
Abstract
This study aimed to determine any alteration in the killing of Staphylococcus aureus in murine peritoneal macrophages when chloroquine (CQ) is used alone compared with when it is used in combination with ciprofloxacin (CIP) or azithromycin (AZM). The study also aimed to find out the implication of reactive oxygen species (ROS) production and cytokine release in the intracellular killing of S. aureus in macrophages. We present here data obtained with a model of S. aureus-infected mouse peritoneal macrophages in which the intracellular growth of the bacteria and the influence of antibiotics was monitored for 30, 60, and 90 minutes in the presence or absence of CQ along with the production of ROS and alteration in levels of antioxidant enzymes and cytokines. It was observed that S. aureus-triggered cytokine response was regulated when macrophages were co-cultured with CQ and AZM as compared with CQ stimulation only. It can be suggested that action of AZM in mediating bacterial killing is enhanced by the presence of CQ, indicating enhanced uptake of AZM during early infection that may be essential for bacteria killing by AZM. Reduction of oxidative stress burden on the S. aureus-infected macrophages may pave the way for better killing of internalized S. aureus by CQ plus ciprofloxacin (CIP) or CQ plus AZM. Based on these observations, one may speculate that in an inflammatory milieu, CQ loaded with AZM elicits a stronger proinflammatory response by increasing the intracellular uptake of AZM or CIP, thus enabling the immune system to mount a more robust and prolonged response against intracellular pathogens.
Introduction
Staphylococcus aureus is a major human pathogen causing significant morbidity and mortality in both community- and hospital-acquired infections.Citation1 S. aureus, which often causes chronic or relapsing disease, has been reported to persist as an opportunistic intracellular organism both in vitro and in vivo.Citation2 S. aureus, commonly known as an extracellular host pathogen, even stimulates the inflammatory response and thus undergoes an intracellular phase within professional phagocytes, which encompass cells of the monocytic/macrophage lineage.Citation3 An effective treatment for the infections caused by intracellular bacterial forms can only be achieved using an antibiotic that can concentrate sufficiently at the site of microbial residence and further maintain activity in the intracellular environment.Citation4,Citation5 However, it has been observed that many microbial pathogens are protected, inside the cell, from antibiotics to which they are susceptible in vitro.Citation6,Citation7 S. aureus has been reported to express a wide array of secreted and cell-surface-associated virulence factors to help evade immune responses.Citation8 The treatment of S. aureus infections has been increasingly problematic due to the high prevalence of multi-antibiotic-resistant strains, such as methicillin-resistant S. aureus,Citation9 and the emergence of vancomycin-resistant S. aureus strains.Citation10
S. aureus has been reported to survive within phagocytic cells both in polymorphonuclear neutrophils (PMN) and monocytes.Citation11 However, recent experiments assessing invasion and the intracellular survival of S. aureus in endothelial and epithelial cells, and macrophages and osteoblastsCitation12–Citation19 have suggested that such events may contribute to the persistence of S. aureus during infections such as septic arthritis.Citation20–Citation22 Moreover, it has long been known that professional phagocytes may serve as intracellular reservoirs of S. aureus.Citation23 In order to survive and induce infection, pathogenic bacteria have to cope with their changing environment, as well as continuous attacks by the host’s antimicrobial defense system.
S. aureus may become intracellular, at least within monocytes, macrophages, and PMN, when host defense mechanisms are activated.Citation24 The facultative intracellular persistence of staphylococci may play an important role in the pathogenesis, because this localization protects them from both humoral and cell-mediated immune responses. The intracellular habitat of S. aureus calls for antibiotics with intracellular activity toward S. aureus. However, antibiotics have routinely been tested only for their in vitro activity on extracellular bacteria. Obviously, the intracellular activity of antibiotics can significantly differ from that exerted extracellularly. In general it seems impossible to deduce the intracellular antibacterial activity of antibiotics from standard susceptibility tests.Citation25 Moreover, it has been reported that approximately one-third of clinical S. aureus isolates not only survived in but also killed their eukaryotic host cells, and this was accompanied by increased in vivo virulence.Citation26 Therefore, a successful anti-staphylococcal therapy should include the elimination of intracellular bacteria and the rescue of host cells from staphylococci-induced cell death.
Earlier studies on the intracellular accumulation and activity of ciprofloxacin (CIP) and levofloxacin using different cellular models of S. aureus-infected phagocytes have also reported a bacteriostatic rather than a bactericidal effect of fluoroquinolones.Citation27–Citation29 Indeed, to become active, the intracellular portion of the antibiotic must come into direct contact with S. aureus, which resides mainly in the phagolysosomes of phagocytic cells. Fluoroquinolones are able to diffuse in various subcellular compartments.Citation30 In particular, macrolides have potential “nonantibiotic” anti-inflammatory activity.Citation31 Macrolides strongly accumulate within phagocytes (the necessary basis of their intracellular bioactivity), and this cellular concentration can modify phagocytic cell activities.Citation32 In addition, the phagolysosomal environment is apt to inactivate fluoroquinolones. Indeed, phagolysosomes are characterized by a low pH (4.8–5.0)Citation33 and fluoroquinolones, including levofloxacin, are known to have a reduced activity in acidic mediaCitation34 in comparison to macrolides such as azithromycin (AZM).
In the study reported here, AZM, a macrolide, was essentially chosen because, bacteriostatic and time-dependent antibiotics accumulate in cells to a large extent, and are largely localized in acidic vacuoles.Citation38,Citation39 CIP, an example of a fluoroquinolone, was chosen because it shows a marked concentration-dependent bactericidal effect but accumulates quickly in cells and distributes in the cytosol.Citation40,Citation41
It has been reported that acid pH conditions markedly decrease macrolides’ activity – for example, the alkalinization of phagolysosomes markedly increases the intracellular activity of amikacin.Citation42 The activity of fluoroquinolones is, however, less affected by acid pH.Citation43 Although it has been suggested that the alkalinization of phagolysosomes markedly increases the intracellular activity of macrolides. Conditions such as pH 5.0 remarkably reduce the activity of AZM, whereas pH 7.3 restores its activity.Citation43 After chloroquine (CQ) treatment, pH quantification in the phagosome was not done, but there are reports supporting our hypothesis in relation to the concentration of CQ and time of incubation used in our study. In one study,Citation44 the treatment of human macrophages with 100 μM CQ raised the pH of Cryptococcus neoformans-laden phagosomes to 7.3, whereas the activity of CIP was less affected by pH. However, as far as we are aware, the macrophage response after S. aureus infection, especially in relation to ROS production and cytokine release by phagocytes during CIP or AZM treatment in alkalinized intra-lysosomal pH, has not yet been studied.
CQ is known to have anti-inflammatory effects and has been used in the treatment of various diseases including rheumatoid arthritis, septic shock, and discoid lupus erythematosus.Citation45 Some of the effects of CQ in these diseases appear through interfering with the cytokine responses of the host.Citation46 CQ is known to inhibit several functions of macrophages,Citation47 but its effect on macrophages production of ROS after S. aureus infection and its possible implication in the intracellular survival of S. aureus have not yet been reported as far as we are aware.
It has been seen that CQ administration attenuates the production of typical serum pro- and anti-inflammatory cytokines such as tumor necrosis factor alpha (TNF-α) and interleukin (IL)-10.Citation48 Moreover, it was confirmed that CQ exerts some anti-inflammatory effects through the downregulation of TNF-α production and signaling in macrophages, as well as cytokine pattern production.Citation49 Cooper and Mag-were reported that the lysosomotropic effects of CQ are widely responsible for its anti-inflammatory properties that a decrease in the production of proinflammatory cytokines such as interferon gamma (IFN-γ, TNF-α, IL-1, and IL-6), and also emphasized the importance of non-lysosomotropic mechanisms (it was shown that CQ could inhibit TNF-α release in macrophages through inhibition of TNF-α messenger RNA synthesis).Citation50
This study was attempted to determine whether there is any alteration in the killing of S. aureus in murine peritoneal macrophages when CQ is used alone compared with when it is used in combination with CIP or AZM. We also wanted to find out the implication of ROS production and cytokine release in the intracellular killing of S. aureus in macrophages. We present here data obtained from a model of S. aureus-infected mouse peritoneal macrophages in which the intracellular growth of the bacteria and the influence of antibiotics were monitored for 30, 60, and 90 minutes in the presence or absence of CQ. Murine peritoneal macrophages were selected because they are quite permissive toward a variety of intracellular infectious agents, allowing detailed analysis of the effects of antibiotics without too much interference from the host-derived mechanisms of defense. We selected commonly used antibiotics (CIP and AZM) on the basis of their uptake by macrophages and their intracellular activity during alkalinization of phagosomes. In fact, we noted that the S. aureus-triggered cytokine response was regulated when macrophages were co-cultured with CQ and AZM as compared with CQ stimulation only. Based on these observations, one may speculate that, in an inflammatory milieu, CQ loaded with AZM elicits a stronger proinflammatory response in neighboring macrophages than AZM alone, thus enabling the immune system to mount a more robust and prolonged response against intracellular pathogens.
Materials and methods
Maintenance of animals and cells
All experiments involving animals were conducted according to the protocols that had been approved by the Institutional Animal Ethics Committee (IAEC) of the Department of Physiology, University of Calcutta, under the guidance of the Indian Ministry of Environment and Forest’s Committee for the Purpose of Control and Supervision of Experiments on Animals (approval number: 820/04/ac/CPCSEA, dated September 5, 2013). Wild-type male Swiss albino mice were used throughout the study. To minimize the feeling of hypoxia or discomfort before and during dissection and tissue collection, mice were anaesthetized with an inhaled anesthetic (ether) before terminal surgery. Euthanasia was performed by general anesthesia followed by vital tissue removal using 2%–3% ether for induction and 1% for maintenance. Macrophages were prepared from the peritoneal fluids of thioglycolate-administered mice.
Preparation of bacteria for in vitro infection of peritoneal macrophages
The S. aureus strain AG-789, which was found to be methicillin resistant and catalase positive, was obtained from Apollo Gleneagles Hospital, Calcutta, West Bengal, India. The S. aureus strain (AG-789) was grown overnight in Luria Bertani broth (HiMedia Laboratories, Mumbai, India), diluted with fresh broth then cultured until the mid-logarithmic phase of growth. Bacteria were harvested, washed twice with sterile saline, and adjusted to the desired inoculums spectrophotometrically before infection (optical density [OD]620 =0.2 for 5.0×107 cells/mL for S. aureus) and the number of colony-forming units (CFU) in the desired inoculums was confirmed by serial dilution and culture on blood agar.Citation20–Citation22
Isolation and stimulation of peritoneal macrophages
The mice, aged 6–8 weeks and fed standard laboratory chow and water, were injected intraperitoneally with 2 mL of 4% sterile thioglycolate broth, and the resulting peritoneal exudates were harvested by lavage of the peritoneal cavities with endotoxin-free Hank’s solution 4–5 days later. Peritoneal macrophages were suspended in 0.83% ammonium chloride solution containing 10% (v/v) Tris buffer (pH 7.65) to lyse erythrocytes. The cells were resuspended in Roswell Park Memorial Institute (RPMI) 1640 medium supplemented with 10% fetal bovine serum (FBS), 100 IU/mL penicillin, and 100 μg/mL streptomycin. Cells were then kept in plastic petri dishes for their adherence to the surface of the plastic petri dishes. Non-adherent cells were removed by aspiration and washing with RPMI 1640 medium before the addition of S. aureus. The adherent macrophages, more than 95% of which appeared to be typical macrophages by light microscopy, were used for each experiment.Citation15–Citation17 The peritoneal macrophages so collected were suspended in Hank’s Balanced Salt Solution (HBSS). Murine peritoneal macrophages (5×106 cells/mL) were infected with S. aureus (5×106 CFU/mL) for 30, 60, and 90 minutes at 37°C, in a humidified, 5% CO2 Heal Force HF151 CO2 Incubator (Shanghai, People’s Republic of China) in the presence or absence of antibiotics and CQ at a dose of 5 μg/mLCitation34 either alone or in combination with AZM or CIP at a dose of four times the minimum inhibitory concentration (MIC) for the S. aureus strain (AG-789).
Assay for intracellular killing
The presence of a large number of S. aureus bacteria in some cells that had undergone in vitro culture observed in the bacterial plate count suggests that the bacteria grow intracellularly. In addition, it has been reported that S. aureus multiplies in the presence of intact macrophages in cell cultures and would not grow in macrophage-conditioned media.Citation15–Citation17 Therefore, the number of S. aureus bacteria grown in a petri dish after time-dependent phagocytosis indicates the number of S. aureus bacteria that have survived inside the macrophages after ingestion.
For this, murine peritoneal macrophages (5×106 cells/mL) were mixed with S. aureus (5×106 CFU/mL) in a cell to bacterium ratio of 1:1Citation15–Citation17 in RPMI-FBS (5%) and incubated at 37°C in a cell-culture incubator for different times in the presence and absence of antibiotics and CQ either alone or in combination. After centrifugation, cell-culture supernatants were collected and stored for further assay at −70°C. Phagocytosis was stopped by adding cold (4°C) RPMI 1640 and extracellular S. aureus were removed by washing the suspension in RPMI 1640. The pellets were disrupted in sterile water containing 0.01% bovine serum albumin (BSA) by vigorously vortexing to release the intracellular bacteria in the lysates. The lysates containing bacteria were plated at serial dilutions on mannitol agar plates. The plates were incubated at 37°C for a day or two and the number of colonies was determined.
Hydrogen-peroxide production
Activation of leukocytes by inflammatory stimuli results in the local release of ROS and induces hydrogen-peroxide (H2O2) production. Studies in animals support a critical role for phagocyte oxidative burst in controlling S. aureus infection.Citation35–Citation37 Therefore, the quantification of H2O2 in this experimental setup was quite relevant.
So, after time-dependent phagocytosis, supernatants were collected and cell lysates were prepared from the pellet. H2O2 assay of the supernatants and lysates was performed according to the an earlier described method, with slight modification.Citation51 Briefly, 70 μL of supernatant or lysate, 20 μL horseradish peroxidase (500 μg/mL), 70 μL of phenol red (500 μg/mL), and 40 μL medium were added to each well of the microtiter plates and allowed to incubate for 2 hours at 37°C. The reaction was stopped by adding 25 μL of 2 N sodium hydroxide (NaOH) and the absorbance reading was taken at 620 nm. The control set received 40 μL of HBSS in place of the supernatant/lysate. A standard H2O2 curve was plotted and the H2O2 release from the supernatants and lysates evaluated and expressed in terms of μM/106 cells.
Assay for determination of superoxide anion
Because intracellular killing is dependent on oxidants, it was expected that superoxide anion production also contributes to S. aureus oxidant-dependent killing mechanisms. Since superoxide anion production has been implicated in many physiological and pathological processes, including host innate immune and inflammatory responses to pathogens, we were also interested in determining the amount of superoxide anion released in our experimental setup.
Superoxide anion release assay measures the change in color of cytochrome C (cytC), when reduced by O2− released from the stimulated macrophages. Cell-free supernatants recovered from S. aureus-stimulated macrophages in the presence or absence of antibiotics and CQ alone or in combination were incubated in the presence of cytochrome C (100 μL at 2 mg/mL in HBSS). The reaction was terminated by placing the tubes in ice for 5 minutes. The production of superoxide anion was monitored spectrophotometrically using a UV-1800 UV–VIS (ultraviolet–visible) spectrophotometer (Shimadzu Corporation, Tokyo, Japan) at 550 nm in reference to the blank. Based on the extinction coefficient of (reduced–oxidized CytC), OD =21 × 103 M−1cm−1 and because the vertical light path passing through 100 μL CytC was 3 mm, the absorbance value could be converted to nmol of superoxide anion based on the following formula: nmol of superoxide anion = (mean absorbance at 550 nm × 15.87).Citation52
Assay for quantification of nitric-oxide production
Nitric-oxide (NO) release was determined by Griess assay. Thioglycolate-elicited mouse peritoneal macrophages in serum-free RPMI 1640 medium were mixed with S. aureus and the mixture was incubated in the presence or absence of the anti-toll-like receptor (TLR)-2 antibody for different times at 37°C with 5% CO2 before being centrifuged. Supernatant or lysate (50 μL) was incubated separately in 40 μM Tris (pH 7.9) containing 40 μM of the reduced form of b-nicotinamide adenine dinucleotide phosphate (NADPH), 40 μM flavine adenine dinucleotide, and 0.05 U/mL nitrate reductase at 37°C for 15 minutes. Reduced samples were incubated with an equal volume of Griess reagent consisting of sulphanilamide (0.25% [w/v]) and N-1-naphthylethylene-diamine (0.025% [w/v]) for 10 minutes. Absorbance was measured at 550 nm with the UV-1800 UV–VIS spectrophotometer (Shimadzu Corporation). The total nitrate/nitrite concentration was determined by comparison to a reduced sodium nitrate (NaNO3) standard curve.Citation53
Determination of antioxidant enzyme activity in supernatants and lysates after time-dependent phagocytosis of S. aureus in the presence or absence of antibiotics and chloroquine alone or in combination
Assay of catalase enzyme activity
Typically, S. aureus-stimulated leukocytes produce proinflammatory cytokines, which trigger ROS production in the tissues through NADPH oxidase activation. Thus, the level of antioxidant enzymes or their activity displays the intracellular complex mechanisms of the host’s defense. Therefore, to determine the activity of these antioxidant enzymes in neutralizing the ROS molecules produced, we estimated the antioxidant enzyme activity in the supernatant or cell-free lysate of macrophages after infection in the presence or absence of CQ or antibiotics. Catalase enzyme activity in the supernatants or cell-free lysates was determined spectrophotometrically by measuring the decrease in H2O2 concentration at 240 nm. At time 0, 10 μL of supernatant or cell-free lysate was added separately to 2.89 mL of potassium phosphate buffer (pH 7.4) in a quartz cuvette. To this, 0.1 mL of 300 mM H2O2 was added and absorbance was measured at 240 nm for 5 minutes at 1-minute intervals. Catalase activity was expressed in terms of mmol/min mg protein.Citation20–Citation22
Assay of superoxide dismutase enzyme activity
In addition to blocking phagolysosomal acidification, subsequent treatment of macrophages with antibiotics might regulate intracellular ROS and can also protect bacteria from oxidative stress by modulating bacterial superoxide dismutase (SOD) expression or catalase activity. So estimation of SOD enzyme activity in this experimental setup was deemed helpful to get an idea of ROS level and is being scavenged by antioxidant enzymes.
Supernatant or cell-free lysate (100 μL) was mixed separately with 1.5 mL of a Tris-ethylenediaminetetraacetic acid (EDTA)-hydrochloric acid (HCl) buffer (pH 8.5), then 100 μL of 7.2 mmol/L pyrogallol was added and the reaction mix ture was incubated at 25°C for 10 minutes. The reaction was terminated by the addition of 50 μL of 1 M HCl and measured at 420 nm. One unit was determined to be the amount of enzyme that inhibited the oxidation of pyrogallol by 50%. The activity was expressed as U/mg protein.Citation20–Citation22
Assay of reduced glutathione content
SOD, catalase, and glutathione (GSH) enzymes are important scavengers of the superoxide anion and hydrogen peroxide. These enzymes prevent the generation of the hydroxyl radical and protect the cellular constituents from oxidative damage. SOD rapidly dismutates superoxide anion to H2O2, which is further degraded by catalase into water and oxygen. It was demonstrated that a decrease in the concentration of cellular GSH indicates oxidative damage due to stress or inflammation.Citation54 So estimation of GSH level was also rational in this setup.
Reduced GSH content (as acid-soluble sulfhydryl) was estimated by its reaction with 5,5′-dithiobis-(2-nitrobenzoic acid) (DTNB; Ellman’s reagent) following the method of Sedlak and Lindsay with some modifications.Citation55 An amount of the sample (0.3 mL) was mixed with 0.3 mL 10% trichloroacetic acid (TCA) followed by vortexing. Then the mixture was centrifuged at 5,000 rpm for 10 minutes at 4°C. To 250 μL of the resultant mixture, supernatant was added along with 500 μL 0.8 M Tris-HCl followed by addition of 25 μL DTNB. The absorbance was measured at 412 nm using a UV–VIS spectrophotometer to determine GSH content. The values were expressed as nmol of GSH per mg protein.
Determination of lipid peroxidation level
Lipid peroxidation (LPO) of both the supernatants and lysates were determined as thiobarbituric acid (TBA)-reactive substances. In brief, the supernatant or lysate was mixed with TCA-TBA-HCl reagent, mixed thoroughly, and heated for 20 minutes at 80°C. The tubes containing the samples were then cooled to room temperature. The absorbance of the pink chromogen present in the clear supernatant after centrifugation at 1,200× g for 10 minutes at room temperature was measured at 532 nm using a UV–VIS spectrophotometer. Tetraethoxypropane was used as the standard. The values were expressed as nmol of TBA-reactive substances per mg protein.Citation20
Assay of bactericidal enzyme (lysozyme) activity
Through lysosomal degradation and response to secondary ligands, macrophages are able to extend and maximize the inflammatory response. The importance of lysosomal degradation and bactericidal enzymes like lysozymes in the immune response is supported by numerous studies. Pathogens evolving to inhibit lysosomal fusion, acidification, and lysosomal enzymes are necessary to release additional ligands and enhance innate signaling. Therefore, the estimation of lysozyme enzyme activity in this study was quite rational.
The supernatants and cell-free lysates obtained in the phagocytosis assay were used to estimate the lysozyme content after varying phagocytic times. Lysozyme enzyme activity was estimated according to the method described by Colowick et al.Citation56 A suspension of Micrococcus lysodeikticus in 0.15 M potassium phosphate buffer (pH 6.2) was prepared and its optical density was fixed at 0.6–0.7 at 450 nm. An amount of this suspension (900 μL) was mixed with 50 μL of 1% Triton™ X-100 and the reaction was started by adding 50 μL supernatant or cell-free lysate. The decrease in optical density was recorded at 450 nm as a function of time. The change in absorbance for the first minute only was used to determine the rate of the reaction. One unit of enzyme was defined as the amount of enzyme that produced a decrease in absorbance of 0.001/min at 450 nm.
Assay for protein estimation of the sample
The protein content of the supernatants and lysates was measured by previously described method.Citation57
Tumor necrosis factor alpha and interferon gamma enzyme-linked immunosorbent assays
Murine peritoneal macrophages (5×106 cells/mL) were mixed with S. aureus (5×106 CFU/mL) in a cell to bacterium ratio of 1:1Citation58,Citation59 in RPMI-FBS (5%) and incubated at 37°C in a cell-culture incubator for various times in the presence and absence of antibiotics and CQ either alone or in combination. After incubation, the cell-culture supernatants were collected and stored at −70°C prior to analysis. Supernatants from different groups were normalized to the protein content by Bradford protein assay before the assay and levels of TNF-α and IFNγ were determined. Production was assayed using enzyme-linked immunosorbent assay (ELISA) kits according to the instructions provided by the manufacturer (RayBiotech, Inc., Norcross, GA, USA) in an ELISA reader (Bio-Rad Laboratories Inc., Hercules, CA, USA).
Statistical analysis
Isolated peritoneal macrophages from mice were pooled together to obtain the requisite amount of macrophages (5×106 cells/mL) and the different parameters were measured. This was repeated three times for each parameter (eg, H2O2 production) then the mean value of the triplicate experiments was taken for calculation. Data were expressed as mean ± standard deviation. Means were compared between groups using analysis of variance. A P-value <0.05 was considered significant.
Results
Effect of chloroquine and antibiotic treatment either alone or in combination on phagocytosis of S. aureus by peritoneal macrophages
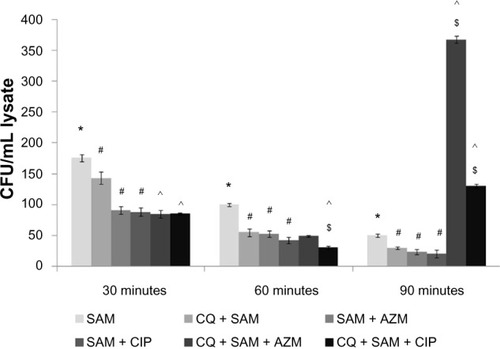
Control macrophages were readily able to phagocytose S. aureus. After 90 minutes of incubation the number of viable bacteria within macrophages declined. However, the number of bacteria phagocytosed by CQ-treated macrophages appeared to be lower than that observed for macrophages not treated with CQ at the different incubation times. Overall, the experimental results indicate that CQ treatment leads to a reduction in the uptake of S. aureus cells by macrophages, presumably by interfering with phagocytosis. CIP and AZM do not affect the viability of macrophages (data not shown), but almost immediately after being applied to infected macrophages inhibits S. aureus growth and replication (data not shown).
There were statistically significant differences between the number of viable bacteria recovered from CQ plus antibiotic-treated macrophages before S. aureus infection and that from the CQ-pretreated plus S. aureus-infected macrophages at 30 and 60 minutes postinfection. However, the number of bacteria surviving intracellularly in the CQ plus AZM-treated macrophages before S. aureus infection was greater than that in the CQ plus CIP-treated macrophages before infection at 90 minutes of incubation when compared to CQ-treated S. aureus-infected macrophages ().
Figure 1 Effect of chloroquine (CQ) and antibiotic treatment on intracellular killing assay on Staphylococcus aureus-infected murine peritoneal macrophages. Intracellular killing assay was performed on the lysate after time-dependent phagocytosis in the presence or absence of CQ or antibiotics.
Notes: Intracellular killing assay is expressed in terms of the colony-forming units (CFU)/ml lysate. Results are presented as mean ± standard deviation of three independent experiments. *significant difference with respect to CM; #significant difference with respect to SAM; ^significant difference with respect to chloroquine (CQ) plus S. aureus-infected macrophages at the P<0.05 level of significance; $significant difference with respect to either ciprofloxacin (CIP) or azithromycin (AZM) plus S. aureus-infected macrophages.
Abbreviations: CM, control macrophages; CQ + SAM, chloroquine-pretreated + S. aureus-infected macrophages; CQ + SAM + AZM, chloroquine-pretreated macrophages infected with S. aureus and then treated with azithromycin; CQ + SAM + CIP, chloroquine-pretreated macrophages infected with S. aureus and then treated with ciprofloxacin; SAM, S. aureus-infected macrophages; SAM + AZM, S. aureus-infected macrophages treated with azithromycin; SAM + CIP, S. aureus-infected macrophages treated with ciprofloxacin.
Effect of chloroquine and antibiotic treatment on S. aureus infection-induced hydrogen-peroxide production in the lysate and supernatant of murine peritoneal macrophages
The amount of H2O2 released in the lysate () or supernatant () was significantly higher (P<0.05) in S. aureus-infected macrophages than in control macrophages at 30, 60, and 90 minutes after S. aureus infection. There was a statistically significant increase in the amount of H2O2 produced (P<0.05) in CQ-pretreated macrophages at 30 and 60 minutes post-incubation in both supernatant and lysate but there was a decrease at 90 minutes in the supernatant when compared with untreated S. aureus-infected macrophages ().
Figure 2 Effect of chloroquine (CQ) and antibiotic treatment on Staphylococcus aureus infection-induced alteration in H2O2 release in the lysate and supernatant in murine peritoneal macrophages. The lysate (A) and supernatant (sup) (B) recovered after time-dependent phagocytosis in the presence or absence of chloroquine or antibiotics were used to determine the H2O2 release.
Notes: H2O2 release is expressed in terms of μM/106 cells. Results are presented as mean ± standard deviation of three independent experiments. *significant difference with respect to CM; #significant difference with respect to SAM; ^significant difference with respect to CQ plus S. aureus-infected macrophages at the P<0.05 level of significance; $significant difference with respect to either ciprofloxacin (CIP) or azithromycin (AZM) plus S. aureus-infected macrophages.
Abbreviations: CM, control macrophages; CQ + SAM, chloroquine-pretreated + S. aureus-infected macrophages; CQ + SAM + AZM, chloroquine-pretreated macrophages infected with S. aureus and then treated with azithromycin; CQ + SAM + CIP, chloroquine-pretreated macrophages infected with S. aureus and then treated with ciprofloxacin; SAM, S. aureus-infected macrophages; SAM + AZM, S. aureus-infected macrophages treated with azithromycin; SAM + CIP, S. aureus-infected macrophages treated with ciprofloxacin.
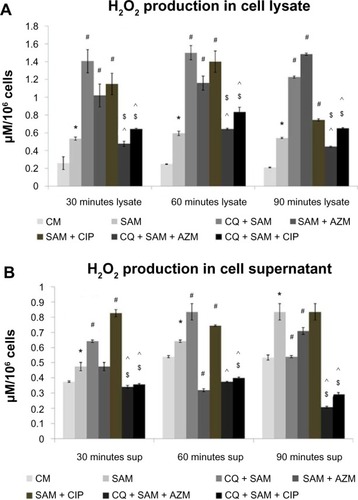
Macrophages pretreated either with CIP or AZM alone before S. aureus infection also showed increased production of H2O2 in the lysate in comparison to the amount released by untreated S. aureus-infected macrophages. However, administration of either CIP or AZM to CQ-pretreated macrophages before S. aureus infection showed reduced H2O2 release both in the supernatant and lysate when compared with CQ plus S. aureus-infected macrophages (). The H2O2 release was also lower in CQ-pretreated macrophages were incubated with CIP or AZM before S. aureus infection than in CIP- or AZM-treated S. aureus-infected macrophages at 30, 60, and 90 minutes in the cell lysate ().
Effect of CQ and antibiotic treatment on S. aureus infection-induced superoxide anion production in the lysate and supernatant by murine peritoneal macrophages
The amount of superoxide anion (nmol/106 cells) released in the supernatant and lysates was also estimated. The increase in the O2− released in lysate () and supernatant () after different times of phagocytosis in the presence of S. aureus infection was found to be significant (P<0.05) when compared with the O2− release of noninfected control macrophages. A statistically significant increase in the amount of O2− released both in the supernatant and lysate was observed (P<0.05) in CQ-pretreated macrophages before infection, when compared with untreated S. aureus-infected macrophages at 30, 60, and 90 minutes post-incubation ().
Figure 3 Effect of chloroquine (CQ) and antibiotic treatment on Staphylococcus aureus infection in the lysate and supernatant of murine peritoneal macrophages. The lysate (A) and supernatant (sup) (B) after time-dependent phagocytosis in the presence or absence of CQ or antibiotics were used to determine the release of superoxide anion.
Notes: Superoxide anion release is expressed in terms of nM/106 cells. Results are presented as mean ± standard deviation of three independent experiments. *Significant difference with respect to CM; #significant difference with respect to SAM; ^significant difference with respect to CQ plus S. aureus-infected macrophages at the P<0.05 level of significance; $significant difference with respect to either ciprofloxacin (CIP) or azithromycin (AZM) plus S. aureus-infected macrophages.
Abbreviations: CM, control macrophages; CQ + SAM, chloroquine-pretreated plus S. aureus-infected macrophages; CQ + SAM + AZM, chloroquine-pretreated macrophages infected with S. aureus and then treated with azithromycin; CQ + SAM + CIP, chloroquine-pretreated macrophages infected with S. aureus and then treated with ciprofloxacin; SAM, S. aureus-infected macrophages; SAM + AZM, S. aureus-infected macrophages treated with azithromycin; SAM + CIP, S. aureus-infected macrophages treated with ciprofloxacin.
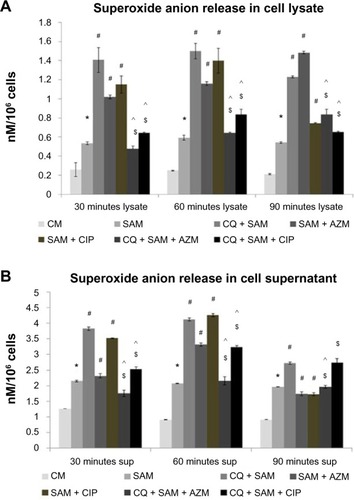
Treatment of macrophages with CIP and AZM before S. aureus infection resulted in an increased amount of O2− being released into the supernatant and lysate at 30 and 60 minutes but a decreased amount at 90 minutes in cell supernatant post-incubation when compared with untreated S. aureus-infected macrophages (). The amount of O2− released significantly decreased in 30 and 60 minutes in both the supernatant and lysate in CQ-pretreated and AZM/CIP-treated S. aureus macrophages but remained same at 90 minutes in supernatant when compared with CQ plus S. aureus-infected macrophages after phagocytosis. O2− release was significantly lower at 30, 60, and 90 minutes in the cell lysate and at 30 and 60 minutes in cell supernatant when CQ-pretreated macrophages were incubated with CIP or AZM before S. aureus infection compared with CIP- or AZM-treated S. aureus-infected macrophages at 30, 60, and 90 minutes in the cell lysate ().
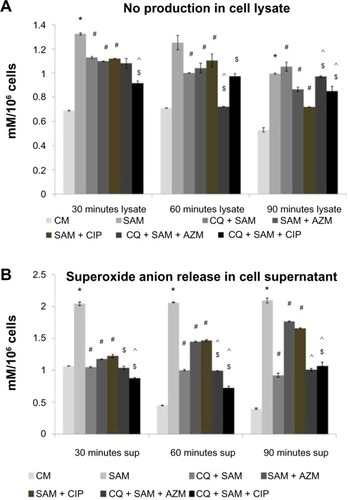
Effect of chloroquine and antibiotic treatment on S. aureus infection-induced nitric-oxide production in the lysate and supernatant of murine peritoneal macrophages
CQ inhibits NO production when added during the stimulation of peritoneal macrophages with S. aureus. A statistically significant decrease in the amount of NO released both in the lysate () and supernatant () was observed (P<0.05) in CQ-pretreated macrophages before infection, compared with untreated S. aureus-infected macrophages at 30 and 60 minutes post-incubation. The amount of NO release decreased in the case of macrophages treated with AZM or CIP alone before S. aureus infection compared with untreated S. aureus-infected macrophages (). The NO release was significantly lower at 30 and 60 minutes in the supernatant and at 60 minutes in the cell lysate when CQ-pretreated macrophages were incubated with CIP or AZM before S. aureus infection, compared with CIP- or AZM-treated S. aureus-infected macrophages.
Figure 4 Effect of chloroquine (CQ) and antibiotic treatment on Staphylococcus aureus infection induced NO release in the lysate and supernatant in murine peritoneal macrophages. The lysate (A) and supernatant (B) recovered after time-dependent phagocytosis in the presence or absence of CQ or antibiotics were used to determine the nitric-oxide release by Griess assay.
Notes: nitric-oxide release is expressed in terms of mM/106 cells. Results are presented as mean ± standard deviation of three independent experiments. *Significant difference with respect to CM; #significant difference with respect to SAM; ^significant difference with respect to CQ plus S. aureus-infected macrophages at the P<0.05 level of significance; $significant difference with respect to either ciprofloxacin (CIP) or azithromycin (AZM) plus S. aureus-infected macrophages.
Abbreviations: CM, control macrophages; CQ + SAM, chloroquine-pretreated plus S. aureus-infected macrophages; CQ + SAM + AZM, chloroquine-pretreated macrophages infected with S. aureus and then treated with azithromycin; CQ + SAM + CIP, chloroquine-pretreated macrophages infected with S. aureus and then treated with ciprofloxacin; SAM, S. aureus-infected macrophages; SAM + AZM, S. aureus-infected macrophages treated with azithromycin; SAM + CIP, S. aureus-infected macrophages treated with ciprofloxacin.
Effect of chloroquine and antibiotic treatment on S. aureus infection-induced alteration in the catalase enzyme activity in the supernatant and lysate of murine peritoneal macrophages
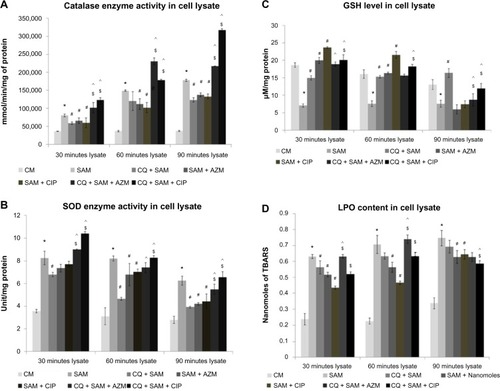
and show a marked increase in the catalase enzyme activity in the supernatant and lysate of peritoneal macrophages infected with S. aureus after 30, 60, and 90 minutes of phagocytosis compared with the noninfected control group. A significant decrease in catalase enzyme activity was observed in the supernatants of CQ-treated murine peritoneal macrophages before infection compared with in those of the CQ non-treated infected group. Treatment of macrophages with either CIP or AZM before S. aureus infection decreased the amount of catalase enzyme activity in the supernatant compared with the amount found in untreated S. aureus-infected macrophages (). However treatment of S. aureus with AZM increased catalase activity both in the supernatant and lysate in comparison to treatment with CIP but the catalase production was less compared with that in untreated S. aureus-infected macrophages. This increase in the catalase enzyme activity was even more prominent when CQ-pretreated macrophages were incubated with CIP rather than with AZM before S. aureus infection in comparison to CQ plus S. aureus-infected macrophages at 30, 60, and 90 minutes in both supernatant and lysate ( and ).
Figure 5 Effect of chloroquine (CQ) and antibiotic treatment on Staphylococcus aureus infection-induced alteration in catalase, superoxide dismutase (SOD) enzyme activity, glutathione (GSH) level, and lipid peroxidation (LPO) content in the supernatant in murine peritoneal macrophages. The supernatant (sup) was recovered after time-dependent phagocytosis in the presence or absence of CQ or antibiotic treatment, as described in the “Methods and materials” section, and were used to determine the catalase (A), SOD enzyme activity (B), GSH level (C), and LPO content (D) in the supernatant.
Notes: Catalase enzyme activity was measured in the presence of 15 μmoles of H2O2/mL of phosphate buffer and is expressed in terms of mmol/min mg protein (A); SOD activity is expressed in terms of SOD units/mg protein (B); GSH content is expressed in terms of μM/mg protein (C); and lPO content is expressed in terms of nmol of thiobarbituric acid-reactive substances (TBARS) (D). Results are shown as mean ± standard deviation of three independent experiments. *significant difference with respect to CM; #significant difference with respect to SAM; ^significant difference with respect to CQ plus S. aureus-infected macrophages at the P<0.05 level of significance; $significant difference with respect to either ciprofloxacin (CIP) or azithromycin (AZM) plus S. aureus-infected macrophages.
Abbreviations: CM, control macrophages; CQ + SAM, chloroquine-pretreated plus S. aureus-infected macrophages; CQ + SAM + AZM, chloroquine-pretreated macrophages infected with S. aureus and then treated with azithromycin; CQ + SAM + CIP, chloroquine-pretreated macrophages infected with S. aureus and then treated with ciprofloxacin; SAM, S. aureus-infected macrophages; SAM + AZM, S. aureus-infected macrophages treated with azithromycin; SAM + CIP, S. aureus-infected macrophages treated with ciprofloxacin.
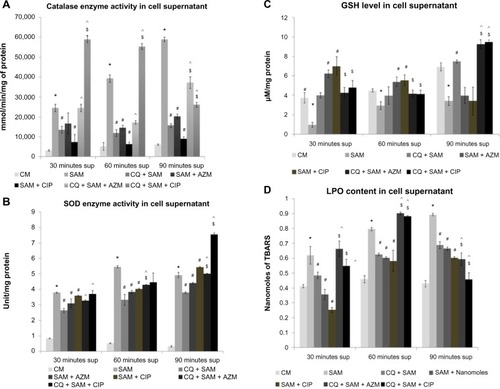
Figure 6 Effect of chloroquine (CQ) and antibiotic treatment on Staphylococcus aureus infection-induced alteration in catalase, superoxide dismutase (SOD) enzyme activity, glutathione (GSH) level, and lipid peroxidation (LPO) content in the lysate in murine peritoneal macrophages. The cell-free lysate was recovered after time-dependent phagocytosis in the presence or absence of CQ or antibiotic treatment, as described in the “Methods and materials” section, and were used to determine the catalase (A), SOD enzyme activity (B), GSH level (C), and LPO content (D) in the lysate.
Notes: Catalase enzyme activity was measured in the presence of 15 μmoles of H2O2/ml of phosphate buffer and is expressed in terms of millimoles/minute mg protein (A); SOD activity is expressed in terms of SOD units/mg protein (B); GSH content is expressed in terms of μM/mg protein (C); and LPO content is expressed in terms of nmol of thiobarbituric acid-reactive substances (TBARS) (D). Results are shown as mean ± standard deviation of three independent experiments. *significant difference with respect to CM; #significant difference with respect to SAM; ^significant difference with respect to CQ plus S. aureus-infected macrophages at the P<0.05 level of significance; $significant difference with respect to either ciprofloxacin (CIP) or azithromycin (AZM) plus S. aureus-infected macrophages.
Abbreviations: CM, control macrophages; CQ + SAM, chloroquine-pretreated plus S. aureus-infected macrophages; CQ + SAM + AZM, chloroquine-pretreated macrophages infected with S. aureus and then treated with azithromycin; CQ + SAM + CIP, chloroquine-pretreated macrophages infected with S. aureus and then treated with ciprofloxacin; SAM, S. aureus-infected macrophages; SAM + AZM, S. aureus-infected macrophages treated with azithromycin; SAM + CIP, S. aureus-infected macrophages treated with ciprofloxacin.
Effect of chloroquine and antibiotic treatment on S. aureus infection-induced alteration in the superoxide dismutase enzyme activity in the supernatant and lysate of murine peritoneal macrophages
and show that there was a significant increase in the SOD enzyme activity in the supernatant and lysate of peritoneal macrophages infected with S. aureus after 30, 60, and 90 minutes of phagocytosis compared with in the noninfected control group (P<0.05). A significant decrease in the SOD enzyme activity was observed in the supernatants and lysates of CQ-treated murine peritoneal macrophages before infection compared with in the CQ non-treated infected group. Treatment of macrophages with either CIP or AZM alone before S. aureus infection resulted in decreased SOD enzyme activity at all time intervals in the lysate and only at 30 and 60 minutes in the supernatant when compared with untreated S. aureus-infected macrophages. The activity of SOD enzyme was found to increase at 30, 60, and 90 minutes in both supernatant and lysate when CQ-pretreated macrophages were incubated with CIP or AZM before S. aureus infection in comparison to in CQ-pretreated S. aureus-infected macrophages, however the enzyme activity was found to increase with time in the supernatant of CQ- and CIP-pretreated S. aureus-infected macrophages ().
Effect of chloroquine and antibiotic treatment on S. aureus infection-induced alteration in the glutathione content in the supernatant and lysate of murine peritoneal macrophages
The GSH content in the supernatant () and lysate () was significantly lower (P<0.05) in S. aureus-infected macrophages than in control macrophages at 30, 60, and 90 minutes after S. aureus infection. A statistically significant increase in the amount of GSH in the lysate was observed (P<0.05) in CQ-pretreated macrophages before infection at 30, 60, 90 minutes post-incubation, when compared with in untreated S. aureus-infected macrophages ().
Macrophages pretreated with AZM or CIP alone before S. aureus infection showed an increased amount of GSH content at 30 and 60 minutes in the supernatant and lysate but a decreased amount at 90 minutes post-incubation when compared with the amount of GSH released by untreated S. aureus-infected macrophages. However, administration of CIP or AZM to the CQ-pretreated macrophages before S. aureus infection increased the GSH content in the supernatant at 90 minutes compared with in the CQ plus S. aureus-infected macrophages (). Compared with CIP- or AZM-treated S. aureus-infected macrophages, there was less GSH production observed in both the supernatant and lysate at 30 and 60 minutes but increased GSH production at 90 minutes in the cell supernatant in CQ-pretreated macrophages incubated with CIP or AZM before S. aureus infection.
Effect of chloroquine and antibiotic treatment on S. aureus infection-induced alteration in the lipid peroxidation level in the supernatant and lysate of murine peritoneal macrophages
The amount of LPO released in the supernatant () and lysate () was significantly higher (P<0.05) in S. aureus-infected macrophages than in control macrophages at 30, 60, and 90 minutes after S. aureus infection. A statistically significant decrease in LPO production in the supernatant and lysate was observed (P<0.05) in CQ-pretreated macrophages at 30, 60, and 90 minutes post-incubation in the cell supernatant as well as at 30 minutes in lysate when compared with untreated S. aureus-infected macrophages ( and ).
Macrophages pretreated either with CIP or AZM alone before S. aureus infection also showed decreased production of LPO in the supernatant and lysate in comparison with that released in untreated S. aureus-infected macrophages. However, administration of either CIP or AZM to the CQ-pretreated macrophages before S. aureus infection increased LPO release, both in supernatant and lysate at 30 and 60 minutes when compared with CQ plus S. aureus-infected macrophages ( and ). Compared with in CIP- or AZM-treated S. aureus-infected macrophages, the LPO released was significantly higher after 30 and 60 minutes in supernatant and lysate, respectively, in CQ-pretreated macrophages incubated with CIP or AZM before S. aureus infection ( and ).
Effect of chloroquine and antibiotic treatment on S. aureus infection-induced alteration in the lysozyme enzyme activity in the supernatant and lysate of murine peritoneal macrophages
shows there was a significant increase in the lysozyme enzyme activity in the supernatant and lysate of peritoneal macrophages infected with S. aureus after 30, 60, and 90 minutes of phagocytosis compared with in the noninfected control group (P<0.05). A significant decrease in the lysozyme enzyme activity was observed in the supernatants and lysates of CQ-treated murine peritoneal macrophages before infection compared with in the non-CQ-treated infected group. Treatment of macrophages with either CIP or AZM alone before S. aureus infection also resulted in a decreased amount of lysozyme enzyme activity in the supernatant and lysate compared with in the untreated S. aureus-infected macrophages (). However, the lysozyme enzyme activity was restored significantly in the lysate at 30 and 60 minutes when CQ-pretreated macrophages were incubated with either CIP or AZM before being infected with S. aureus in comparison with in CQ plus S. aureus-infected macrophages (). The lysozyme released was significantly higher at 30 and 60 minutes in supernatant and lysate, respectively, when CQ-pretreated macrophages were incubated with CIP or AZM before being infected with S. aureus, compared with when CIP- or AZM-treated S. aureus-infected macrophages were not pretreated with CQ.
Table 1 effect of chloroquine (CQ) and antibiotic treatment on S. aureus infection-induced lysozyme enzyme activity in the supernatant and lysate by murine peritoneal macrophages
Effect of chloroquine and antibiotic treatment on S. aureus infection-induced interferon gamma and tumor necrosis factor alpha production in murine peritoneal macrophages
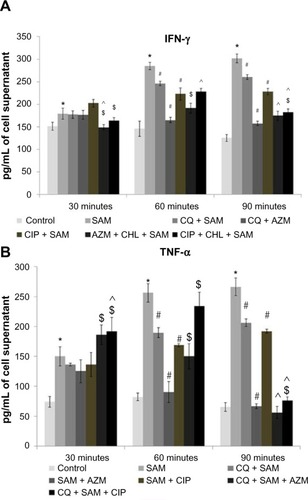
There was a significant (P<0.05) gradual decrease in IFN-γ () and TNF-α () levels in the media (culture supernatant) when phagocytic time increased in the presence of CQ at 60 and 90 minutes compared with the cytokine production after time-dependent phagocytosis in CQ non-treated infected macrophages. Macrophages pretreated either with CIP or AZM alone before S. aureus infection also showed reduced levels of TNF-α and IFN-γ at both 60 and 90 mins in the supernatant in comparison to that released by the untreated S. aureus-infeccted macrophages. However, the levels of TNF-α and IFN-γ decreased significantly in the medium at 90 minutes when CQ-pretreated macrophages were incubated with either CIP or AZM before being infected with S. aureus in comparison to CQ plus S. aureus-infected macrophages. The level of TNF-α decreased significantly in the medium at 60 and 90 minutes when CQ-pretreated macrophages were incubated with either CIP or AZM before being infected with S. aureus in comparison to CQ plus S. aureus-infected macrophages ().
Figure 7 levels of (A) interferon gamma (IFN-γ) and (B) tumor necrosis factor alpha (TNF-α) in the supernatants collected from Staphylococcus aureus-infected macrophages in the presence or absence of chloroquine (CQ) or antibiotics were determined utilizing enzyme-linked immunosorbent assay according to the manufacturer’s recommendations.
Notes: Results from triplicate experiments are presented. *significant difference with respect to CM; #significant difference with respect to SAM; ^significant difference with respect to CQ plus S. aureus-infected macrophages at the P<0.05 level of significance; $significant difference with respect to either ciprofloxacin (CIP) or azithromycin (AZM) plus S. aureus-infected macrophages.
Abbreviations: CM, control macrophages; CQ + SAM, chloroquine-pretreated plus S. aureus-infected macrophages; CQ + SAM + AZM, chloroquine-pretreated macrophages infected with S. aureus and then treated with azithromycin; CQ + SAM + CIP, chloroquine-pretreated macrophages infected with S. aureus and then treated with ciprofloxacin; SAM, S. aureus-infected macrophages; SAM + AZM, S. aureus-infected macrophages treated with azithromycin; SAM + CIP, S. aureus-infected macrophages treated with ciprofloxacin.
Discussion
The intracellular persistence of S. aureus within macrophages, especially the antibiotic-resistant types, has posed serious problems and difficulties in the treatment of several diseases. Moreover, while it has been shown that cells take up antibiotics with known intracellular bactericidal activity, that they no longer confer protection in vitro. The possible reasons for this discrepancy are numerous: 1) the degree of plasma-membrane permeability may determine the concentration gradients of the antibiotic inside and outside of the cell; 2) antibiotics and bacteria may segregate subcellularly in diverse intracellular compartments, because of the dynamic exposure of the infectious agent to the unbound antibiotic drug fraction at the relevant effect site, resulting in a pharmacodynamic effect in in vivo conditions, whereas such a pharmacodynamic effect remains absent under the static in vitro conditions; 3) antibiotics may be inactivated by the intracellular milieu; and 4) differences in the physiological state of the internalized pathogen (resting vs dividing) can modulate the susceptibility to antibiotics.Citation60 Another complicating factor is the extracellular multiplication of the surface-associated bacteria or those released from lysed infected cells, since these organisms would be subject to repeated endocytic events.
Efforts to obviate this difficulty by adding appropriate antibiotics and by restricting the entry of viable staphylococci associated with the cell’s surface in vitro require a novel combination of drugs involving an efficacious antibiotic with intracellular bactericidal activity and an alkalinization of the intra-lysosomal pH by using CQ to enhance the uptake of antibiotics. Thus, the aim of our study was to formulate an efficacious combination using CQ (for alkalinization of the lysosomal pH) to observe the effect of CIP and AZM on the killing of staphylococcal cells in isolated murine peritoneal macrophages and staphylococcal cells subsequent lysis to reduce the pathogen load within the phagocytic cells depending on the uptake of antibiotics. We chose mouse peritoneal macrophages as we could investigate phagocytosis within 2 hours of isolation, compared with in the 1 week required for differentiation of human monocyte-derived macrophages.Citation61,Citation62 Moreover, the peritoneal macrophages used in the study were elicited with thioglycolate broth, which did not potentially affect intracel lular killing, as evident in our earlier studies.Citation15–Citation17 Therefore, the inclusion of data set by using untreated or non-elicited macrophages by thioglycolate broth, such as alveolar macrophages or splenic macrophages, was not essential in our study. When testing the CQ as a modulator of antibiotic uptake, several assays were performed: intracellular killing, ROS production, antioxidant enzymes, lysozyme release, and finally measurement of cytokines released by host cells in response to all these treatments before S. aureus infection. The rationale for performing these assays has already been mentioned in the “Materials and methods” section. These assays are very much linked and do not stand individually.
CQ (which neutralizes lysosomal pH) in the presence of CIP or AZM strikingly increased the killing of intracellularly surviving S. aureus, as evident from the number of bacterial CFU (). It has been reported that pH inside macrophage lysosomes seems to be 4.7 to 4.8.Citation63 It was evident from both the mentioned reference and from our own study that CQ was taken up by macrophages most rapidly after 30 minutes when macrophages were exposed to 100 μM CQ.Citation63 It has been reported that the optimal pH for AZM accumulation in PMN is 8.7.Citation64 Here, it can be suggested that AZM accumulated at its maximum level within 60 minutes of CQ treatment. In mouse peritoneal macrophages that had endocytosed fluorescein isothiocyanate-labeled dextran, Ohkuma and Poole determined that treatment with 100 μM CQ raised the pH to 6.4.Citation63 It was shown that the elevated pH value can remain stable for as long as 2 hours.Citation65 CQ causes a remarkable pH increase, even at 1 μM concentration, and it has also been reported that a medium pH does not have any obvious effect on intra-lysosomal pH.Citation63 Intra-lysosomal pH may play an important role in the antimicrobial activity of macrophages. It is worth noting that the kinetics of the transient pH rise coincides with the phagocytic activity of the macrophages, such as respiratory burst with the degranulation of vesicles containing lysosomal enzymes. We did not actually quantify the pH in the phagosomes following CQ treatment;Citation66 however, there are reports that can support our hypothesis in relation to the concentration of CQ and time of incubation used in our study. In another study, the treatment of human macrophages with 100 μM CQ raised the pH of C. neoformans-laden phagosomes to 7.3.Citation67 Indeed, in our study, the lysosomal bactericidal enzyme lysozyme, which requires an acid pH, was inhibited in the CQ-pretreated macrophages. However, CQ in the presence of AZM or CIP significantly increased the release of lysozyme for the intracellular killing of S. aureus during early infection. CQ is a lysosomotropic agent that prevents endosomal acidification. It accumulates inside the acidic parts of the cell, including in the endosomes and lysosomes. This accumulation leads to the inhibition of lysosomal enzymes, which require an acidic pH, and prevents the fusion of endosomes and lysosomes. The antirheumatic activity of CQ is thought to be related to its effects on lysosomal functions and protein processing.Citation68 Based on the similar cationic and lysosomal accumulating properties, as well as interaction with membrane phospholipids, the action of CQ and AZM on a number of intracellular processes related to the killing of intracellular bacteria has been investigated.Citation69 The present study indicates that due to AZM uptake (during the process of alkalinizing the lysosomal pH) or lysozyme release during early infection presumably comes from the lysosomal compart ment as a result of phagocytosis-induced degranulation. The alkalinization-induced killing of S. aureus by AZM causes more bactericidal lysozyme release during early infection and alters both the uptake and release of AZM in murine macrophages. It has also been reported that the ingestion of zymosan by human PMN modestly enhances the release of AZM. Agents like CQ are also reported to increase the efflux of AZM.Citation70
CQ in the presence of antibiotics has been reported to improve the killing of bacteria where intra-macrophage survival plays a key step in disease pathogenesis.Citation71 It has also been demonstrated that CQ inhibits intra-macrophage replication of Crohn’s disease-associated Escherichia coli and enhances the antimicrobial effects of the antibiotics doxycycline and CIP.Citation72 Several fluoroquinolone antibiotics are known to penetrate host cell membranes and accumulate at high concentrations intracellularly.Citation73 In our study, at 30 and 60 minutes of incubation, we found that colony number decreases.
Also the bioavailability of CIP is elevated in humans in the presence of CQ.Citation74 Recently, it has become apparent that at least some of the immunomodulatory effects of AZM are due to its action on macrophages.Citation75 Previous studies have linked the immunomodulatory action of macrolides with changes in a variety of different molecular targets and signaling processes, including activation of the lysosomes in which AZM accumulates.Citation76 We hypothesized that the effect of CQ-induced alkalinization on intra-lysosomal pH might be reduced at 90 minutes postinfection, subsequently leading to less uptake of AZM (as acid pH conditions markedly decrease the activity of macrolides). Due to the reduced uptake of AZM by the macrophages, the killing of intracellular bacteria would be reduced and ultimately result in the survival of intracellular S. aureus at 90 minutes postinfection. AZM is an antibacterial agent that, together with other members of the macrolide group of antibiotics, has been shown under a variety of clinical and experimental conditions to exert anti-inflammatory and immunomodulatory actions.Citation77 Other studies on the intracellular accumulation and activity of CIP using different cellular models of S. aureus-infected phagocytes have also reported a bacteriostatic rather than bactericidal effect of fluoroquinolone.Citation78 These concordant data clearly indicate that as yet unknown factors must have worked together to decrease the intracellular antibiotic efficiency of fluoroquinolone.
CIP, a quinolone antibiotic, is lysosomotropic and has been found to induce apoptosis (cell death) acting independently from ROS.Citation79 On the other hand, an in vitro combination of CIP and CQ was reported to have a significant additive effect on a clinical isolate of Plasmodium falciparum.Citation80 Macrophage phagocytosis is associated with a burst of respiratory activity that results in the production of a variety of molecules and free radicals called ROS, such as superoxide anion and H2O2. These ROS can damage lipids, proteins, and nucleic acids. LPO level has been used as a marker of oxidative stress. SOD, catalase, and GSH are important scavengers of superoxide anion and H2O2. These enzymes protect the generations of hydroxyl radicals and protect the cellular constituents from oxidative damage. As ROS and nitrogen intermediates are the key players in the clearance of intracellular pathogens (in macrophages), we hypothesized that the antioxidant and anti-inflammatory properties of CIP or AZM might be responsible for the killing of the intracellular pathogens tested. To validate our hypothesis, CQ-treated/-untreated macrophages were infected with S. aureus to induce the generation of ROS and NO in the cells, and the amounts of H2O2, superoxide anion, and NO released in the supernatant as well as in the cell-free lysate were analyzed in these cells with and without CIP or AZM treatment. The increased killing of bacterial cells in the CQ-treated macrophages that was found may be due to greater production of H2O2 and superoxide anion, and this result may be because of the bactericidal action of CQ.Citation81 More killing of bacterial cells has been observed with AZM and CIP treatment in CQ-pretreated infected macrophages but less production of H2O2 was observed in our study. This may be because macrolides like AZM inhibit ROS production (which is time dependent).Citation82
GSH, being the major intracellular thiol, provides protection against oxidative injury. GSH depletion may be associated with augmentation of an oxidative stress-mediated production of proinflammatory cytokines, particularly TNF-α and IFN-γ, because GSH is reported to play an important role in the polarization of Th1/Th2 balance.Citation83 The increase of GSH and decrease of LPO in CQ-treated groups may be indicators of a reduction in oxidative stress, which is more prominent when CQ is combined with AZM/CIP.Citation84
In this study, the oxidative stress generated by the macrophages was well orchestrated by the antioxidant defense mechanism of the cell, as was quite noticeable from the enhanced catalase and SOD activity in the supernatant as well as in the lysate when the CQ-pretreated macrophages were incubated with CIP or AZM before S. aureus infection. The observed increases in catalase activity may be as a result of cell-mediated immunity involving macrophages that may have provided several highly reactive molecules and ions such as H2O2, hydroxyl ions, and oxygen ions. The reason for this observed result may be due to the fact that the same site produces superoxide anion for dismutation to H2O2, the substrate for catalase, and glutathione peroxidase. The latter enzyme was not considered in this study and the possibility exists that this enzyme may have been involved in the neutralization of and conversion of H2O2. In addition, this enzyme has been reported to react with superoxide and hydroxyl radicals. It has also been reported that catalase activity is extended when cells are treated with pharmacological agents that delay proteolysis by decreasing lysosome acidification.Citation85 As internalized catalase (as was evident in the lysate after 90 minutes) has access to intracellular oxidants diffusing from all directions, intracellular catalase has the potential to be more effective than catalase released in the medium after 90 minutes. Therefore, the intracellular delivery of catalase may permit effective interception and detoxification of ROS produced inside the cells and diffusing through the plasma membrane from released activated macrophages.Citation86 It has been reported that catalase delivered by nanoparticles affected antioxidant protection for over 2 hours following internalization. It has also been found that this therapeutic time window can be further extended by the use of drugs, which either inhibit pH-dependent lysosomal proteolysis or prevent microtubule-dependent lysosomal trafficking.Citation85 Enhanced catalase activity in the AZM-pretreated group in our study may be because of the anti-inflammatory property of macrolides. The increase in catalase activity might be a defense mechanism for the bacteria. Enzyme activities are affected by many factors, including temperature, pH, and enzyme inhibitors/activators. Enzyme-catalyzed reaction rate, which is usually used to measure enzyme activity, is also affected by the concentration of the substrate and enzyme. The anti-inflammatory activity of macrolides may reduce oxidative response, and sequentially reduce production of ROS, including the substrate of catalase, H2O2.
This raises the possibility that pharmacological agents that are already approved for clinical use in rheumatoid arthritis (here, CQ) in combination with antibiotics (AZM or CIP) to improve the in vivo efficacy of AZM uptake is an approach with the potential to enhance therapeutic intervention in acute pathological conditions associated with infection-induced oxidative stress, inflammation, and septic shock.Citation87
The presence of intracellular S. aureus at 90 minutes postinfection in the macrophages pretreated with CQ and those incubated with CIP/AZM before being infected with S. aureus (due to less AZM uptake and less alkalinization induced by CQ) may automatically trigger the synthesis of SOD. This may point to the fact that the induced oxidative stress seen in the decline in LPO level in the combined-treatment group may not have necessitated the use of SOD to its optimum level. The substrate for this enzyme may have emerged as a result of xenobiotic metabolism. The reason for the observed increase in SOD activity in this study may be the result of the combined effect of intracellular S. aureus and CQ metabolism through a biotransformation pathway that may produce superoxide anion.Citation88
Our study reveals that CQ treatment in the presence of AZM/CIP decreases the level of proinflammatory cytokines, particularly TNF-α and IFN-γ, which agrees with the report that CQ is effective in the treatment of diseases associated with the increased release of proinflammatory cytokines.Citation89,Citation90 Some recent studies have shown that CQ administration attenuates the production of TNF-α; others have confirmed that CQ exerts some anti-inflammatory effects through downregulation of TNF-α production and signaling in macrophages as well as cytokine pattern production. TNF-α was able to enhance the oxidative response of optimally stimulated PMN and interfere with macrolides. An increased uptake of AZM has been reported in IFN-γ- and TNF-α -treated human macrophages and IFN-γ-treated THP-1 cells.Citation62,Citation89,Citation90 In the murine macrophage cell line RAW 264.7, 100 μM CQ interfered with the post-translational processing of TNF-α.Citation62 Alkalinizing agents including CQ inhibit the expression and release of TNF-α from Candida-stimulated human peripheral blood mononuclear cells.Citation62 Our findings are supported by a previous study evaluating the effect of CQ (100 μM) on TNF-α release from human whole blood showing that CQ interferes with TNF-α release in human cells at physiologic concentrations. A previous study using murine monocytic cell lines which demonstrated that CQ (100 μM) inhibited lipopolysaccharides-stimulated TNF-α release also supports our study.Citation91 Anti-mycobacterial synergy between TNF-α and AZM has been reported in a model of human macrophages. The same group observed increased uptake of AZM by macrophages in the presence of TNFα or IFN-γ.Citation92 Similarly, it has been shown that IFN-γ enhances the cellular accumulation of AZM in the human myelomonocytic cell line THP-1. In earlier studies on RAW 264.7 macrophage cell lines, it was reported that CQ, AZM, and several other macrolides inhibited proinflammatory cytokine production.Citation76
Previous research marked the transition of antimalarial agents from the treatment of malaria to that of rheumatic diseases.Citation96 However, the clinical implication of our data in relation to S. aureus infection-induced arthritis or sepsis might be speculative. TNF-α and IFN-γ are critical for the development of both innate and adaptive immune response. On the other hand, it has been argued that the pathogenesis of septic shock and arthritis involves the deregulated production of TNF-α and IFN-γ and other proinflammatory mediators.Citation93 Therefore, numerous studiesCitation94 have postulated that the immunomodulation of proinflammatory cytokines like TNF-α and IFN-γ would reduce sepsis-related morbidity and mortality. By further dissecting the mechanism by which CQ in the presence of AZM reduces TNF-α and IFN-γ release and ROS production, it might be possible to design congeners with either greater or lesser anti-inflammatory properties. Such drugs in combination could prove more efficacious in certain disease states.
Previous work has indicated that, for optimal NO production, macrophages need to be primed with IFN-γ. Moreover, TNF-α was able to enhance the oxidative response of macrophages and prime the effect of AZM uptake.Citation95 Since acid pH inhibits CQ accumulation in macrophages,Citation97 we hypothesized that at 90 minutes there might be less CQ and that the alkalinizing effect of CQ on phagosomes would withdraw; as a result of this, we postulated that there would be less uptake of AZM and this would subsequently lead to reduced TNF-α and IFN-γ production. Therefore, due to the priming effect induced by TNF-α and IFN-γ on AZM uptake, NO production is also limited and the number of bacteria that survive intracellularly within the macrophages is increased by suppressing the induction of inducible NO synthase messenger RNA in a dose-dependent manner.Citation98 NO production is further reduced when AZM is administered after CQ pretreatment of infected macrophages, thus inhibiting the production of harmful hydroxyl radicals and peroxynitrite ions further confirming immunomodulatory action of CQ by protecting host cells.Citation99
CQ is commonly used to study the role of endosomal acidification in cellular processes such as intracellular TLR signaling.Citation100 CQ’s antagonistic effect on TLRs and cytokines is reported to be potentially relevant and beneficial to arthritis.Citation101 In order to function, intracellular TLRs require an acidic pH; probably they are proteolyzed by acid-dependent proteases that function normally within the acidic endoplasm of lysosomes. CQ, by altering the lysosomal pH, possibly prevents the functional transformation of intracellular TLRs and inhibits their function. Research published recently proposes that TLR antagonism by CQ might not be driven entirely by endosomal pH alteration but by mechanical inhibition, whereby antimicrobial agents structurally bind to nucleic acids and mask their TLR-binding epitopes.Citation102,Citation103
Conclusion
This study aimed to determine if there was any alteration in the killing of S. aureus in murine peritoneal macrophages when CQ was used alone compared with in combination with CIP or AZM, and also to find out the implications of ROS production and cytokine release in the intracellular killing of S. aureus by the macrophages. It was found that S. aureus-triggered cytokine response was regulated when the macrophages were co-cultured with CQ and AZM as compared with CQ stimulation only. It can be suggested that the action of AZM in mediating bacterial killing is enhanced in the presence of CQ, indicating that there is an enhanced uptake of AZM during early infection that may be essential for the killing of bacteria by AZM. Reduction of the oxidative stress burden on the S. aureus-infected macrophages may pave the way for better killing of internalized S. aureus by CQ plus CIP or CQ plus AZM. Based on these observations, it can be concluded that CQ loaded with AZM elicits a stronger proinflammatory response by increasing the intracellular uptake of AZM thus enabling the immune system to mount a more robust and prolonged response against intracellular pathogens.
Acknowledgments
This work was supported by a grant from the Department of Biotechnology, Government of West Bengal, Calcutta, West Bengal, India (grant number: vide sanction order 786-BT/Estt/RD-4/13; date: October 30, 2013) to Biswadev Bishayi. Biswadev Bishayi is indebted to the Department of Science and Technology, Government of India, for providing the instruments procured under the Department of Science and Technology Promotion of University Research and Scientific Excellence program to the Department of Physiology, University of Calcutta.
Disclosure
The authors declare no conflicts of interest in this work. The authors also state that they have no direct financial relation with the commercial identities mentioned in this paper that might lead to a conflict of interest.
References
- LowyFDStaphylococcus aureus infectionsN Engl J Med199833985205329709046
- BrouilletteEGrondinGShkretaLLacassePTalbotBGIn vivo and in vitro demonstration that Staphylococcus aureus is an intracellular pathogen in the presence or absence of fibronectin-binding proteinsMicrob Pathog200335415916812946328
- GordonRJLowyFDPathogenesis of methicillin-resistant Staphylococcus aureus infectionClin Infect Dis200846Suppl 5S350S35918462090
- CarrynSChanteuxHSeralCMingeot-LeclercqMPVan BambekeFTulkensPMIntracellular pharmacodynamics of antibioticsInfect Dis Clin North Am200317361563414711080
- van den BroekPJAntimicrobial drugs, microorganisms, and phagocytesRev Infect Dis19891122132452649959
- KrutOSommerHKrönkeMAntibiotic-induced persistence of cytotoxic Staphylococcus aureus in non-phagocytic cellsJ Antimicrob Chemother200453216717314729736
- Al-NawasBShahPMIntracellular activity of ciprofloxacin and moxifloxacin, a new 8-methoxyquinolone, against methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother19984166556589687105
- FosterTJImmune evasion by staphylococciNat Rev Microbiol200531294895816322743
- ZetolaNFrancisJSNuermbergerELBishaiWRCommunity-acquired meticillin-resistant Staphylococcus aureus: an emerging threatLancet Infect Dis20055527528615854883
- HiramatsuKVancomycin-resistant Staphylococcus aureus: a new model of antibiotic resistanceLancet Infect Dis20011314715511871491
- GreshamHDLowranceJHCaverTEWilsonBSCheungALLindbergFPSurvival of Staphylococcus aureus inside neutrophils contributes to infectionJ Immunol200016473713372210725730
- HudsonMCRampWKNicholsonNCWilliamsASNousiainenMTInternalization of Staphylococcus aureus by cultured osteoblastsMicrob Pathog19951964094198852281
- BaylesKWWessonCALiouLEFoxLKBohachGATrumbleWRIntracellular Staphylococcus aureus escapes the endosome and induces apoptosis in epithelial cellsInfect Immun19986613363429423876
- MenziesBEKourtevaIInternalization of Staphylococcus aureus by endothelial cells induces apoptosisInfect Immun19986612599459989826383
- DasDSahaSSBishayiBIntracellular survival of Staphylococcus aureus: correlating production of catalase and superoxide dismutase with levels of inflammatory cytokinesInflamm Res200857734034918607538
- DasDBishayiBStaphylococcal catalase protects intracellularly survived bacteria by destroying H2O2 produced by the murine peritoneal macrophagesMicrob Pathog2009472576719439176
- DasDBishayiBContribution of Catalase and Superoxide Dismutase to the Intracellular Survival of Clinical Isolates of Staphylococcus aureus in Murine MacrophagesIndian J Microbiol201050437538422282603
- LowyFDIs Staphylococcus aureus an intracellular pathogen?Trends Microbiol20008834134310920387
- HamzaTLiBDifferential responses of osteoblasts and macrophages upon Staphylococcus aureus infectionBMC Microbiol20141420725059520
- MalPDuttaKBandyopadhyayDBasuAKhanRBishayiBAzithromycin in combination with riboflavin decreases the severity of Staphylococcus aureus infection induced septic arthritis by modulating the production of free radicals and endogenous cytokinesInflamm Res201362325927423229721
- MalPDuttaSBandyopadhyayDDuttaKBasuABishayiBGentamicin in combination with ascorbic acid regulates the severity of Staphylococcus aureus infection-induced septic arthritis in miceScand J Immunol201276652854022924656
- MalPGhoshDBandyopadhyayDDuttaKBishayiBAmpicillin alone and in combination with riboflavin modulates Staphylococcus aureus infection induced septic arthritis in miceIndian J Exp Biol2012501067768923214260
- RogersDETompsettRThe survival of staphylococci within human leukocytesJ Exp Med195295220923014907971
- UnderhillDMGoodridgeHInformation processing during phagocytosisNat Rev Immunol201212749250222699831
- PechèreJCThe activity of azithromycin in animal models of infectionEur J Clin Microbiol Infect Dis199110108218271662625
- van den BroekPJActivity of antibiotics against microorganisms ingested by mononuclear phagocytesEur J Clin Microbiol Infect Dis19911021141181864273
- TulkensPMIntracellular distribution and activity of antibioticsEur J Clin Microbiol Infect Dis19911021001061864271
- SeralCBarcia-MacayMMingeot-LeclercqMPTulkensPMVan BambekeFComparative activity of quinolones (ciprofloxacin, levofloxacin, moxifloxacin and garenoxacin) against extracellular and intracellular infection by Listeria monocytogenes and Staphylococcus aureus in J774 macrophagesJ Antimicrob Chemother200555451151715731197
- NguyenHAGrelletJPaillardDDuboisVQuentinCSauxMCFactors influencing the intracellular activity of fluoroquinolones: a study using levofloxacin in a Staphylococcus aureus THP-1 monocyte modelJ Antimicrob Chemother200657588389016533826
- HallIHSchwabUEWardESIvesTDisposition and intracellular levels of moxifloxacin in human THP-1 monocytes in unstimulated and stimulated conditionsInt J Antimicrob Agents200322657958714659655
- LabroMTImmunomodulatory action of antibacterial agentsClinical Immunotherapy199666454464
- LabroMTIntraphagocytic penetration of macrolide antibioticsBryskierAJButzlerJPNeuHCTulkensPMMacrolides: Chemistry, Pharmacology and Clinical UseParisArnette Blackwell1993379388
- VergneIConstantPLanéelleGPhagosomal pH determination by dual fluorescence flow cytometryAnal Biochem199825511271329448851
- PooleBOhkumaSEffect of weak bases on the intralysosomal pH in mouse peritoneal macrophagesJ Cell Biol19819036656696169733
- ChakravorttyDHenselMInducible nitric oxide synthase and control of intracellular bacterial pathogensMicrobes Infect2005562162712787738
- PollockJDWilliamsDAGiffordMAMouse model of X-linked chronic granulomatous disease, an inherited defect in phagocyte super-oxide productionNat Genet199592022097719350
- van SorgeNMBeasleyFCGusarovIMethicillin-resistant Staphylococcus aureus bacterial nitric-oxide synthase affects antibiotic sensitivity and skin abscess developmentJ Biol Chem31201328896417642623322784
- CarlierMBGarcia-LuqueIMontenezJPTulkensPMPiretJAccumulation, release and subcellular localization of azithromycin in phagocytic and non-phagocytic cells in cultureInt J Tissue React1994165–62112207558665
- Miossec-BartoliCPilatreLPeyronPThe new ketolide HMR3647 accumulates in the azurophil granules of human polymorphonuclear cellsAnitmicrob Agents Chemother1999431024572462
- CarlierMBScorneauxBZeneberghADesnottesJFTulkensPMAnimal model pharmacokinetics and pharmacodynamics: a critical reviewJ Anitmicrob Agents Chemother199026Suppl B2739
- AndesDCraigWAAnimal model pharmacokinetics and pharmacodynamics: a critical reviewInt J Antimicrob Agents200219426126811978497
- MaurinMRaoultDPhagolysosomal alkalinization and intracellular killing of Staphylococcus aureus by amikacinJ Infect Dis199416923303368106766
- SeralCVan BambekeFTulkensPMQuantitative analysis of gentamicin, azithromycin, telithromycin, ciprofloxacin, moxifloxacin, and oritavancin (LY333328) activities against intracellular Staphylococcus aureus in mouse J774 macrophagesAntimicrob Agents Chemother20034772283229212821480
- LevitzSMNongSHSeetooKFCryptococcus neoformans resides in an acidic phagolysosome of human macrophagesInfect Immun19996728858909916104
- AugustijnsPGeusensPVerbekeNChloroquine levels in blood during chronic treatment of patients with rheumatoid arthritisEur J Clin Pharmacol19924244294331307690
- NewmanSLGooteeLBrunnerGDeepeGSJrChloroquine induces human macrophage killing of Histoplasma capsulatum by limiting the availability of intracellular iron and is therapeutic in a murine model of histoplasmosisJ Clin Invest1994934142214298163646
- PicotSPeyronFVuillezJPPolackBAmbroise-ThomasPChloroquine inhibits tumor necrosis factor production by human macrophages in vitroJ Infect Dis199116448301894954
- YasudaHLeelahavanichkulATsunodaSChloroquine and inhibition of Toll-like receptor 9 protect from sepsis-induced acute kidney injuryAm J Physiol Renal Physiol20082945F1050F105818305095
- ThoméRMoraesASBombeiroALChloroquine treatment enhances regulatory T cells and reduces the severity of experimental autoimmune encephalomyelitisPlos One201386e6591323799062
- CooperRGMagwereTChloroquine: novel uses and manifestationsIndian J Med Res2008127430531618577785
- LeighPCVan FurthRVan ZwetTLIn vitro determination of phagocytosis and intracellular killing by polymorphonuclear and mononuclear phagocytesWeirDMHandbook of Experimental Immunology4th edOxfordBlackwell Scientific1986146
- AbsolomDRBasic methods for the study of phagocytosisMethods Enzymol1986132951803821529
- NandiDMishraMKBasuABishayiBProtective effects of interleukin-6 in lipopolysaccharide (LPS)-induced experimental endotoxemia are linked to alteration in hepatic anti-oxidant enzymes and endogenous cytokinesImmunobiology2010215644345119773091
- AvciEAkarslanZZErtenHCoskun-CevherSOxidative stress and cellular immunity in patients with recurrent aphthous ulcersBraz J Med Biol Res201447535536024760117
- SedlakJLindsayRHEstimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagentAnal Biochem19682511922054973948
- ColowickSPKaplanNOAbsolomDRBasic methods for the study of phagocytosisSabatoGDEverseJMethods in Enzymology132Immunochemical techniques part J: phago-cytosis and cell-mediated cytotoxicityNew YorkAcademic Press1986160
- BradfordMMA rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye bindingAnal Biochem197672248254942051
- BishayiBBandyopadhyayDMajhiAAdhikaryRPossible role of Toll-like receptor-2 in the intracellular survival of Staphylococcus aureus in murine peritoneal macrophages: involvement of cytok-ines and anti-oxidant enzymesScand J Immunol201480212714324846691
- BishayiBBandyopadhyayDMajhiAAdhikaryRExpression of CXCR1 (Interleukin-8 Receptor) in Murine Macrophages After Staphylococcus aureus Infection and its Possible Implication on Intracellular Survival Correlating with Cytokines and Bacterial Anti-Oxidant EnzymesInflammation Epub8172014 Available from: http://www.ncbi.nlm.nih.gov/pubmed/25129059Accessed August 17, 2014
- IgnatowskiTABidlackJMDifferential kappa-opioid receptor expression on mouse lymphocytes at varying stages of maturation and on mouse macrophages after selective elicitationJ Pharmacol Exp Ther1999290286387010411603
- PolliackAGordonSScanning electron microscopy of murine macrophages. Surface characteristics during maturation, activation, and phagocytosisLab Invest19753354694771186128
- WeberSMLevitzSMChloroquine interferes with lipopolysaccharide-induced TNF-alpha gene expression by a nonlysosomotropic mechanismJ Immunol200016531534154010903761
- OhkumaSPooleBFluorescence probe measurement of the intralysosomal pH in living cells and the perturbation of pH by various agentsProc Natl Acad Sci U S A19787573327333128524
- LeeHand WHandDebra LCharacteristics and mechanisms of azithromycin accumulation and efflux in human polymorphonu-clear leukocytesInternational Journal of Antimicrobial Agents20011841942511711255
- HandWLKing-ThompsonNLHolmanJWEntry of roxithromycin (RU 965), imipenem, cefotaxime, trimethoprim, and metronidazole into human polymorphonuclear leukocytesAnitmicrob Agents Chemother1987311015531557
- LevitzSMHarrisonTSTabuniALiuXChloroquine induces human mononuclear phagocytes to inhibit and kill Cryptococcus neoformans by a mechanism independent of iron deprivationJ Clin Invest19971006164016469294133
- LevitzSM1NongSHSeetooKFHarrisonTSSpeizerRASimonsERCryptococcus neoformans resides in an acidic phago-lysosome of human macrophagesInfect Immun199967228858909916104
- FoxRIMechanism of action of hydroxychloroquine as an antirheumatic drugSemin Arthritis Rheum1993232 Suppl 182918278823
- GeisowMJD’Arcy HartPYoungMRTemporal changes of lysosome and phagosome pH during phagolysosome formation in macrophages: studies by fluorescence spectroscopyJ Cell Biol19818936456526166620
- SteinbergTHHandWLEffect of phagocyte membrane stimulation on antibiotic uptake and intracellular bactericidal activityAntimicrob Agents Chemother19873146606623606071
- LeiperKMorrisAIRhodesJMOpen label trial of oral clarithromycin in active Crohn’s diseaseAliment Pharmacol Ther200014680180610848665
- FlanaganPKCampbellBJRhodesJMOC-140 Hydroxychloroquine as a treatment for crohn’s disease: enhancing antibiotic efficacy and macrophage killing of E coliGut201261A60A61
- OliphantCMGreenGMQuinolones: a comprehensive reviewAm Fam Physician200265345546411858629
- IloCEIlonduNAOkwoliNEffect of chloroquine on the bioavailability of ciprofloxacin in humansAm J Ther200613543243516988539
- VrančićMBanjanacMNujićKAzithromycin distinctive-lymodulates classical activation of human monocytes in vitroBr J Pharmacol201216551348136021726210
- MunićVBanjanacMKoštrunSIntensity of macrolide anti-inflammatory activity in J774A.1 cells positively correlates with cellular accumulation and phospholipidosisPharmacol Res201164329830721473915
- CrosbiePAWoodheadMALong-term macrolide therapy in chronic inflammatory airway diseasesEur Respir J200933117118119118228
- AndersonRJoonéGKIn vitro investigation of the intraphagocytic bio-activities of ciprofloxacin and the new fluoroquinolone agents, clinafloxacin (CI-960) and PD 131628Chemotherapy19933964244318222871
- BoyaPAndreauKPoncetDLysosomal membrane permeabilization induces cell death in a mitochondrion-dependent fashionJ Exp Med2003197101323133412756268
- KazzimOJAdegbolagunOMOshoOAnumuduCIAdditive effects of ciprofloxacin on the in-vitro activity of chloroquine against a clinical isolate of Plasmodium falciparumAnn Trop Med Parasitol2006100757958416989683
- AbrantesPDimopoulosGGrossoARdo RosárioVESilveiraHChloroquine mediated modulation of Anopheles gambiae gene expressionPLoS One200837e258718596975
- KanohSRubinBKMechanisms of action and clinical application of macrolides as immunomodulatory medicationsClin Microbiol Rev201023359061520610825
- MurataYShimamuraTHamuroJThe polarization of T(h)1/T(h)2 balance is dependent on the intracellular thiol redox status of macrophages due to the distinctive cytokine productionInt Immunol200214220121211809739
- EliaACPrearoMPaciniNDörrAJAbeteMCEffects of selenium diets on growth, accumulation and antioxidant response in juvenile carpEcotoxicol Environ Saf201174216617320554323
- CheesemanKHSlaterTFAn introduction to free radical biochemistryBr Med Bull1993494814938221017
- WolinMSGupteSAOecklerRASuperoxide in the vascular systemJ Vasc Res200239319120712097818
- MuroSCuiXGajewskiCMurcianoJCMuzykantovVRKovalMSlow intracellular trafficking of catalase nanoparticles targeted to ICAM-1 protects endothelial cells from oxidative stressAm J Physiol Cell Physiol20032855C1339C134712878488
- IyaweHOOnigbindeAOAinaOOEffect of chloroquine and ascorbic acid interaction on the oxidative stress status of Plasmodium berghei infested miceInt J Pharmacol20062114
- SacreKCriswellLAMcCuneJMHydroxychloroquine is associated with impaired interferon-alpha and tumor necrosis factor-alpha production by plasmacytoid dendritic cells in systemic lupus erythematosusArthritis Res Ther2012143R15522734582
- OuyangQHuangZWangZChenXNiJLinLEffects of pristane alone or combined with chloroquine on macrophage activation, oxidative stress, and Th1/Th2 skewnessJ Immunol Res201420142014 Article ID 613136
- JeongJYJueDMChloroquine inhibits processing of tumor necrosis factor in lipopolysaccharide-stimulated RAW 264.7 macrophagesJ Immunol199715810490149079144507
- VazifehDBryskierALabroMTEffect of proinflammatory cytokines on theinterplay between roxithromycin, HMR 3647, or HMR 3004 and human polymorphonuclear neutrophilsAntimicrob Agents Chemother200044351152110681311
- KolliasGDouniEKassiotisGKontoyiannisDThe function of tumour necrosis factor and receptors in models of multi-organ inflammation, rheumatoid arthritis, multiple sclerosis and inflammatory bowel diseaseAnn Rheum Dis1999581132139
- SchulteWBernhagenJBucalaRCytokines in sepsis: potent immunoregulators and potential therapeutic targets–an updated viewMediators Inflamm2013201316597423853427
- MosserDMZhangXActivation of murine macrophagesCurr Protoc Immunol20081414.2
- FurstDEPharmacokinetics of hydroxychloroquine and chloroquine during treatment of rheumatic diseasesLupus19965Suppl 1S11S158803904
- NujićKBanjanacMMunićVPolančecDEraković HaberVImpairment of lysosomal functions by azithromycin and chloroquine contributes to anti-inflammatory phenotypeCellular Immunol20122791788623099154
- ParkYCPaeHOYooJCChoiBMJueDMChungHTChloroquine inhibits inducible nitric oxide synthase expression in murine peritoneal macrophagesPharmacol Toxicol199985418819110563518
- TamaokiJKondoMKohriKAoshibaKTagayaENagaiAMacrolide antibiotics protect against immune complex-induced lung injury in rats: role of nitric oxide from alveolar macrophagesJ Immunol199916352909291510453038
- RutzMMetzgerJGellertTToll-like receptor 9 binds single-stranded CpG-DNA in a sequence- and pH-dependent mannerEur J Immunol20043492541255015307186
- BlasiusALBeutlerBIntracellular toll-like receptorsImmunity201032330531520346772
- KuznikABencinaMSvajgerUJerasMRozmanBJeralaRMechanism of endosomal TLR inhibition by antimalarial drugs and imidazoquinolinesJ Immunol201118684794480421398612
- WallaceDJGudsoorkarVSWeismanMHVenuturupalliSRNew insights into mechanisms of therapeutic effects of antimalarial agents in SLENat Rev Rheumatol20128952253322801982