
Abstract
The COVID-19 pandemic has caused devastating loss of life and a healthcare crisis worldwide. SARS-CoV-2 is the causative pathogen of COVID-19 and is transmitted mainly through the respiratory tract, where the virus infects host cells by binding to the ACE2 receptor. SARS-CoV-2 infection is associated with acute pneumonia, but neuropsychiatric symptoms and different brain injuries are also present. The possible routes by which SARS-CoV-2 invades the brain are unclear, as are the mechanisms underlying brain injuries with the resultant neuropsychiatric symptoms in patients with COVID-19. Ferroptosis is a unique iron-dependent form of non-apoptotic cell death, characterized by lipid peroxidation with high levels of glutathione consumption. Ferroptosis plays a primary role in various acute and chronic brain diseases, but to date, ferroptosis in COVID-19-related brain injuries has not been explored. This review discusses the mechanisms of ferroptosis and recent evidence suggesting a potential pathogenic role for ferroptosis in COVID-19-related brain injury. Furthermore, the possible routes through which SARS-CoV-2 could invade the brain are also discussed. Discoveries in these areas will open possibilities for treatment strategies to prevent or reduce brain-related complications of COVID-19.
Introduction
Coronavirus disease 2019 (COVID-19), triggered by the coronavirus-2 virus of the severe acute respiratory syndrome (SARS-CoV-2), is a critical health concern worldwide.Citation1 By January 2022, over 349 million cases of COVID-19 and over 5.5 million people have died worldwide.Citation2 Central nervous system (CNS) involvement is one possible cause of death among patients with COVID-19.Citation3 Analysis of clinical data for 214 patients with COVID-19 indicated that 36% had CNS manifestations, including acute cerebrovascular disease with disturbances in consciousness.Citation4 Furthermore, SARS-CoV-2 infection has been shown to affect the CNS and cause a series of neuropsychiatric symptoms (eg, anosmia, headache, cognitive and attention deficits, disturbances in consciousness, anxiety, depression, delirium, and suicidal behavior) and diseases (eg, acute necrotizing encephalopathy, encephalitis, hypoxic brain injury, strokes, myelitis, tonic epilepsy, neurogenic respiratory failure, generalized myoclonus, and Kawasaki syndrome).Citation5–10 This wide range of symptoms and diseases suggests that various underlying mechanisms cause and promote brain injury in patients with COVID-19. Therefore, exploring possible underlying mechanisms can help guide the development of treatment for brain injuries with subsequent neuropsychiatric symptoms in patients with COVID-19.
Among these mechanisms, ferroptosis may contribute to the pathogenesis of COVID-19-related brain injuries. First described in 2012, Dixon et alCitation11 explained that ferroptosis is an iron-dependent programmed nonapoptotic cell death with unique biological processes and pathophysiologic characteristics. The pathogenic processes of ferroptosis involve an excessive iron metabolism that produces iron-dependent oxidative stress and causes damage to nucleic acids, proteins, and lipids that eventually induce cell death.Citation12,Citation13 Ferroptosis has recently gained considerable attention in brain research and has significant implications for several neurologic diseases, such as ischemic stroke, intracerebral hemorrhage (ICH), Alzheimer’s disease, and Parkinson’s disease.Citation12–16 In addition, emerging evidence indicates that ferroptosis is a nexus between metabolism, redox biology, and human diseases such as COVID-19.Citation17 However, the specific underlying mechanisms of ferroptosis remain unclear. Exploring the role of ferroptosis in COVID-19 infection and COVID-19-related brain injury, in particular, could be valuable for identifying therapeutic targets to prevent or lessen brain complications of COVID-19 and improve prognosis. In this regard, brain-permeable ferroptosis inhibitors or iron-chelating agents could be tested to assess whether they affect neuropsychiatric symptoms caused by COVID-19-related brain injuries.Citation17
Brain Injuries Caused by COVID-19
Neuropsychiatric Symptoms and Brain Injuries in Some Patients with COVID-19
COVID-19, mainly described as acute pneumonia, is increasingly recognized as a disease involving multiple organ dysfunction, including brain microbleeds.Citation18 All the results of different groups indicated the presence of neuropsychiatric symptoms in some patients infected with SARS-CoV-2 (). Two studies conducted in Germany and the United Kingdom showed that post-COVID neuropsychiatric symptoms were observed in 20% to 70% of patients, respectively. These symptoms lasted months after the resolution of respiratory symptoms.Citation8 A systematic review concluded that the average prevalence of headaches among COVID-19 patients was 8%.Citation19 An analysis of the clinical characteristics of 138 hospitalized patients with COVID-19 showed that the incidence of dizziness was between 7% and 9.4%.Citation20 A study of 31 patients with COVID-19 showed that 45% experienced anosmia, 29% had hyposmia, and 6% had dysosmia,Citation21 indicating that COVID-19 can damage the olfactory bulb. A review of the neurologic manifestations of COVID-19 showed that an average of 25% of the patients had CNS dysfunction, 7.5% had disturbances in consciousness, 3% had acute cerebrovascular disease, and 0.5% had ataxia.Citation22
Table 1 Studies Reporting Neuropsychiatric Diseases and Symptoms Related to COVID-19
Furthermore, COVID-19 may cause cognitive impairment, such as confusion, inattention, anxiety, and disorientation.Citation23,Citation24 Neuropsychological evaluation of 57 patients with COVID-19 who had recovered showed that 81% of the cases had cognitive impairment ranging from mild to severe.Citation25 In addition, there have been case reports of seizures in some patients with COVID-19. The severity of the disease was affected by systemic inflammation, which differs from ‘common seizures’ that have specific cortical, thalamic, or posterior fossa involvement.Citation26 Together, these neuropsychiatric symptoms may be related to COVID-19-related brain damage to particular regions of the brain.
Brain injuries (eg, cerebral vascular pathology, arteriosclerosis, ischemic strokes, and ICH have been discovered in COVID-19. A magnetic resonance imaging study of 13 patients with COVID-19 showed that 11 (85%) had bilateral frontotemporal hypoperfusion, 3 (23%) had acute or subacute strokes, and 8 (62%) had enlargement of the papillary space.Citation24 Another study with high-resolution magnetic resonance imaging of 13 patients who died of COVID-19 showed abnormalities in the brains of 10 patients (76.9%).Citation27 The incidence of brain injury in critically ill and dead patients is higher than in noncritically ill patients.Citation22 A systematic review of patients with COVID-19 indicated that 35.6% had inflammation and activation of astrocytes and microglia in the brain, and 28.1% had hypoxic-ischemic injury.Citation28 Of these patients, 29.5% had arteriosclerosis, 12.4% had ICH, and 2.7% had infarcts in the cerebral cortex and subcortex.Citation28 COVID-19 is now an independent risk factor for acute stroke.Citation29 As an important but under-recognized complication of COVID-19,Citation30 acute stroke affects approximately 1–3% of hospitalized patients and 6% of ICU patients; male patients with COVID-19 have a higher incidence of strokes (62%) compared to female patients.Citation31 In a retrospective study of 219 hospitalized COVID-19 patients in China, 11 patients (5%) had an acute stroke (10 cases of ischemic stroke, 1 case of hemorrhagic stroke).Citation32 All of the above data indicate the presence of different brain injuries among COVID-19 patients.
SARS-CoV-2 Invasion with Subsequent Brain Injuries
Like SARS-CoV and the Middle East Respiratory Syndrome Coronavirus,Citation33 SARS-CoV-2 can also invade and spread throughout the brain.Citation34,Citation35 Increasing numbers of studies provide direct evidence for the neuroinvasiveness of SARS-CoV-2 (). However, the mechanism by which the virus infects the specific regions of the brain has not been fully elucidated. In this review, we speculated and concluded that SARS-CoV-2 infection could cause brain damage through three routes (). The SARS-CoV-2 viral particles can invade the brain through the vascular pathway once the blood-brain barrier (BBB) is broken,Citation36 or through peripheral nerves such as the vagus nerve and reverse transneuronal transport of axonal transport,Citation34,Citation37 or through the cerebral spinal flow (CSF) pathway.Citation34
Table 2 Evidence Supporting the Invasion of SARS-CoV-2 into the Central Nervous System
Figure 1 SARS-CoV-2 infection can cause brain damage through three possible routes. COVID-19 infection causes various brain injuries and diseases (eg, hypoxic brain injury, encephalitis, hemorrhagic necrotizing encephalopathy, acute disseminated encephalomyelitis, and acute stroke). SARS-CoV-2 invades the brain likely through three routes: (1) the vascular pathway where SARS-CoV-2 damages the blood-brain barrier to invade the brain via blood or lymphatic circulation; (2) the peripheral nerve pathway in which SARS-CoV-2 infects the olfactory bulb, the trigeminal nerve, the vagus nerve, and finally the brain through trans-neuronal or retrograde axonal transport; and (3) the cerebrospinal fluid pathway, SARS-CoV-2 can enter the brain through circulating cerebrospinal fluid to trigger a series of proinflammatory and immune responses that eventually result in various brain injuries with neuropsychiatric symptoms.
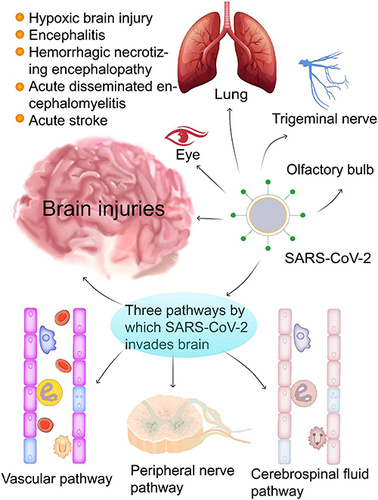
SARS-CoV-2 can infect the brain through compromised vasculature. Cytokine storms can severely damage the BBB, allowing viral particles from SARS-CoV-2 to circulate in the bloodstream to enter the brain.Citation34 The serum level of S100B, an astrocyte marker, increased when symptoms of the CNS appearedCitation36 and reflected destruction of the BBB,Citation38 which is conducive to invasion by SARS-CoV-2 and opens a vasculature path for the virus with inflammatory cytokines. Neuroinvasion of the virus can cause severe brain edema and seizures due to a cerebrovascular event resulting from the hypercoagulable state of blood in COVID-19 patients.Citation39 Cerebral endothelial cells damaged by the SARS-CoV-2 virus produce thrombin, a key coagulation factor that leads to the formation of microthrombi within blood vessels.Citation40 Changes in blood components, such as increased thrombin and D-dimer levels and thrombocytopenia caused by COVID-19, can promote mini-strokes or microbleeds.Citation41 Neuropsychiatric symptoms of COVID-19 may develop due to damage to brain tissue caused by these mini-strokes or microbleeds.Citation41
SARS-CoV-2 can enter the brain by crossing the neural-mucosal interface in the olfactory mucosa, conjunctiva, or taste buds and invades the brain by retrograde axonal transport.Citation42 The invasion of SARS-CoV-2 through peripheral nerve terminals (eg, olfactory nerve, trigeminal nerve, glossopharyngeal nerve, or vagus nerve) could be a reasonable route to brain infection,Citation34,Citation35 leading to hyposmia, hypoplasia, and hypogeusia in patients with COVID-19.Citation43 Anosmia is a unique symptom of COVID-19Citation21 and may represent a neural invasion of the virus through the olfactory bulb during the early or chronic stages.Citation43,Citation44 SARS-CoV-2 may directly infect olfactory sensory neurons through the nerve-mucosal interface in the olfactory epithelium and then enter the CNS through the olfactory nerve.Citation42 The SARS-CoV-2 virus spreads primarily through respiratory droplets that enter the respiratory tract. However, a small fraction of viruses remain in the nose, conjunctiva, and oral cavity, rich in angiotensin-converting enzyme-2 (ACE2).Citation34,Citation45 One report indicated the presence of SARS-CoV-2 RNA in conjunctival swabs and tear samples from some patients with COVID-19.Citation45 Conjunctival contact with droplets containing SARS-CoV-2 can result in the retrograde neuronal transmission of the virus to infect the brain and impair vision.Citation46 In addition, SARS-CoV-2 can enter the brain through the glossopharyngeal nerve or the vagus nerve that connects the nucleus of the solitary tract through the sensory neurons of the tongue.Citation46 Once the peripheral nerve, the nucleus of the solitary tract, the thalamus, or other pathways related to taste conduction are invaded and damaged by SARS-CoV-2, hypogeusia occurs in patients.Citation43 Furthermore, the solitary tract nucleus is located in the brainstem, close to the respiratory and cardiovascular control centers.Citation42 After SARS-CoV-2 infects the solitary nucleus along the nerve, it can infect this nucleus, resulting in refractory dyspnea.Citation47
Although the frequency of SARS-CoV-2 RNA in the CSF of COVID-19 patients is low (~1.3%),Citation3 it cannot exclude the CSF as a potential route for the invasion of SARS-CoV-2 into the brain.Citation48 The brain has its lymphatic drainage system, in which the CSF plays a primary role. A new study showed that SARS-CoV-2 could invade peripheral lymphatic vessels connected to the brain’s lymphatic system.Citation49 SARS-CoV-2 can invade the CSF and damage the brain through this anatomical connection. Furthermore, when the BBB is damaged, SARS-CoV-2 overflows from blood vessels and invades brain tissue and the CSF, resulting in brain edema and dysfunction.Citation34 SARS-CoV-2 antibodies have been detected in the CSF of patients with COVID-19 encephalopathy.Citation50 These antibodies may stimulate glial cells leading to neuroinflammation and other cytokine storm and oxidative stress that cause immune-mediated neurological damage.Citation51
Furthermore, infiltration of the SARS-CoV-2 virus into the brain stem or limbic system can result in dysfunction of the autonomic nervous system and emotional dysfunction, respectively, in some patients with COVID-19.Citation8 In short, the COVID-19 virus can invade the brain via the routes above, resulting in brain damage with neuropsychiatric symptoms. These specific neuropsychiatric symptoms can vary in patients according to the location these pathologic events occur in the brain.
After invasion into the brain, SARS-CoV-2 invades brain cells through the ACE2 receptor, which is an identified receptor for SARS-CoV-2 invasion.Citation52 ACE2 is present mainly in mucosal epithelial cells of the lung and arteries but is also highly expressed in brain cells such as neurons, oligodendrocytes, and astrocytes.Citation53 The serine 4 transmembrane protease protein, which facilitates the fusion of SARS-CoV-2 and cellular membranes, is detectable in the cerebral cortex, caudate nucleus, and hippocampus and determines virus tropism.Citation37 Animal studies showed that SARS-CoV-2 damages the cerebral vascular endothelium and infects the brain parenchyma; ACE2 was a key mediator of neuronal damage.Citation54 Furthermore, the SARS-CoV-2 virus spike binds to ACE2 receptors to generate excessive cytokines and promote the formation of blood clots,Citation55 leading to cell apoptosis, necrosis, ferroptosis, and eventually brain tissue damage.Citation56
Infection with SARS-CoV-2 can cause cytokine storms,Citation57 and large amounts of inflammatory factors produced during a cytokine storm can damage brain tissues.Citation36 High levels of tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), and interleukin-1β (IL-1β) induce a neuroinflammatory response in the brain.Citation58,Citation59 Besides, COVID-19 infection activates various inflammatory pathways, such as that for tryptophan-kynurenine metabolism, which is an immunoregulatory pathway.Citation60 2,3-dioxygenase-1 (IDO1) is located in the critical branch of kynurenine metabolism and is activated by inflammatory factors such as TNF-α and IL-6.Citation61 Activating IDO1 in COVID-19 can convert tryptophan to kynurenine and deprive T lymphocytes of tryptophan to inhibit T lymphocyte activation, thus limiting the host immune response.Citation62 Furthermore, the IDO1-kynurenine-aryl hydrocarbon receptor pathway is activated, leading to the lethal consequences of SARS-CoV-2 infection and immune evasion.Citation63 IDO1 activation increases kynurenine metabolism in COVID-19, and kynurenine produces large amounts of chemokines and neurotoxic substances that cause neuronal damage and long-term brain impairment.Citation61 In addition, enhanced inflammation decreases the synthesis of monoamines and trophic factors, thus inhibiting neurotransmitters and neuronal growth.Citation64 Therefore, cerebral inflammation and hypoxic brain injury result in short- and long-term neuropsychiatric symptoms.
Ferroptosis and Its Underlying Mechanism
Ferroptosis is a programmed non-apoptotic cell death associated with an increase in the liable ferrous iron pool (LIP) caused by iron overload. The accumulation of lethal levels of iron-dependent lipid peroxides and reactive oxygen species (ROS) due to the inactivation of glutathione peroxidase 4 (GPX4) and the depletion of glutathione (GSH) are the core mechanisms of ferroptosis.Citation65–67 At the molecular level, ferroptosis is characterized by changes in GSH, GPX4, and ROS levels and the expression of genes that regulate iron homeostasis and lipid peroxidation.Citation68 Changes in intracellular organelle microstructure are also a feature of ferroptosis. For example, during ferroptosis, mitochondria condense and become small, and the lipid bilayer membrane thickens and contracts with the outer membrane rupture. These features can be visualized by deep staining. Additionally, decreased or absent mitochondrial cristae are accompanied by a reduction in cell size and cell connections that lead to cell separation, although the nuclear and cell membranes remain intact.Citation69 F4-hydroxynonenal, cytotoxic malondialdehyde, and other products of lipid peroxide degradation are the primary biomarkers of ferroptosis.Citation70
Ferroptosis is closely related to several pathologic processes, including inflammation, T-cell immunity, acute renal failure, blood disorders, ischemia/reperfusion injury, and CNS diseases.Citation65,Citation71,Citation72 Key regulatory targets of ferroptosis include system Xc- activity, intracellular LIP, GPX4 activity, GSH production, lipid ROS production, and phosphatidylethanolamine (PE) biosynthesis, detailed below.
(1) System Xc- activity regulation: The cell membrane system Xc-, a cystine/glutamate antitransporter comprising heterodimers SLC3A2 and SLC7A11, is involved in GSH synthesis to protect cells from oxidative damage. Beclin 1 regulates antioxidant capacity by directly blocking the activity of the Xc-system by binding to its core component SLC7A11 to promote ferroptosis.Citation73 Furthermore, inhibition of Beclin 1 expression decreases lipid peroxidation, thus mitigating brain edema and neurologic deficits after subarachnoid hemorrhage, probably due to increased system Xc- activity.Citation74 Therefore, the system Xc- could be a therapeutic target for treating COVID-19-related brain injury.
(2) GPX4-GSH regulation: As a central regulator of ferroptosis, GPX4 inhibits the formation of toxic lipid ROS by converting lipid hydroperoxides to non-toxic lipid alcohols.Citation75,Citation76 The availability of NADPH, a cell reducing agent, is critical since it transforms GSH from an oxidized state to a reduced form. If GPX4 cannot effectively degrade ROS, phospholipid hydroperoxides accumulate that can induce ferroptosis in the presence of iron.Citation77 Importantly, GPX4 overexpression can protect cells against ferroptosis.Citation78 Furthermore, selenium (Se) is a critical modulator of GPX4 activity,Citation79 and pharmacological supplementation with Se can enhance GPX4 activity by upregulating its transcription through coactivation of the transcription factor activating protein 2 gamma and the specificity protein 1 that effectively protects neurons against GPX4-dependent ferroptosis.Citation79 Furthermore, activation of the nuclear factor E2-related factor 2 (Nrf2)/Kelch-like ECH-associated protein 1(KEAP-1)-GPX4 pathway also exhibits excellent antiferroptosis effects by inhibiting lipid peroxidation by upregulation of GPX4 and SLC7A11 transcription and increases in NADPH and GSH levels.Citation80
(3) Lipid ROS regulation: Inactivation of SLC7A11 and GSH depletion leads to ROS accumulation. Hydroxyl radicals formed by the Fenton reaction can react with polyunsaturated fatty acids (PUFAs) in lipid membranes in the presence of reduced iron, leading to lipid peroxidation.Citation81 ROS production and detoxification imbalance leads to oxidative stress and subsequent free radical-mediated damage to DNA, proteins, and lipids.Citation82 In this regard, lipid antioxidants (for example, vitamin E, ferrostatin-1 (Fer-1), and liprostatin-1) can decrease intracellular lipid peroxidation and inhibit ferroptosis,Citation83,Citation84 thus showing neuroprotection after acute brain injuries (for example, ICH and traumatic brain injury).Citation85–90
(4) Iron regulation: Iron exists in the forms of Fe2+ or Fe3+ and maintains a dynamic balance in vivo. Iron regulating proteins (eg, ceruloplasmin, transferrin, membrane protein transferrin receptor 1, prostate 6 transmembrane epithelial antigen 3 (Steap3), divalent metal transporter 1 (DMT1) or family of zinc-iron regulatory proteins 8/14, and ferritin) participate in iron-binding, iron transport, and maintain iron homeostasis.Citation91–93 In a pathologic state, an increase in iron absorption and a decrease in iron storage lead to intracellular iron accumulation and ferroptosis,Citation68 which can reprogram iron uptake, production, utilization, and storage of intracellular iron.Citation93 Iron chelators (eg, deferoxamine and deferiprone) can bind to free iron to inactivate iron-containing enzymes and inhibit Fenton reactions, as well as down-regulate hepcidin expression.Citation94,Citation95 A new study showed that the iron chelator PBT434 [5,7-dichloro-2-((ethylamino)methyl)-8-hydroxy-3-methylquinazolin-4(3H)-one] chelates interstitial iron to inhibit iron uptake and stimulate iron efflux in brain microvascular endothelial cells and other cells of the neurovascular unit.Citation96 In addition, the tetracycline class antibiotic minocycline, an iron-chelating agent, has an anti-inflammatory effect.Citation97 Various iron chelators exhibit cerebral protection against acute traumatic and non-traumatic brain injuries.Citation12,Citation89,Citation98 For example, the brain-permeable iron chelator VK28 (5-[4-(2-hydroxyethyl) piperazine-1-ylmethyl]-quinoline-8-ol), the lipid-permeable iron chelator 2.2’-dipyridyl, and the iron chelator deferoxamine currently in clinical use decrease iron accumulation, ROS production, microglial activation, and neuronal death. Treatment with these iron chelators can protect the brain against iron toxicity and improve neurologic recovery after ICH.Citation99
(5) Regulation of PE biosynthesis: The members of the long-chain family of acyl-coenzyme A synthase 4 and the lysophosphatidylcholine acyltransferase 3 involved in PE biosynthesis are vital enzymes that can regulate lipid peroxidation and ferroptosis.Citation100,Citation101 A new study showed that energy- or stress-mediated activation of AMP protein kinase inhibits ferroptosis by phosphorylation and inactivation of acetyl-CoA carboxylase and subsequently decreases PUFA biosynthesis.Citation102 Therefore, given that inhibition of the expression of members of the long-chain family of acyl-coenzyme A synthase 4 attenuates oxidative stress, BBB damage, brain edema, and behavioral and cognitive deficits after subarachnoid hemorrhage,Citation14 it is necessary to investigate whether inhibition of ferroptosis can prevent or reduce brain injury in patients with COVID-19.
The Potential Role of Ferroptosis in COVID-19-Related Brain Injury
Ferroptosis May Exist in COVID-19-Related Brain Injuries
The brains of patients with COVID-19 could be susceptible to ferroptosis. Ferroptosis involves the following three factors: iron, amino acids, and unsaturated fatty acids.Citation103 Iron is the most abundant trace metal in the brain and is needed for normal cell metabolism and contributes to physiologic processes such as oxygen transport, energy production, and transmission of neurotransmitters.Citation104 The brain is sensitive to oxidative stress and lipid peroxidation due to its high level of PUFAs.Citation103 PUFA phospholipids are constituents of the cell membrane and carry out various physiologic functions (eg, neuronal synaptic connection, neuronal plasticity, and network development, as well as neurotransmitter release). They are also involved in the pathogenesis of ferroptosis.Citation105 The neuron cell membranes are rich in PUFAs and cholesterol that are susceptible to ROS-mediated oxidation caused by a cytokine storm in COVID-19.Citation103 The enzyme activity of GPX4 and superoxide dismutase is much lower in brain tissues compared to other tissues, indicating that neurons could be highly susceptible to iron overload.Citation90 SLC7A11 (also known as system Xc-) expressed in glial cells enhances glutamate release, thus inducing neuronal ferroptosis.Citation106 Astrocytes can store iron and prevent iron overload in neuronal synapses.Citation107 Nrf2 overexpression in astrocytes inhibits ferroptosis, probably due to the outward release of GSH from astrocytes.Citation108 Meanwhile, inducible loss of GPX4 expression in neurons can induce ferroptosis.Citation67
Neuronal death and the resultant neurologic dysfunction occur in patients with brain injuries, and ferroptosis is one of the death mechanisms. Ferroptosis is pathogenic in aging, tissue repair, tumor development, cerebral ischemiaCitation109 and ICH,Citation86,Citation110 and is involved in the pathogenesis of spinal cord injury,Citation111 subarachnoid hemorrhage,Citation14 traumatic brain injury,Citation112 epilepsy,Citation113 Alzheimer’s disease,Citation15 Parkinson’s disease,Citation16 and other neurodegenerative diseases. In addition, the disruption of iron homeostasis is present in almost all neuropathological specimens of neurological disorders,Citation67 including COVID-19-related brain injuries.Citation1 Therefore, like other brain diseases, ferroptosis may also exist in COVID-19 brain injury and plays a vital role in the process of brain injuries.
The pathogenesis of various complications of COVID-19 and brain injury, in particular, may be closely related to an imbalance of intracellular iron homeostasis that leads to ferroptosis.Citation1 Ferritin stores large amounts of iron within a cell. The level of ferritin in the CSF may reflect the level of iron in the brain.Citation114 Two recent studies revealed elevated serum levels of iron and ferritin associated with cerebral ischemia or disease severity in COVID-19 patients.Citation115,Citation116 A cohort study with 39 COVID-19 patients demonstrated that serum ferritin levels are correlated with the severity of COVID-19 diseaseCitation116 and are related to hyperferritinemia syndrome.Citation117 Since iron and ferritin levels are associated with ferroptosis,Citation118 we speculated that the severity of COVID-19 is also related to the occurrence of ferroptosis. Another study of 6 patients with COVID-19 complicated with ischemic stroke revealed that all 6 patients were in a high-grade prethrombotic state, with high blood levels of D-dimer and ferritin.Citation115 Patients with COVID-19 and stroke have higher ferritin levels than patients with stroke alone, and ICH patients have higher serum ferritin levels than patients with ischemic stroke.Citation119 Although, to date, no reports have directly linked ferroptosis and COVID-19-related brain injury, these biomarkers of ferroptosis are altered in COVID-19-related brain injury, suggesting that ferroptosis may exist and indeed play an underlying role in disease progression.
Three contributing factors may cause ferroptosis in COVID-19-related brain injury according to recent studies: i) hepcidin release, the principal regulator of systemic iron homeostasis; ii) excessive iron influx through the transferrin receptor during SARS-CoV-2 replication; and iii) SARS-CoV-2 attack on hemoglobin and release of free iron into the circulation.Citation120–124 IL-6 can initially stimulate the synthesis of ferritin and hepcidin in a COVID-19-related cytokine storm.Citation120,Citation121 The amino acid sequence at the distal end of the cytoplasmic tail of the SARS-CoV-2 spike glycoprotein has highly similar to that of the hepcidin protein.Citation122 Hepcidin and hepcidin-like proteins bind to ferroportin, the cellular iron exporter, causing its degradation, thus preventing iron outflow and increasing intracellular ferritin.Citation123 In addition, iron is required for SARS-CoV-2 replication and enters cells through transferrin receptors.Citation124 Excess intracellular iron interacts with molecular oxygen to trigger the Fenton reaction and induce ferroptosis when GPX4 does not eliminate excess lipid ROS.
Furthermore, hemoglobin, attacked by SARS-CoV-2, dissociates porphyrin from iron and releases free iron into the circulation, resulting in iron overload.Citation125,Citation126 Dysfunctional iron metabolism in COVID-19 finally leads to ferroptosis, releasing intracellular ferritin and causing ferritinemia.Citation118 Together, this current evidence suggests that ferroptosis may exist in COVID-19-related brain injury and that ferroptosis plays a role in brain complications in COVID-19.
The Potential Role of Ferroptosis Underlies COVID-19-Related Brain Injury
Ferroptosis can induce the release of pathogen-associated molecular patterns and damage-associated molecular patterns, together with excessive oxidative stress.Citation127 Ferroptosis can cause local brain damage, leading to neuronal death and neural network damage.Citation104 Additionally, the sensitivity of oligodendrocytes to iron can cause axonal damage through ferroptosis.Citation104 Ferroptosis can also induce tau phosphorylation and form neurofibrillary tangles, leading to cognitive impairment.Citation76 Ferroptosis is also immunogenic and proinflammatory and creates a positive feedback loop.Citation127 As a result, ferroptosis may be one of the causes of COVID-19-related enhanced inflammatory reaction and multiple organ failure syndromes involving the brain with CNS symptomsCitation17 (). COVID-19-related brain injury contains many kinds of brain disease, such as stroke. Our previous studies have shown that stroke can cause ferroptosis.Citation98,Citation128 Iron toxicity contributes to stroke-induced early brain damage, and ferroptosis plays a significant role in the secondary brain injury from stroke.Citation12 Therefore, ferroptosis may be the effect of COVID-19-related stroke, but in turn, it plays a vital cause in the secondary brain injury of COVID-19-related stroke and further aggravate brain damage.
Figure 2 The potential role of ferroptosis in COVID-19-related brain injury. After SARS-CoV-2 infection, transferrin receptors recognize transferrin that carries Fe3+, which enters cells through endocytosis to form endosomes. IL-6 promotes ferritin synthesis, which stores Fe3+ and releases Fe3+ through ferritinophagy; after that, the endoplasmic reticulum metal reductase Steap3 reduces Fe3+ into Fe2+ to form LIP. GPX4 protects the cell against lipid peroxidation and inhibits ferroptosis. Under iron overload conditions combined with GPX4 depletion or inhibition, mitochondria generate large amounts of ROS, leading to lipid peroxidation, cell membrane damage, and ferroptosis. Excess ROS depletes intracellular GSH, and this depletion forms a positive feedback loop and aggravates lipid peroxidation. During this peroxidation, damage-associated molecular patterns and alarmins (eg, HMGB1, IL-33, TNF-α, IL-1 β, and IL-6) are released that activate NF-kB and other proinflammatory signaling pathways, eventually leading to neuroinflammation and cell death. SARS-CoV-2 infection can damage the brain, resulting in BBB disruption and bleeding accompanied by a cytokine storm.
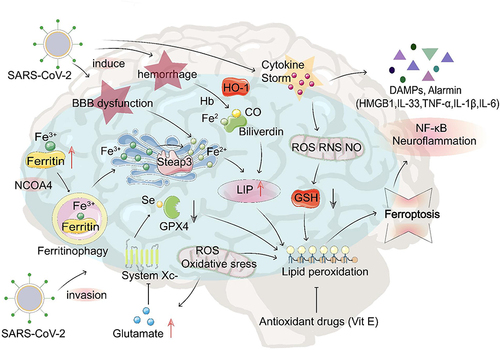
After SARS-CoV-2 infection, transferrin recognized by transferrin receptors carries excess Fe3+ into cells. The metal reductase Steap3 in the endoplasmic reticulum reduces Fe3+, and ferric iron enters the cytoplasm to form LIP. Under conditions of intracellular iron overload, the interaction between excess intracellular iron and molecular oxygen induces a Fenton reaction, producing excessive hydroxyl radicals.Citation129 Phospholipid hydroperoxide is produced by lipoxygenase/arachidonic acid lipoxygenase-mediated peroxidation of PE phospholipids containing PUFA.Citation76 When GPX4 activity is inhibited, phospholipid hydroperoxide enhances lipid peroxidation of plasma membranes, nucleic acids, and proteins, causing oxidative damage and ferroptosis.Citation72 Finally, damage-associated molecular patterns and alarmins (HMGB1, IL-33, and TNF) are released, exacerbating inflammation, cell death, and neurodegeneration.Citation127
During the cytokine storm in COVID-19, levels of proinflammatory mediators (eg, NF-κB, TNF-α, and IL-1β) are increased in the brain, leading to increased expression of AQP4 that promotes brain edema.Citation130 These factors are also involved in the mechanisms of cell ferroptosis.Citation90 The iron accumulation in the area surrounding the edema can cause secondary brain damage.Citation12 The formation and enlargement of edema in the brain can cause a compressive effect and increased intracranial pressure, leading to brain herniation and death.Citation131 GPX4 overexpression reduces cerebral edema, while treatment with Fer-1, an inhibitor of lipid peroxidation, reduces the proinflammatory cytokine levels (eg, NF-κB, TNF-α, and IL-1β) in the brain and reduces cerebral edema.Citation132
As discussed in SARS-CoV-2 Invasion with Subsequent Brain Injuries, the cytokine storm can cause BBB dysfunction,Citation34 which, together with inflammation in the injured brain of COVID-19 patients, allows more iron to enter the brain. Ferritin transported to the brain must pass through the BBB under the control of DMT1.Citation133 However, the gene that encodes DMT1 has binding sites for AP-1 and NF-κB,Citation133 both of which are potent inflammatory factors. Since COVID-19-induced cytokine storm produces numerous proinflammatory factors, including AP-1 and NF-κB,Citation134 it can lead to increased DMT1 expression, thus in turn, more iron is transported to the brain, causing iron accumulation or overload, thereby causing neuronal death and aggravated brain damage.Citation135 Previous studies have shown that intracellular iron influx and increased DMT1 expression are early downstream responses of NF-kB activation that accelerate neuronal damage in the brain.Citation133 A study involving four COVID-19 patients revealed that three patients had an elevated CSF/serum albumin index, indicating BBB dysfunction; simultaneously, the CSF level of the inflammatory cytokine IL-6 was increased in two patients, indicating a proinflammatory response in the brain.Citation36 The cytokine storm in COVID-19 produces substantial oxidative stress, which inactivates transferrin. Releasing a large amount of “free iron” increases serum iron levels,Citation68 leading to ferritinemia.Citation118
Elevated iron levels in the brains of COVID-19 patients can damage brain tissue. The toxic effects of iron overload on carbohydrates, lipids, proteins, and nucleic acids in the brain have been established.Citation136 Iron deposition leads to lipid peroxidation,Citation12,Citation103 the most effective inducer of ferroptosis.Citation137 During the severe cytokine storm of COVID-19, excessive increases in the circulating “free iron” will aggravate the proinflammatory response and induce morphological changes in red blood cells and fibrin. The resulting production of hydroxyl free radicals causes oxidative stress. It generates a remarkable procoagulation state that promotes the formation of dense clots in the brain and leads to ischemic stroke.Citation117
A recent study documented the formation of cerebral thrombosis and the appearance of stroke in patients with COVID-19.Citation138 The cerebral blood vessels rupture after a hemorrhagic stroke in COVID-19 patients, allowing blood to accumulate within the brain parenchyma. Heme oxygenase degrades hemoglobin metabolites into ferrous iron, biliverdin, and carbon monoxide in the brain.Citation125,Citation126 This metabolic pathway modulates secondary brain injury.Citation139 Iron released into the extracellular space can generate free radicals and ROS through the Fenton reaction, causing oxidative damage to molecules such as PUFAs, DNA, and proteins, thus further aggravating tissue damage.Citation13,Citation140 ROS activates microglia and astrocytes to release proinflammatory cytokines and increase the release of Fe2+ from LIP, leading to iron-dependent oxidative damage and neuronal cell death, exacerbating secondary brain injuries.Citation90 Compared to controls, mice with ischemic stroke had lower GPX4 activity and GSH levels. A subsequent increase in iron deposition and lipid peroxidation eventually resulted in ferroptosis.Citation109 In ICH, hemoglobin breaks down and releases large amounts of ferrous iron and mitochondrial atrophy; both are features of ferroptosis.Citation86,Citation87,Citation141 Furthermore, lipid peroxidation inhibitors such as Fer-1 protect against damage caused by ischemic and hemorrhagic stroke.Citation142 Similar pathogenic mechanisms can lead to COVID-19-related stroke.
Potential Therapeutic Targets for COVID-19-Related Brain Injury
Investigating the potential role of ferroptosis in COVID-19-related brain injury is an emerging research direction and may yield therapeutic targets (). Regulation of iron and ROS, Nrf2/KEAP1 pathway, GPX4-GSH pathway, as primary targets, can make a difference through antimalarial drugs, antioxidants, iron chelators, (-)-epicatechin, etc. These ferroptosis inhibitors may serve as a potential intervention to protect neurons against various brain injuries and patients with COVID-19-related brain injuries.Citation71,Citation110 Antimalarial drugs inhibit the expression of iron transporters and the activity of Fe2+/H+ cotransporters, thus reducing iron and iron-catalyzed free radicals in brain tissues.Citation135 Tocotrienol, a component of vitamin E, inhibits ferroptosis of neurons.Citation108 Nrf2 is also a promising target for reducing ferroptosis and treating brain injuries.Citation143 In this sense, we have shown that (-)-epicatechin, a brain-permeable flavanol, protects against ICH by activating the Nrf2-dependent and Nrf2-independent pathways and may serve as a potential intervention for patients with ICHCitation88,Citation144 or patients with COVID-19-related brain injuries.
Table 3 Potential Therapeutic Targets for COVID-19-Related Brain Injury
Concluding Remarks
The SARS-CoV-2 vaccines are essential to prevent further morbidity and mortality from COVID-19.Citation145 Although studies have found an increased risk of neurological complications in individuals who have been vaccinated against COVID-19, the risk of these complications is greater after testing positive for SARS-CoV-2.Citation146 Expert opinions indicate that it is advisable to correct the iron deficiency before administering the COVID-19 vaccine.Citation147 Iron is a nutrient required for immunity, and iron deficiency reduces the efficacy of vaccines.Citation147 However, no studies have investigated the role of the COVID-19 vaccine on ferroptosis. In theory, vaccination against COVID-19 would reduce the risk of COVID-19 infection, therefore reducing ferroptosis-mediated brain damage. We could only speculate that COVID-19 vaccines may reduce the occurrence of ferroptosis and may help recover from the disease on ferroptosis. This is an excellent direction for us to further research COVID-19.
Due to inflammation, iron and vitamin deficiencies, anemia is a common manifestation in COVID-19.Citation148 Studies have shown that serum iron levels are abnormally low in severe infections, but iron overload is high in pulmonary fibrotic tissue and the brain.Citation118,Citation149 Similarly, a significant serum iron deficiency can be detected in patients with COVID-19.Citation122,Citation150 Anemic patients with COVID-19 had elevated levels of inflammation markers (IL-6 levels and C-reactive protein) compared to those without anemia and survived a more severe course of COVID-19,Citation151,Citation152 but did not directly influence mortality.Citation148 Various evidence indicates that anemia and low serum iron concentration are independent risk factors associated with severe illness and poor outcomes in hospitalized patients and death from COVID-19.Citation150,Citation153,Citation154 A case report showed an 8.2 times greater chance of developing severe pneumonia in patients with anemia.Citation148,Citation151 In a cohort study, patients with anemia were more likely to have one or more comorbidities and severe COVID-19 illness compared to patients without anemia.Citation153
Furthermore, previous studies showed a significant association between prior iron deficiency anemia and ischemic stroke.Citation155 Transfusions have remained a considerable way to manage stroke so far.Citation156 Since no articles report the relationship, we could only speculate that anemia may be associated with COVID-19-related brain injury. Thus, immune-mediated disruption of iron homeostasis and ferroptosis, releasing intracellular ferritin and causing ferriminemia in patients with COVID-19, with a strong possibility of progressing to severe CNS disease.Citation157 The effects of anemia and iron deficiency severity on subsequent CNS manifestations will be worthwhile exploration.
The epidemiological survey showed that the median time from the onset of the first symptom to dyspnea was 5.0 days, 7.0 days to hospital admission, and 8.0 days to the intensive care unit.Citation20 The incubation period may be long enough for the virus to enter and destroy medulla neurons, leading to brain damage.Citation47 However, there have been no experiments or studies that investigate the time frame of ferroptosis in COVID-19. Changes in biomarkers of ferroptosis (eg, transferrin receptor, transferrin, and hepcidin) have not been reported in COVID-19.Citation158 Based on the hypothesis we put forward that ferroptosis occurs and aggravates brain injuries in COVID-19, ferroptosis can occur early or late after COVID-19 infection. Whether it is associated with neuropsychiatric disturbances in COVID-19 depends on whether it affects neuropsychiatric pathways, which needs further research. The effect of inhibiting ferroptosis by iron chelation on COVID-19-related injury may be an excellent direction worth exploring in depth.
Increasing evidence indicates that SARS-CoV-2 infection can cause various brain injuries with neuropsychiatric symptoms. Ferroptosis may be the basis for the pathogenic mechanisms of various brain injuries associated with the COVID-19 virus. BBB dysfunction and neuroinflammation in COVID-19 allow more iron to flow into brain tissues. Free iron released into the extracellular space generates free radicals, causing oxidative damage to molecules such as PUFAs, DNA, and proteins. Ferroptosis can also be induced by free iron and further aggravates brain tissue damage.Citation13,Citation140 Although there is no direct evidence linking ferroptosis with brain injuries from COVID-19, the rationale exists. Now we hypothesize that ferroptosis occurs and aggravates brain injuries in COVID-19. The potential role of ferroptosis in COVID-19-related brain injuries warrants careful study to identify promising therapeutic targets to combat SARS-CoV-2-induced brain injuries with neuropsychiatric symptoms.
Abbreviations
BBB, blood-brain barrier; CNS, central nervous system; GPX4, glutathione peroxidase 4; GSH, glutathione; Hb, hemoglobin; HMGB1, high mobility group box-1; HO-1, heme oxygenase-1; IL, interleukin; LIP, labile iron pool; NCOA4, nuclear receptor coactivator 4; NF-kB, nuclear factor kappa B; RNS, reactive nitrogen species; ROS, reactive oxygen species; Se, selenocysteine; Steap3, prostate 6 transmembrane epithelial antigen 3; TNF, tumor necrosis factor; VitE, vitamin E.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no conflicts of interest in this work.
Acknowledgements
Ruoyu Zhang, Chen Sun, and Yunze Han are undergraduate students.
Additional information
Funding
References
- Habib HM, Ibrahim S, Zaim A, Ibrahim WH. The role of iron in the pathogenesis of COVID-19 and possible treatment with lactoferrin and other iron chelators. Biomed Pharmacother. 2021;136:111228. doi:10.1016/j.biopha.2021.111228
- World Health Organization. Coronavirus (COVID-19) dashboard data table. Available from: https://covid19.who.int/. Accessed March 16, 2022.
- Liu JM, Tan BH, Wu S, et al. Evidence of central nervous system infection and neuroinvasive routes, as well as neurological involvement, in the lethality of SARS-CoV-2 infection. J Med Virol. 2021;93(3):1304–1313. doi:10.1002/jmv.26570
- Mao L, Jin H, Wang M, et al. Neurologic manifestations of hospitalized patients with coronavirus disease 2019 in Wuhan, China. JAMA Neurol. 2020;77(6):683–690. doi:10.1001/jamaneurol.2020.1127
- Najjar S, Najjar A, Chong DJ, et al. Central nervous system complications associated with SARS-CoV-2 infection: integrative concepts of pathophysiology and case reports. J Neuroinflammation. 2020;17(1):231. doi:10.1186/s12974-020-01896-0
- Divani AA, Andalib S, Biller J, et al. Central nervous system manifestations associated with COVID-19. Curr Neurol Neurosci Rep. 2020;20(12):60. doi:10.1007/s11910-020-01079-7
- Morgello S. Coronaviruses and the central nervous system. J Neurovirol. 2020;26(4):459–473. doi:10.1007/s13365-020-00868-7
- Boldrini M, Canoll PD, Klein RS. How COVID-19 affects the brain. JAMA Psychiatry. 2021;78(6):682–683. doi:10.1001/jamapsychiatry.2021.0500
- De Felice FG, Tovar-Moll F, Moll J, Munoz DP, Ferreira ST. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and the central nervous system. Trends Neurosci. 2020;43(6):355–357. doi:10.1016/j.tins.2020.04.004
- Kaseda ET, Levine AJ. Post-traumatic stress disorder: a differential diagnostic consideration for COVID-19 survivors. Clin Neuropsychol. 2020;34(7–8):1498–1514. doi:10.1080/13854046.2020.1811894
- Dixon SJ, Lemberg KM, Lamprecht MR, et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell. 2012;149(5):1060–1072. doi:10.1016/j.cell.2012.03.042
- Wan J, Ren H, Wang J. Iron toxicity, lipid peroxidation and ferroptosis after intracerebral haemorrhage. Stroke Vasc Neurol. 2019;4(2):93–95. doi:10.1136/svn-2018-000205
- Weiland A, Wang Y, Wu W, et al. Ferroptosis and its role in diverse brain diseases. Mol Neurobiol. 2019;56(7):4880–4893. doi:10.1007/s12035-018-1403-3
- Qu XF, Liang TY, Wu DG, et al. Acyl-CoA synthetase long chain family member 4 plays detrimental role in early brain injury after subarachnoid hemorrhage in rats by inducing ferroptosis. CNS Neurosci Ther. 2021;27(4):449–463. doi:10.1111/cns.13548
- Lane DJR, Ayton S, Bush AI. Iron and Alzheimer’s disease: an update on emerging mechanisms. J Alzheimers Dis. 2018;64(s1):S379–s395. doi:10.3233/JAD-179944
- Mahoney-Sánchez L, Bouchaoui H, Ayton S, et al. Ferroptosis and its potential role in the physiopathology of Parkinson’s disease. Prog Neurobiol. 2021;196:101890. doi:10.1016/j.pneurobio.2020.101890
- Yang M, Lai CL. SARS-CoV-2 infection: can ferroptosis be a potential treatment target for multiple organ involvement? Cell Death Discov. 2020;6:130. doi:10.1038/s41420-020-00369-w
- Fitsiori A, Pugin D, Thieffry C, Lalive P, Vargas MI. COVID-19 is associated with an unusual pattern of brain microbleeds in critically ill patients. J Neuroimaging. 2020;30(5):593–597. doi:10.1111/jon.12755
- Rodriguez-Morales AJ, Cardona-Ospina JA, Gutiérrez-Ocampo E, et al. Clinical, laboratory and imaging features of COVID-19: a systematic review and meta-analysis. Travel Med Infect Dis. 2020;34:101623. doi:10.1016/j.tmaid.2020.101623
- Wang D, Hu B, Hu C, et al. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus-infected pneumonia in Wuhan, China. JAMA. 2020;323(11):1061–1069. doi:10.1001/jama.2020.1585
- Beltrán-Corbellini Á, Chico-García JL, Martínez-Poles J, et al. Acute-onset smell and taste disorders in the context of COVID-19: a pilot multicentre polymerase chain reaction based case-control study. Eur J Neurol. 2020;27(9):1738–1741. doi:10.1111/ene.14273
- Koralnik IJ, Tyler KL. COVID-19: a global threat to the nervous system. Ann Neurol. 2020;88(1):1–11. doi:10.1002/ana.25807
- Zhou Y, Xu J, Hou Y, et al. Network medicine links SARS-CoV-2/COVID-19 infection to brain microvascular injury and neuroinflammation in dementia-like cognitive impairment. Alzheimers Res Ther. 2021;13(1):110. doi:10.1186/s13195-021-00850-3
- Helms J, Kremer S, Merdji H, et al. Neurologic features in severe SARS-CoV-2 infection. N Engl J Med. 2020;382(23):2268–2270. doi:10.1056/NEJMc2008597
- Jaywant A, Vanderlind WM, Alexopoulos GS, et al. Frequency and profile of objective cognitive deficits in hospitalized patients recovering from COVID-19. Neuropsychopharmacology. 2021:1–6.
- Zombori L, Bacon M, Wood H, et al. Severe cortical damage associated with COVID-19 case report. Seizure. 2021;84:66–68. doi:10.1016/j.seizure.2020.11.014
- Lee MH, Perl DP, Nair G, et al. Microvascular injury in the brains of patients with Covid-19. N Engl J Med. 2021;384(5):481–483. doi:10.1056/NEJMc2033369
- Pajo AT, Espiritu AI, Apor A, Jamora RDG. Neuropathologic findings of patients with COVID-19: a systematic review. Neurol Sci. 2021;42(4):1255–1266. doi:10.1007/s10072-021-05068-7
- Markus HS, Brainin M. COVID-19 and stroke-A global World Stroke Organization perspective. Int J Stroke. 2020;15(4):361–364. doi:10.1177/1747493020923472
- Wijeratne T, Sales C, Karimi L, Crewther SG. Acute ischemic stroke in COVID-19: a case-based systematic review. Front Neurol. 2020;11:1031. doi:10.3389/fneur.2020.01031
- Vogrig A, Gigli GL, Bnà C, Morassi M. Stroke in patients with COVID-19: clinical and neuroimaging characteristics. Neurosci Lett. 2021;743:135564. doi:10.1016/j.neulet.2020.135564
- Li Y, Li M, Wang M, et al. Acute cerebrovascular disease following COVID-19: a single center, retrospective, observational study. Stroke Vasc Neurol. 2020;5(3):279–284. doi:10.1136/svn-2020-000431
- Li K, Wohlford-Lenane C, Perlman S, et al. Middle east respiratory syndrome coronavirus causes multiple organ damage and lethal disease in mice transgenic for human dipeptidyl peptidase 4. J Infect Dis. 2016;213(5):712–722. doi:10.1093/infdis/jiv499
- Li Z, Liu T, Yang N, et al. Neurological manifestations of patients with COVID-19: potential routes of SARS-CoV-2 neuroinvasion from the periphery to the brain. Front Med. 2020;14(5):533–541. doi:10.1007/s11684-020-0786-5
- Yachou Y, El Idrissi A, Belapasov V, Ait Benali S. Neuroinvasion, neurotropic, and neuroinflammatory events of SARS-CoV-2: understanding the neurological manifestations in COVID-19 patients. Neurol Sci. 2020;41(10):2657–2669. doi:10.1007/s10072-020-04575-3
- Perrin P, Collongues N, Baloglu S, et al. Cytokine release syndrome-associated encephalopathy in patients with COVID-19. Eur J Neurol. 2021;28(1):248–258. doi:10.1111/ene.14491
- Guadarrama-Ortiz P, Choreño-Parra JA, Sánchez-Martínez CM, et al. Neurological aspects of SARS-CoV-2 infection: mechanisms and manifestations. Front Neurol. 2020;11:1039. doi:10.3389/fneur.2020.01039
- Michetti F, D’Ambrosi N, Toesca A, et al. The S100B story: from biomarker to active factor in neural injury. J Neurochem. 2019;148(2):168–187. doi:10.1111/jnc.14574
- Kadono Y, Nakamura Y, Ogawa Y, et al. A case of COVID-19 infection presenting with a seizure following severe brain edema. Seizure. 2020;80:53–55. doi:10.1016/j.seizure.2020.06.015
- Vinayagam S, Sattu K. SARS-CoV-2 and coagulation disorders in different organs. Life Sci. 2020;260:118431. doi:10.1016/j.lfs.2020.118431
- Al Saiegh F, Ghosh R, Leibold A, et al. Status of SARS-CoV-2 in cerebrospinal fluid of patients with COVID-19 and stroke. J Neurol Neurosurg Psychiatry. 2020;91(8):846–848. doi:10.1136/jnnp-2020-323522
- Meinhardt J, Radke J, Dittmayer C, et al. Olfactory transmucosal SARS-CoV-2 invasion as a port of central nervous system entry in individuals with COVID-19. Nat Neurosci. 2021;24(2):168–175. doi:10.1038/s41593-020-00758-5
- Mahalaxmi I, Kaavya J, Mohana Devi S, Balachandar V. COVID-19 and olfactory dysfunction: a possible associative approach towards neurodegenerative diseases. J Cell Physiol. 2021;236(2):763–770. doi:10.1002/jcp.29937
- Chiu A, Fischbein N, Wintermark M, et al. COVID-19-induced anosmia associated with olfactory bulb atrophy. Neuroradiology. 2021;63(1):147–148. doi:10.1007/s00234-020-02554-1
- Zhou L, Xu Z, Castiglione GM, et al. ACE2 and TMPRSS2 are expressed on the human ocular surface, suggesting susceptibility to SARS-CoV-2 infection. Ocul Surf. 2020;18(4):537–544. doi:10.1016/j.jtos.2020.06.007
- Panariello F, Cellini L, Speciani M, De Ronchi D, Atti AR. How does SARS-CoV-2 affect the central nervous system? A working hypothesis. Front Psychiatry. 2020;11:582345. doi:10.3389/fpsyt.2020.582345
- Li YC, Bai WZ, Hashikawa T. The neuroinvasive potential of SARS-CoV2 may play a role in the respiratory failure of COVID-19 patients. J Med Virol. 2020;92(6):552–555. doi:10.1002/jmv.25728
- Virhammar J, Kumlien E, Fällmar D, et al. Acute necrotizing encephalopathy with SARS-CoV-2 RNA confirmed in cerebrospinal fluid. Neurology. 2020;95(10):445–449. doi:10.1212/WNL.0000000000010250
- Bostancıklıoğlu M. Temporal correlation between neurological and gastrointestinal symptoms of SARS-CoV-2. Inflamm Bowel Dis. 2020;26(8):e89–e91. doi:10.1093/ibd/izaa131
- Andriuta D, Roger PA, Thibault W, et al. COVID-19 encephalopathy: detection of antibodies against SARS-CoV-2 in CSF. J Neurol. 2020;267(10):2810–2811. doi:10.1007/s00415-020-09975-1
- Satarker S, Nampoothiri M. Involvement of the nervous system in COVID-19: the bell should toll in the brain. Life Sci. 2020;262:118568. doi:10.1016/j.lfs.2020.118568
- Dong M, Zhang J, Ma X, et al. ACE2, TMPRSS2 distribution and extrapulmonary organ injury in patients with COVID-19. Biomed Pharmacother. 2020;131:110678. doi:10.1016/j.biopha.2020.110678
- Pezzini A, Padovani A. Lifting the mask on neurological manifestations of COVID-19. Nat Rev Neurol. 2020;16(11):636–644. doi:10.1038/s41582-020-0398-3
- Aghagoli G, Gallo Marin B, Katchur NJ, et al. Neurological involvement in COVID-19 and potential mechanisms: a review. Neurocrit Care. 2021;34(3):1062–1071. doi:10.1007/s12028-020-01049-4
- Terpos E, Ntanasis-Stathopoulos I, Elalamy I, et al. Hematological findings and complications of COVID-19. Am J Hematol. 2020;95(7):834–847. doi:10.1002/ajh.25829
- Loganathan S, Kuppusamy M, Wankhar W, et al. Angiotensin-converting enzyme 2 (ACE2): COVID 19 gate way to multiple organ failure syndromes. Respir Physiol Neurobiol. 2021;283:103548. doi:10.1016/j.resp.2020.103548
- Girija ASS, Shankar EM, Larsson M. Could SARS-CoV-2-induced hyperinflammation magnify the severity of coronavirus disease (CoViD-19) leading to acute respiratory distress syndrome? Front Immunol. 2020;11:1206. doi:10.3389/fimmu.2020.01206
- Zhu H, Wang Z, Yu J, et al. Role and mechanisms of cytokines in the secondary brain injury after intracerebral hemorrhage. Prog Neurobiol. 2019;178:101610. doi:10.1016/j.pneurobio.2019.03.003
- De Santis G. SARS-CoV-2: a new virus but a familiar inflammation brain pattern. Brain Behav Immun. 2020;87:95–96. doi:10.1016/j.bbi.2020.04.066
- Grohmann U, Mondanelli G, Belladonna ML, et al. Amino-acid sensing and degrading pathways in immune regulation. Cytokine Growth Factor Rev. 2017;35:37–45. doi:10.1016/j.cytogfr.2017.05.004
- Bouças AP, Rheinheimer J, Lagopoulos J. Why severe COVID-19 patients are at greater risk of developing depression: a molecular perspective. Neuroscientist. 2020:1073858420967892. doi:10.1177/1073858420967892
- Turski WA, Wnorowski A, Turski GN, Turski CA, Turski L. AhR and IDO1 in pathogenesis of Covid-19 and the “Systemic AhR Activation Syndrome:” a translational review and therapeutic perspectives. Restor Neurol Neurosci. 2020;38(4):343–354. doi:10.3233/RNN-201042
- Engin AB, Engin ED, Engin A. The effect of environmental pollution on immune evasion checkpoints of SARS-CoV-2. Environ Toxicol Pharmacol. 2021;81:103520. doi:10.1016/j.etap.2020.103520
- Ng J, Papandreou A, Heales SJ, Kurian MA. Monoamine neurotransmitter disorders–clinical advances and future perspectives. Nat Rev Neurol. 2015;11(10):567–584. doi:10.1038/nrneurol.2015.172
- Li J, Cao F, Yin HL, et al. Ferroptosis: past, present and future. Cell Death Dis. 2020;11(2):88. doi:10.1038/s41419-020-2298-2
- Amaral EP, Costa DL, Namasivayam S, et al. A major role for ferroptosis in Mycobacterium tuberculosis-induced cell death and tissue necrosis. J Exp Med. 2019;216(3):556–570. doi:10.1084/jem.20181776
- Conrad M, Kagan VE, Bayir H, et al. Regulation of lipid peroxidation and ferroptosis in diverse species. Genes Dev. 2018;32(9–10):602–619. doi:10.1101/gad.314674.118
- Bogdan AR, Miyazawa M, Hashimoto K, Tsuji Y. Regulators of iron homeostasis: new players in metabolism, cell death, and disease. Trends Biochem Sci. 2016;41(3):274–286. doi:10.1016/j.tibs.2015.11.012
- Gao M, Yi J, Zhu J, et al. Role of mitochondria in ferroptosis. Mol Cell. 2019;73(2):354–363.e3. doi:10.1016/j.molcel.2018.10.042
- Ibrahim WH, Habib HM, Kamal H, St clair DK, Chow CK. Mitochondrial superoxide mediates labile iron level: evidence from Mn-SOD-transgenic mice and heterozygous knockout mice and isolated rat liver mitochondria. Free Radic Biol Med. 2013;65:143–149. doi:10.1016/j.freeradbiomed.2013.06.026
- Xie Y, Hou W, Song X, et al. Ferroptosis: process and function. Cell Death Differ. 2016;23(3):369–379. doi:10.1038/cdd.2015.158
- Sun Y, Chen P, Zhai B, et al. The emerging role of ferroptosis in inflammation. Biomed Pharmacother. 2020;127:110108. doi:10.1016/j.biopha.2020.110108
- Song X, Zhu S, Chen P, et al. AMPK-mediated BECN1 phosphorylation promotes ferroptosis by directly blocking system X(c)(-) activity. Curr Biol. 2018;28(15):2388–2399.e5. doi:10.1016/j.cub.2018.05.094
- Guo Y, Liu X, Liu D, et al. Inhibition of BECN1 suppresses lipid peroxidation by increasing system X(c)(-) activity in early brain injury after subarachnoid hemorrhage. J Mol Neurosci. 2019;67(4):622–631. doi:10.1007/s12031-019-01272-5
- Forcina GC, Dixon SJ. GPX4 at the crossroads of lipid homeostasis and ferroptosis. Proteomics. 2019;19(18):e1800311. doi:10.1002/pmic.201800311
- Stockwell BR, Friedmann Angeli JP, Bayir H, et al. Ferroptosis: a regulated cell death nexus linking metabolism, redox biology, and disease. Cell. 2017;171(2):273–285. doi:10.1016/j.cell.2017.09.021
- Yang WS, SriRamaratnam R, Welsch ME, et al. Regulation of ferroptotic cancer cell death by GPX4. Cell. 2014;156(1–2):317–331. doi:10.1016/j.cell.2013.12.010
- Sui X, Zhang R, Liu S, et al. RSL3 drives ferroptosis through GPX4 inactivation and ROS production in colorectal cancer. Front Pharmacol. 2018;9:1371. doi:10.3389/fphar.2018.01371
- Alim I, Caulfield JT, Chen Y, et al. Selenium drives a transcriptional adaptive program to block ferroptosis and treat stroke. Cell. 2019;177(5):1262–1279.e25. doi:10.1016/j.cell.2019.03.032
- Hassannia B, Wiernicki B, Ingold I, et al. Nano-targeted induction of dual ferroptotic mechanisms eradicates high-risk neuroblastoma. J Clin Invest. 2018;128(8):3341–3355. doi:10.1172/JCI99032
- Wen X, Wu J, Wang F, et al. Deconvoluting the role of reactive oxygen species and autophagy in human diseases. Free Radic Biol Med. 2013;65:402–410. doi:10.1016/j.freeradbiomed.2013.07.013
- Latunde-Dada GO. Ferroptosis: role of lipid peroxidation, iron and ferritinophagy. Biochim Biophys Acta Gen Subj. 2017;1861(8):1893–1900. doi:10.1016/j.bbagen.2017.05.019
- Shah R, Margison K, Pratt DA. The potency of diarylamine radical-trapping antioxidants as inhibitors of ferroptosis underscores the role of autoxidation in the mechanism of cell death. ACS Chem Biol. 2017;12(10):2538–2545. doi:10.1021/acschembio.7b00730
- Feng H, Stockwell BR. Unsolved mysteries: how does lipid peroxidation cause ferroptosis? PLoS Biol. 2018;16(5):e2006203. doi:10.1371/journal.pbio.2006203
- Shen L, Lin D, Li X, et al. Ferroptosis in acute central nervous system injuries: the future direction? Front Cell Dev Biol. 2020;8:594. doi:10.3389/fcell.2020.00594
- Li Q, Han X, Lan X, et al. Inhibition of neuronal ferroptosis protects hemorrhagic brain. JCI Insight. 2017;2(7):e90777. doi:10.1172/jci.insight.90777
- Li Q, Weiland A, Chen X, et al. Ultrastructural characteristics of neuronal death and white matter injury in mouse brain tissues after intracerebral hemorrhage: coexistence of ferroptosis, autophagy, and necrosis. Front Neurol. 2018;9:581. doi:10.3389/fneur.2018.00581
- Chang CF, Cho S, Wang J. (-)-Epicatechin protects hemorrhagic brain via synergistic Nrf2 pathways. Ann Clin Transl Neurol. 2014;1(4):258–271. doi:10.1002/acn3.54
- Qin D, Wang J, Le A, et al. Traumatic brain injury: ultrastructural features in neuronal ferroptosis, glial cell activation and polarization, and blood-brain barrier breakdown. Cells. 2021;10:5. doi:10.3390/cells10051009
- Tang S, Gao P, Chen H, et al. The role of iron, its metabolism and ferroptosis in traumatic brain injury. Front Cell Neurosci. 2020;14:590789. doi:10.3389/fncel.2020.590789
- Gao M, Monian P, Quadri N, Ramasamy R, Jiang X. Glutaminolysis and transferrin regulate ferroptosis. Mol Cell. 2015;59(2):298–308. doi:10.1016/j.molcel.2015.06.011
- DeGregorio-Rocasolano N, Martí-Sistac O, Gasull T. Deciphering the iron side of stroke: neurodegeneration at the crossroads between iron dyshomeostasis, excitotoxicity, and ferroptosis. Front Neurosci. 2019;13:85. doi:10.3389/fnins.2019.00085
- Zhou B, Liu J, Kang R, et al. Ferroptosis is a type of autophagy-dependent cell death. Semin Cancer Biol. 2020;66:89–100. doi:10.1016/j.semcancer.2019.03.002
- Cavezzi A, Troiani E, Corrao S. COVID-19: hemoglobin, iron, and hypoxia beyond inflammation. A narrative review. Clin Pract. 2020;10(2):1271. doi:10.4081/cp.2020.1271
- Liu W, Zhang S, Nekhai S, Liu S. Depriving iron supply to the virus represents a promising adjuvant therapeutic against viral survival. Curr Clin Microbiol Rep. 2020;7:13–19.
- Bailey DK, Clark W, Kosman DJ. The iron chelator, PBT434, modulates transcellular iron trafficking in brain microvascular endothelial cells. PLoS One. 2021;16(7):e0254794. doi:10.1371/journal.pone.0254794
- Dai S, Hua Y, Keep RF, et al. Minocycline attenuates brain injury and iron overload after intracerebral hemorrhage in aged female rats. Neurobiol Dis. 2019;126:76–84. doi:10.1016/j.nbd.2018.06.001
- Li Q, Wan J, Lan X, et al. Neuroprotection of brain-permeable iron chelator VK-28 against intracerebral hemorrhage in mice. J Cereb Blood Flow Metab. 2017;37(9):3110–3123. doi:10.1177/0271678X17709186
- Hua W, Chen X, Wang J, et al. Mechanisms and potential therapeutic targets for spontaneous intracerebral hemorrhage. Brain Hemorrhages. 2020;1(2):99–104. doi:10.1016/j.hest.2020.02.002
- Li Y, Feng D, Wang Z, et al. Ischemia-induced ACSL4 activation contributes to ferroptosis-mediated tissue injury in intestinal ischemia/reperfusion. Cell Death Differ. 2019;26(11):2284–2299. doi:10.1038/s41418-019-0299-4
- Sha W, Hu F, Xi Y, Chu Y, Bu S. Mechanism of ferroptosis and its role in type 2 diabetes mellitus. J Diabetes Res. 2021;2021:9999612. doi:10.1155/2021/9999612
- Lee H, Zandkarimi F, Zhang Y, et al. Energy-stress-mediated AMPK activation inhibits ferroptosis. Nat Cell Biol. 2020;22(2):225–234. doi:10.1038/s41556-020-0461-8
- Ren JX, Sun X, Yan XL, Guo ZN, Yang Y. Ferroptosis in neurological diseases. Front Cell Neurosci. 2020;14:218. doi:10.3389/fncel.2020.00218
- Huang S, Li S, Feng H, Chen Y. Iron metabolism disorders for cognitive dysfunction after mild traumatic brain injury. Front Neurosci. 2021;15:587197. doi:10.3389/fnins.2021.587197
- Ingold I, Berndt C, Schmitt S, et al. Selenium utilization by GPX4 is required to prevent hydroperoxide-induced ferroptosis. Cell. 2018;172(3):409–422.e21. doi:10.1016/j.cell.2017.11.048
- Chen D, Fan Z, Rauh M, et al. ATF4 promotes angiogenesis and neuronal cell death and confers ferroptosis in a xCT-dependent manner. Oncogene. 2017;36(40):5593–5608. doi:10.1038/onc.2017.146
- Codazzi F, Pelizzoni I, Zacchetti D, Grohovaz F. Iron entry in neurons and astrocytes: a link with synaptic activity. Front Mol Neurosci. 2015;8:18. doi:10.3389/fnmol.2015.00018
- Ratan RR. The chemical biology of ferroptosis in the central nervous system. Cell Chem Biol. 2020;27(5):479–498. doi:10.1016/j.chembiol.2020.03.007
- Tuo QZ, Lei P, Jackman KA, et al. Tau-mediated iron export prevents ferroptotic damage after ischemic stroke. Mol Psychiatry. 2017;22(11):1520–1530. doi:10.1038/mp.2017.171
- Chen J, Wang Y, Wu J, et al. The potential value of targeting ferroptosis in early brain injury after acute CNS disease. Front Mol Neurosci. 2020;13:110. doi:10.3389/fnmol.2020.00110
- Chen Y, Liu S, Li J, et al. The latest view on the mechanism of ferroptosis and its research progress in spinal cord injury. Oxid Med Cell Longev. 2020;2020:6375938. doi:10.1155/2020/6375938
- Xie BS, Wang YQ, Lin Y, et al. Inhibition of ferroptosis attenuates tissue damage and improves long-term outcomes after traumatic brain injury in mice. CNS Neurosci Ther. 2019;25(4):465–475. doi:10.1111/cns.13069
- Chen S, Chen Y, Zhang Y, et al. Iron metabolism and ferroptosis in epilepsy. Front Neurosci. 2020;14:601193. doi:10.3389/fnins.2020.601193
- Yan N, Zhang J. Iron metabolism, ferroptosis, and the links with Alzheimer’s disease. Front Neurosci. 2019;13:1443. doi:10.3389/fnins.2019.01443
- Beyrouti R, Adams ME, Benjamin L, et al. Characteristics of ischaemic stroke associated with COVID-19. J Neurol Neurosurg Psychiatry. 2020;91(8):889–891. doi:10.1136/jnnp-2020-323586
- Dahan S, Segal G, Katz I, et al. Ferritin as a marker of severity in COVID-19 patients: a fatal correlation. Isr Med Assoc J. 2020;22(8):494–500.
- Colafrancesco S, Alessandri C, Conti F, Priori R. COVID-19 gone bad: a new character in the spectrum of the hyperferritinemic syndrome? Autoimmun Rev. 2020;19(7):102573. doi:10.1016/j.autrev.2020.102573
- Edeas M, Saleh J, Peyssonnaux C. Iron: innocent bystander or vicious culprit in COVID-19 pathogenesis? Int J Infect Dis. 2020;97:303–305. doi:10.1016/j.ijid.2020.05.110
- Hernández-Fernández F, Sandoval Valencia H, Barbella-Aponte RA, et al. Cerebrovascular disease in patients with COVID-19: neuroimaging, histological and clinical description. Brain. 2020;143(10):3089–3103. doi:10.1093/brain/awaa239
- Daher R, Manceau H, Karim Z. Iron metabolism and the role of the iron-regulating hormone hepcidin in health and disease. Presse Med. 2017;46(12 Pt 2):e272–e278. doi:10.1016/j.lpm.2017.10.006
- Bessman NJ, Mathieu JRR, Renassia C, et al. Dendritic cell-derived hepcidin sequesters iron from the microbiota to promote mucosal healing. Science. 2020;368(6487):186–189. doi:10.1126/science.aau6481
- Ehsani S. COVID-19 and iron dysregulation: distant sequence similarity between hepcidin and the novel coronavirus spike glycoprotein. Biol Direct. 2020;15(1):19. doi:10.1186/s13062-020-00275-2
- Ganz T. Iron and infection. Int J Hematol. 2018;107(1):7–15. doi:10.1007/s12185-017-2366-2
- Frazer DM, Anderson GJ. The regulation of iron transport. Biofactors. 2014;40(2):206–214. doi:10.1002/biof.1148
- Wang J, Doré S. Heme oxygenase 2 deficiency increases brain swelling and inflammation after intracerebral hemorrhage. Neuroscience. 2008;155(4):1133–1141. doi:10.1016/j.neuroscience.2008.07.004
- Wang J, Doré S. Heme oxygenase-1 exacerbates early brain injury after intracerebral haemorrhage. Brain. 2007;130(Pt 6):1643–1652. doi:10.1093/brain/awm095
- Linkermann A, Stockwell BR, Krautwald S, Anders HJ. Regulated cell death and inflammation: an auto-amplification loop causes organ failure. Nat Rev Immunol. 2014;14(11):759–767. doi:10.1038/nri3743
- Wu H, Wu T, Li M, Wang J. Efficacy of the lipid-soluble iron chelator 2,2’-dipyridyl against hemorrhagic brain injury. Neurobiol Dis. 2012;45(1):388–394. doi:10.1016/j.nbd.2011.08.028
- Shah R, Shchepinov MS, Pratt DA. Resolving the role of lipoxygenases in the initiation and execution of ferroptosis. ACS Cent Sci. 2018;4(3):387–396. doi:10.1021/acscentsci.7b00589
- Rao KV, Reddy PV, Curtis KM, Norenberg MD. Aquaporin-4 expression in cultured astrocytes after fluid percussion injury. J Neurotrauma. 2011;28(3):371–381. doi:10.1089/neu.2010.1705
- Xiong XY, Wang J, Qian ZM, Yang QW. Iron and intracerebral hemorrhage: from mechanism to translation. Transl Stroke Res. 2014;5(4):429–441. doi:10.1007/s12975-013-0317-7
- Zhang Z, Wu Y, Yuan S, et al. Glutathione peroxidase 4 participates in secondary brain injury through mediating ferroptosis in a rat model of intracerebral hemorrhage. Brain Res. 2018;1701:112–125. doi:10.1016/j.brainres.2018.09.012
- Ingrassia R, Lanzillotta A, Sarnico I, et al. 1B/(-)IRE DMT1 expression during brain ischemia contributes to cell death mediated by NF-κB/RelA acetylation at Lys310. PLoS One. 2012;7(5):e38019. doi:10.1371/journal.pone.0038019
- Chen Z, Hua S. Transcription factor-mediated signaling pathways’ contribution to the pathology of acute lung injury and acute respiratory distress syndrome. Am J Transl Res. 2020;12(9):5608–5618.
- Ong WY, Go ML, Wang DY, Cheah IK, Halliwell B. Effects of antimalarial drugs on neuroinflammation-potential use for treatment of COVID-19-related neurologic complications. Mol Neurobiol. 2021;58(1):106–117. doi:10.1007/s12035-020-02093-z
- Raz E, Jensen JH, Ge Y, et al. Brain iron quantification in mild traumatic brain injury: a magnetic field correlation study. AJNR Am J Neuroradiol. 2011;32(10):1851–1856. doi:10.3174/ajnr.A2637
- Kagan VE, Mao G, Qu F, et al. Oxidized arachidonic and adrenic PEs navigate cells to ferroptosis. Nat Chem Biol. 2017;13(1):81–90. doi:10.1038/nchembio.2238
- Fifi JT, Mocco J. COVID-19 related stroke in young individuals. Lancet Neurol. 2020;19(9):713–715. doi:10.1016/S1474-4422(20)30272-6
- Wang J, Jiang C, Zhang K, et al. Melatonin receptor activation provides cerebral protection after traumatic brain injury by mitigating oxidative stress and inflammation via the Nrf2 signaling pathway. Free Radic Biol Med. 2019;131:345–355. doi:10.1016/j.freeradbiomed.2018.12.014
- Magtanong L, Dixon SJ. Ferroptosis and brain injury. Dev Neurosci. 2018;40(5–6):382–395. doi:10.1159/000496922
- Wu Y, Song J, Wang Y, et al. The potential role of ferroptosis in neonatal brain injury. Front Neurosci. 2019;13:115. doi:10.3389/fnins.2019.00115
- Wu JR, Tuo QZ, Lei P. Ferroptosis, a recent defined form of critical cell death in neurological disorders. J Mol Neurosci. 2018;66(2):197–206. doi:10.1007/s12031-018-1155-6
- Song S, Gao Y, Sheng Y, Rui T, Luo C. Targeting NRF2 to suppress ferroptosis in brain injury. Histol Histopathol. 2021;36(4):383–397. doi:10.14670/HH-18-286
- Lan X, Han X, Li Q, Wang J. (-)-Epicatechin, a natural flavonoid compound, protects astrocytes against hemoglobin toxicity via Nrf2 and AP-1 signaling pathways. Mol Neurobiol. 2017;54(10):7898–7907. doi:10.1007/s12035-016-0271-y
- Hodgson SH, Mansatta K, Mallett G, et al. What defines an efficacious COVID-19 vaccine? A review of the challenges assessing the clinical efficacy of vaccines against SARS-CoV-2. Lancet Infect Dis. 2021;21(2):e26–e35. doi:10.1016/S1473-3099(20)30773-8
- Patone M, Handunnetthi L, Saatci D, et al. Neurological complications after first dose of COVID-19 vaccines and SARS-CoV-2 infection. Nat Med. 2021;27(12):2144–2153. doi:10.1038/s41591-021-01556-7
- Drakesmith H, Pasricha SR, Cabantchik I, et al. Vaccine efficacy and iron deficiency: an intertwined pair? Lancet Haematol. 2021;8(9):e666–e669. doi:10.1016/S2352-3026(21)00201-5
- Bergamaschi G, Borrelli de Andreis F, Aronico N, et al. Anemia in patients with Covid-19: pathogenesis and clinical significance. Clin Exp Med. 2021;21(2):239–246. doi:10.1007/s10238-020-00679-4
- Gupta Y, Maciorowski D, Medernach B, et al. Iron dysregulation in COVID-19 and reciprocal evolution of SARS-CoV-2: natura nihil frustra facit. J Cell Biochem. 2022. doi:10.1002/jcb.30207
- Zhao K, Huang J, Dai D, et al. Serum iron level as a potential predictor of coronavirus disease 2019 severity and mortality: a retrospective study. Open Forum Infect Dis. 2020;7(7):ofaa250. doi:10.1093/ofid/ofaa250
- Chen C, Zhou W, Fan W, et al. Association of anemia and COVID-19 in hospitalized patients. Future Virol. 2021;16(7):459–466. doi:10.2217/fvl-2021-0044
- Sonnweber T, Boehm A, Sahanic S, et al. Persisting alterations of iron homeostasis in COVID-19 are associated with non-resolving lung pathologies and poor patients’ performance: a prospective observational cohort study. Respir Res. 2020;21(1):276. doi:10.1186/s12931-020-01546-2
- Tao Z, Xu J, Chen W, et al. Anemia is associated with severe illness in COVID-19: a retrospective cohort study. J Med Virol. 2021;93(3):1478–1488. doi:10.1002/jmv.26444
- Faghih Dinevari M, Somi MH, Sadeghi Majd E, Abbasalizad Farhangi M, Nikniaz Z. Anemia predicts poor outcomes of COVID-19 in hospitalized patients: a prospective study in Iran. BMC Infect Dis. 2021;21(1):170. doi:10.1186/s12879-021-05868-4
- Chang YL, Hung SH, Ling W, et al. Association between ischemic stroke and iron-deficiency anemia: a population-based study. PLoS One. 2013;8(12):e82952. doi:10.1371/journal.pone.0082952
- Ware RE, Helms RW. Stroke with transfusions changing to hydroxyurea (SWiTCH). Blood. 2012;119(17):3925–3932. doi:10.1182/blood-2011-11-392340
- Hirschhorn T, Stockwell BR. The development of the concept of ferroptosis. Free Radic Biol Med. 2019;133:130–143. doi:10.1016/j.freeradbiomed.2018.09.043
- Chen X, Comish PB, Tang D, Kang R. Characteristics and biomarkers of ferroptosis. Front Cell Dev Biol. 2021;9:637162. doi:10.3389/fcell.2021.637162
- Varatharaj A, Thomas N, Ellul MA, et al. Neurological and neuropsychiatric complications of COVID-19 in 153 patients: a UK-wide surveillance study. Lancet Psychiatry. 2020;7(10):875–882. doi:10.1016/S2215-0366(20)30287-X
- Paterson RW, Brown RL, Benjamin L, et al. The emerging spectrum of COVID-19 neurology: clinical, radiological and laboratory findings. Brain. 2020;143(10):3104–3120. doi:10.1093/brain/awaa240
- Hao F, Tam W, Hu X, et al. A quantitative and qualitative study on the neuropsychiatric sequelae of acutely ill COVID-19 inpatients in isolation facilities. Transl Psychiatry. 2020;10(1):355. doi:10.1038/s41398-020-01039-2
- Zhang J, Lu H, Zeng H, et al. The differential psychological distress of populations affected by the COVID-19 pandemic. Brain Behav Immun. 2020;87:49–50. doi:10.1016/j.bbi.2020.04.031
- Giacomelli A, Pezzati L, Conti F, et al. Self-reported olfactory and taste disorders in patients with severe acute respiratory coronavirus 2 infection: a cross-sectional study. Clin Infect Dis. 2020;71(15):889–890. doi:10.1093/cid/ciaa330
- Nalleballe K, Reddy Onteddu S, Sharma R, et al. Spectrum of neuropsychiatric manifestations in COVID-19. Brain Behav Immun. 2020;88:71–74. doi:10.1016/j.bbi.2020.06.020
- Moriguchi T, Harii N, Goto J, et al. A first case of meningitis/encephalitis associated with SARS-Coronavirus-2. Int J Infect Dis. 2020;94:55–58. doi:10.1016/j.ijid.2020.03.062
- Dinkin M, Gao V, Kahan J, et al. COVID-19 presenting with ophthalmoparesis from cranial nerve palsy. Neurology. 2020;95(5):221–223. doi:10.1212/WNL.0000000000009700
- Jiang RD, Liu MQ, Chen Y, et al. Pathogenesis of SARS-CoV-2 in transgenic mice expressing human angiotensin-converting enzyme 2. Cell. 2020;182(1):50–58.e8. doi:10.1016/j.cell.2020.05.027
- Chen R, Wang K, Yu J, et al. The spatial and cell-type distribution of SARS-CoV-2 receptor ACE2 in the human and mouse brains. Front Neurol. 2020;11:573095. doi:10.3389/fneur.2020.573095
- Qi J, Zhou Y, Hua J, et al. The scRNA-seq expression profiling of the receptor ACE2 and the cellular protease TMPRSS2 reveals human organs susceptible to SARS-CoV-2 infection. Int J Environ Res Public Health. 2021;18(1):284. doi:10.3390/ijerph18010284
- Buzhdygan TP, DeOre BJ, Baldwin-Leclair A, et al. The SARS-CoV-2 spike protein alters barrier function in 2D static and 3D microfluidic in vitro models of the human blood-brain barrier. bioRxiv. 2020. doi:10.1101/2020.06.15.150912
- Song E, Zhang C, Israelow B, et al. Neuroinvasion of SARS-CoV-2 in human and mouse brain. J Exp Med. 2021;218(3). doi:10.1084/jem.20202135
- Paniz-Mondolfi A, Bryce C, Grimes Z, et al. Central nervous system involvement by severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2). J Med Virol. 2020;92(7):699–702. doi:10.1002/jmv.25915
- Puelles VG, Lütgehetmann M, Lindenmeyer MT, et al. Multiorgan and renal tropism of SARS-CoV-2. N Engl J Med. 2020;383(6):590–592. doi:10.1056/NEJMc2011400
- Martin M, Paes VR, Cardoso EF, et al. Postmortem brain 7T MRI with minimally invasive pathological correlation in deceased COVID-19 subjects. Insights Imaging. 2022;13(1):7. doi:10.1186/s13244-021-01144-w
- Shintoku R, Takigawa Y, Yamada K, et al. Lipoxygenase-mediated generation of lipid peroxides enhances ferroptosis induced by erastin and RSL3. Cancer Sci. 2017;108(11):2187–2194. doi:10.1111/cas.13380