
- RussellFAFernandesESCouradeJPKeebleJEBrainSDTumour necrosis factor alpha mediates transient receptor potential vanilloid 1-dependent bilateral thermal hyperalgesia with distinct peripheral roles of interleukin-1beta, protein kinase C and cyclooxygenase- 2 signallingPain2009142326427419231080
- ChengJKJiRRIntracellular signaling in primary sensory neurons and persistent painNeurochem Res200833101970197818427980
- GarrisonSLStuckyCLThe dynamic TRPA1 channel: A suitable pharmacological pain targetCurr Pharm Biotechnol April 52011 [Epub ahead of print]
- KobayashiKFukuokaTObataKDistinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with adelta/c-fibers and colocalization with trk receptorsJ Comp Neurol2005493459660616304633
- TrevisaniMSiemensJMaterazziS4-Hydroxynonenal, an endogenous aldehyde, causes pain and neurogenic inflammation through activation of the irritant receptor TRPA1Proc Natl Acad Sci U S A200710433135191352417684094
- ObataKKatsuraHMizushimaTTRPA1 induced in sensory neurons contributes to cold hyperalgesia after inflammation and nerve injuryJ Clin Invest200511592393240116110328
- ObrejaORatheePKLipsKSDistlerCKressMIL-1 beta potentiates heat-activated currents in rat sensory neurons: Involvement of IL-1RI, tyrosine kinase, and protein kinase CFASEB J200216121497150312374772
- ObrejaOBiasioWAndratschMFast modulation of heat-activated ionic current by proinflammatory interleukin 6 in rat sensory neuronsBrain2005128Pt 71634164115817518
- AndratschMMairNConstantinCEA key role for gp130 expressed on peripheral sensory nerves in pathological painJ Neurosci20092943134731348319864560
- CesarePMcNaughtonPA novel heat-activated current in nociceptive neurons and its sensitization by bradykininProc Natl Acad Sci U S A1996932615435154398986829
- MizumuraKKodaHKumazawaTEvidence that protein kinase C activation is involved in the excitatory and facilitatory effects of bradykinin on canine visceral nociceptors in vitroNeurosci Lett1997237129329406872
- MizumuraKSugiuraTKatanosakaKBanikRKKozakiYExcitation and sensitization of nociceptors by bradykinin: What do we know?Exp Brain Res20091961536519396590
- TominagaMWadaMMasuMPotentiation of capsaicin receptor activity by metabotropic ATP receptors as a possible mechanism for ATP-evoked pain and hyperalgesiaProc Natl Acad Sci U S A200198126951695611371611
- KajiharaYMurakamiMImagawaTOtsuguroKItoSOhtaTHistamine potentiates acid-induced responses mediating transient receptor potential V1 in mouse primary sensory neuronsNeuroscience2010166129230420006972
- SchnizlerKShutovLPVan KaneganMJProtein kinase A anchoring via AKAP150 is essential for TRPV1 modulation by forskolin and prostaglandin E2 in mouse sensory neuronsJ Neurosci200828194904491718463244
- ZhangNInanSCowanAA proinflammatory chemokine, CCL3, sensitizes the heat- and capsaicin-gated ion channel TRPV1Proc Natl Acad Sci U S A2005102124536454115764707
- JungHTothPTWhiteFAMillerRJMonocyte chemoattractant protein-1 functions as a neuromodulator in dorsal root ganglia neuronsJ Neurochem2008104125426317944871
- HolzerPLocal effector functions of capsaicin-sensitive sensory nerve endings: involvement of tachykinins, calcitonin gene-related peptide and other neuropeptidesNeuroscience19882437397683288903
- JancsóGNeuroImmune biology Neruogenic Inflammation in Health and DiseaseAmsterdam, The NetherlandsElsevier2009
- GeppettiPHolzerPNeurogenic InflammationBoca Raton, FLCRC Press1996
- MaggiCATachykinins and calcitonin gene-related peptide (CGRP) as co-transmitters released from peripheral endings of sensory nervesProg Neurobiol19954511987716258
- MaggiCAMeliAThe sensory-efferent function of capsaicin-sensitive sensory neuronsGen Pharmacol19881911433278943
- KichkoTIReehPWTRPV1 controls acid- and heat-induced calcitonin gene-related peptide release and sensitization by bradykinin in the isolated mouse tracheaEur J Neurosci20092991896190419473241
- PriceTJLouriaMDCandelario-SotoDTreatment of trigeminal ganglion neurons in vitro with NGF, GDNF or BDNF: Effects on neuronal survival, neurochemical properties and TRPV1-mediated neuropeptide secretionBMC Neurosci20056415667652
- ThanMNemethJSzilvassyZPinterEHelyesZSzolcsanyiJSystemic anti-inflammatory effect of somatostatin released from capsaicin-sensitive vagal and sciatic sensory fibres of the rat and guinea-pigEur J Pharmacol20003992–325125810884527
- HelyesZSzaboANemethJAntiinflammatory and analgesic effects of somatostatin released from capsaicin-sensitive sensory nerve terminals in a Freund’s adjuvant-induced chronic arthritis model in the ratArthritis Rheum20045051677168515146439
- AdcockJJTRPV1 receptors in sensitisation of cough and pain reflexesPulm Pharmacol Ther2009222657019141328
- FernandesESRussellFASpinaDA distinct role for TRPA1, in addition to TRPV1, in TNFalpha-induced inflammatory hyperalgesia and CFA-induced mono-arthritisArthritis Rheum201163381982921360511
- KeebleJRussellFCurtisBStarrAPinterEBrainSDInvolvement of transient receptor potential vanilloid 1 in the vascular and hyperalgesic components of joint inflammationArthritis Rheum200552103248325616200599
- GhilardiJRRohrichHLindsayTHSelective blockade of the capsaicin receptor TRPV1 attenuates bone cancer painJ Neurosci200525123126313115788769
- GoblirschMJZwolakPClohisyDRAdvances in understanding bone cancer painJ Cell Biochem200596468268816149079
- NagaeMHiragaTYonedaTAcidic microenvironment created by osteoclasts causes bone pain associated with tumor colonizationJ Bone Miner Metab20072529910417323179
- MachDBRogersSDSabinoMCOrigins of skeletal pain: Sensory and sympathetic innervation of the mouse femurNeuroscience2002113115516612123694
- PetersCMGhilardiJRKeyserCPTumor-induced injury of primary afferent sensory nerve fibers in bone cancer painExp Neurol200519318510015817267
- ShinodaMOginoAOzakiNInvolvement of TRPV1 in nociceptive behavior in a rat model of cancer painJ Pain20089868769918455478
- BuddenkotteJSteinhoffMPathophysiology and therapy of pruritus in allergic and atopic diseasesAllergy201065780582120384615
- ShimWSOhUHistamine-induced itch and its relationship with painMol Pain200842918667087
- ImamachiNParkGHLeeHTRPV1-expressing primary afferents generate behavioral responses to pruritogens via multiple mechanismsProc Natl Acad Sci U S A200910627113301133519564617
- DendaMTsutsumiMRoles of transient receptor potential proteins (TRPs) in epidermal keratinocytesAdv Exp Med Biol201170484786021290330
- ShimWSTakMHLeeMHTRPV1 mediates histamine-induced itching via the activation of phospholipase A2 and 12-lipoxygenaseJ Neurosci20072792331233717329430
- BolliRAbdel-LatifANo pain, no gain: The useful function of anginaCirculation2005112233541354316330693
- HaradaNOkajimaKYukselMIsobeHContribution of capsaicin-sensitive sensory neurons to antithrombin-induced reduction of ischemia/reperfusion-induced liver injury in ratsThromb Haemost2005931485615630490
- MizutaniAOkajimaKMurakamiKActivation of sensory neurons reduces ischemia/reperfusion-induced acute renal injury in ratsAnesthesiology2009110236136919194162
- BanvolgyiAPalinkasLBerkiTEvidence for a novel protective role of the vanilloid TRPV1 receptor in a cutaneous contact allergic dermatitis modelJ Neuroimmunol20051691–2869616188326
- SibaevAMassaFYuceBCB1 and TRPV1 receptors mediate protective effects on colonic electrophysiological properties in miceJ Mol Med200684651352016501934
- WardSMBayguinovJWonKJGrundyDBerthoudHRDistribution of the vanilloid receptor (VR1) in the gastrointestinal tractJ Comp Neurol2003465112113512926020
- MassaFSibaevAMarsicanoGBlaudzunHStorrMLutzBVanilloid receptor (TRPV1)-deficient mice show increased susceptibility to dinitrobenzene sulfonic acid induced colitisJ Mol Med200684214214616389550
- AbrahamEMatthayMADinarelloCConsensus conference definitions for sepsis, septic shock, acute lung injury, and acute respiratory distress syndrome: Time for a reevaluationCrit Care Med200028123223510667529
- VincentJLMartinezEOSilvaEEvolving concepts in sepsis definitionsCrit Care Clin2009254665675vii19892246
- UlloaLTraceyKJThe “cytokine profile”: A code for sepsisTrends Mol Med2005112566315694867
- SriskandanSAltmannDMThe immunology of sepsisJ Pathol2008214221122318161754
- UlloaLBrunnerMRamosLDeitchEAScientific and clinical challenges in sepsisCurr Pharm Des200915161918193519519432
- ThornberryNACaspases: Key mediators of apoptosisChem Biol199855R971039578633
- HotchkissRSNicholsonDWApoptosis and caspases regulate death and inflammation in sepsisNat Rev Immunol200661181382217039247
- WescheDELomas-NeiraJLPerlMChungCSAyalaALeukocyte apoptosis and its significance in sepsis and shockJ Leukoc Biol200578232533715817707
- ShimaokaMParkEJAdvances in understanding sepsisEur J Anaesthesiol Suppl20084214615318289433
- BrogdenKAGuthmillerJMSalzetMZasloffMThe nervous system and innate immunity: The neuropeptide connectionNat Immunol20056655856415908937
- JoyceCDFiscusRRWangXDriesDJMorrisRCPrinzRACalcitonin gene-related peptide levels are elevated in patients with sepsisSurgery19901086109711012247835
- ArnalichFSanchezJFMartinezMChanges in plasma concentrations of vasoactive neuropeptides in patients with sepsis and septic shockLife Sci199556275817529863
- BeerSWeighardtHEmmanuilidisKSystemic neuropeptide levels as predictive indicators for lethal outcome in patients with postoperative sepsisCrit Care Med20023081794179812163795
- ForemanJCJordanCCOehmePRennerHStructure-activity relationships for some substance P-related peptides that cause wheal and flare reactions in human skinJ Physiol19833354494656192237
- BrainSDNewboldPKajekarRModulation of the release and activity of neuropeptides in the microcirculationCan J Physiol Pharmacol19957379959988846443
- HarzenetterMDNovotnyARGaisPMolinaCAAltmayrFHolzmannBNegative regulation of TLR responses by the neuropeptide CGRP is mediated by the transcriptional repressor ICERJ Immunol2007179160761517579082
- NumaoTAgrawalDKNeuropeptides modulate human eosinophil chemotaxisJ Immunol199214910330933151385521
- LeviteMCahalonLHershkovizRSteinmanLLiderONeuropeptides, via specific receptors, regulate T cell adhesion to fibronectinJ Immunol1998160299310009551939
- FoxFEKubinMCassinMCalcitonin gene-related peptide inhibits proliferation and antigen presentation by human peripheral blood mononuclear cells: Effects on B7, interleukin 10, and interleukin 12J Invest Dermatol1997108143488980285
- CrossmanDMcEwanJMacDermotJMacIntyreIDolleryCTHuman calcitonin gene-related peptide activates adenylate cyclase and releases prostacyclin from human umbilical vein endothelial cellsBr J Pharmacol19879246957013322463
- OkajimaKHaradaNRegulation of inflammatory responses by sensory neurons: Molecular mechanism(s) and possible therapeutic applicationsCurr Med Chem200613192241225116918352
- GomesRNCastro-Faria-NetoHCBozzaPTCalcitonin gene-related peptide inhibits local acute inflammation and protects mice against lethal endotoxemiaShock200524659059416317392
- ReinshagenMFlamigGErnstSCalcitonin gene-related peptide mediates the protective effect of sensory nerves in a model of colonic injuryJ Pharmacol Exp Ther199828626576619694917
- SutoBBagolyTBorzseiRSurgery and sepsis increase somatostatin-like immunoreactivity in the human plasmaPeptides20103161208121220307604
- BergRMStraussGIToftengFCirculating levels of vasoactive peptides in patients with acute bacterial meningitisIntensive Care Med20093591604160819513693
- BryantPShumateMYumetGLangCHVaryTCCooneyRNCapsaicin-sensitive nerves regulate the metabolic response to abdominal sepsisJ Surg Res2003112215216112888332
- DemirbilekSErsoyMODemirbilekSSmall-dose capsaicin reduces systemic inflammatory responses in septic ratsAnesth Analg20049951501150715502055
- KimCSKawadaTKimBSCapsaicin exhibits anti-inflammatory property by inhibiting IkB-a degradation in LPS-stimulated peritoneal macrophagesCell Signal200315329930612531428
- WangYNovotnyMQuaiserova-MockoVSwainGMWangDHTRPV1-mediated protection against endotoxin-induced hypotension and mortality in ratsAm J Physiol Regul Integr Comp Physiol20082945R1517152318337316
- AngSFMoochhalaSMBhatiaMHydrogen sulfide promotes transient receptor potential vanilloid 1-mediated neurogenic inflammation in polymicrobial sepsisCrit Care Med201038261962819851090
- De WinterBYBredenoordAJVanNLInvolvement of afferent neurons in the pathogenesis of endotoxin-induced ileus in mice: Role of CGRP and TRPV1 receptorsEur J Pharmacol20096151–317718419445917
- MuraiMTsujiFNoseMSA13353 (1-[2-(1-Adamantyl)ethyl]-1-pentyl-3-[3-(4-pyridyl)propyl]urea) inhibits TNF-alpha production through the activation of capsaicin-sensitive afferent neurons mediated via transient receptor potential vanilloid 1 in vivoEur J Pharmacol20085882–330931518508045
- TsujiFMuraiMOkiKEffects of SA13353, a transient receptor potential vanilloid 1 agonist, on leukocyte infiltration in lipopolysaccharide- induced acute lung injury and ovalbumin-induced allergic airway inflammationJ Pharmacol Sci2010112448749020351486
- ClarkNKeebleJFernandesESThe transient receptor potential vanilloid 1 (TRPV1) receptor protects against the onset of sepsis after endotoxinFASEB J200721133747375517601984
- GuptillVCuiXKhaibullinaADisruption of the transient receptor potential vanilloid 1 can affect survival, bacterial clearance, and cytokine gene expression during murine sepsisAnesthesiology March 42011 [Epub ahead of print]
- HelyesZPozsgaiGBorzseiRInhibitory effect of PACAP-38 on acute neurogenic and non-neurogenic inflammatory processes in the ratPeptides20072891847185517698245
- OrliacMLPeroniRNAbramoffTNeumanIPodestaEJAdler-GraschinskyEIncreases in vanilloid TRPV1 receptor protein and CGRP content during endotoxemia in ratsEur J Pharmacol20075661–314515217482593
- HuttemeierPCRitterEFBenvenisteHCalcitonin gene-related peptide mediates hypotension and tachycardia in endotoxic ratsAm J Physiol19932652 Pt 2H7677698368378
- IidaTShimizuINealenMLCampbellACaterinaMAttenuated fever response in mice lacking TRPV1Neurosci Lett20053781283315763167
- DoganMDPatelSRudayaAYSteinerAASzekelyMRomanovskyAALipopolysaccharide fever is initiated via a capsaicin-sensitive mechanism independent of the subtype-1 vanilloid receptorBr J Pharmacol200414381023103215492017