Abstract
Traditionally, the signaled avoidance (SA) paradigm has been used in an attempt to better understand human phobia. Animal models of this type have been criticized for ineffectively representing phobia. The SA model characterizes phobia as an avoidance behavior by presenting environmental cues, which act as warning signals to an aversive stimulus (ie, shock). Discriminated conditioned punishment (DCP) is an alternative paradigm that characterizes phobia as a choice behavior in which fear serves to punish an otherwise adaptive behavior. The present study quantifies the differences between the paradigms and suggests that DCP offers an alternative paradigm for phobia. Rats trained on either SA or DCP were compared on a number of behavioral variables relevant to human phobia. Results indicate that rats in the DCP paradigm responded significantly earlier to warning signals and were more effective at preventing shocks than rats in the SA paradigm. Implications of this alternative paradigm are discussed.
Introduction
In a recent paper, Kessler et al reported that phobias are among the most commonly reported psychological disorders.Citation1 The prominent role of fear as a feature of phobias has long garnered the attention of theorists focusing on the way in which fear responses are conditioned.Citation2 Watson and Rayner first proposed that emotions in general, and fear specifically, can be the result of classical conditioning and experience.Citation3 The role of both classical and operant conditioning continues to be a primary area of focus among many theorists hoping to draw parallels between theories of conditioning and pathologies, such as human phobia.Citation2,Citation4
Much of the work in this area has focused on the relationship between avoidance behaviors and phobias. Mowrer most famously proposed that fear develops as a function of a two-process theory of learning.Citation5 In this two-process theory, Mowrer proposed that fear develops as a function of both classical and operant conditioning. A neutral stimulus paired in time and space with a naturally fear-eliciting unconditioned stimulus (US) develops the ability to elicit fear, becoming a conditioned stimulus (CS), as a function of the principles of classical conditioning. The newly acquired CS then serves as a warning signal, predicting the future arrival of the US. Upon the presentation of the warning signal, an organism can learn a response that allows for the escape from the warning signal and prevents exposure to the US. This learned avoidance response is then reinforced via the principles of operant conditioning. As reported by Mineka, this view of fear conditioning has been a dominant force in the development of theory and models of human fear pathology, including phobia.Citation2
Critics have indicated dissatisfaction in the ability of Mowrer’s theory and of learning models to truly illustrate human phobia.Citation4,Citation5 Costello identified a number of critical features of the human phobic situation that are not reflected in the two-factor avoidance analogs of phobias.Citation6 One such feature was the inability of the warning signals predicting the phobic stimuli to elicit fear. Costello noted that in human phobia, environmental cues that predict the phobic stimuli elicit fear – a feature that signaled avoidance (SA) models have struggled to demonstrate.Citation6 Indeed, as Costello pointed out, work in animal conditioning of fear via SA has provided little evidence of the fear-eliciting properties of classically conditioned warning signals.Citation6 Sidman and Boren successfully trained rats to prevent shocks as an operant response.Citation7 When a 5-second tone was added just prior to the shock, rats delayed their shock-preventing response until the tone (still avoiding the shock) but never learned to respond to avoid both the tone and the shock. If the tone itself should become fear-eliciting via classical conditioning, as proposed by Mowrer, animals would be expected to respond sooner to avoid, not only the shock but also, the fear elicited by the tone itself.Citation5 Mineka, although more sympathetic to a conditioning analysis of phobias, also argued that avoidance learning fails to model human phobia.Citation8 Like Costello, she concluded that although rats can avoid shock, there is no evidence that animals could be trained to avoid warning signals that predict the shock.Citation6 Mineka noted that in naturally occurring phobias (ie, not created experimentally), successful avoidance of environmental cues predicting the phobic stimuli is very common.Citation8 Stampf, in response to Mineka’s critique, described several serial avoidance studies involving sequential cues leading to shock.Citation9–Citation11 If the warning signals were fear-evoking, as predicted by Mowrer, animals would be expected to respond earlier in the sequence of warning signals, avoiding, not only the shock (US) but also, the preceding cues in the sequence (CS).Citation5 These experiments provided mixed results, with animals usually responding at the last opportunity, suggesting an absence of fear of the CS. Stampfl noted that animals did frequently prevent CSs that immediately preceded the shock from occurring, thereby providing some evidence that fear of the CS had developed.Citation9 Despite this additional data from serial stimuli, Stampfl acknowledged that the results were not suffcient to model these factors for phobias: “… [H]uman phobics typically respond predominantly to stimuli early in the sequence, but rats did not display this strong ‘fixation to early stimuli.’”Citation9 In addition, a second study of serial SA using a different methodology provided similar evidence of typically delayed responding.Citation12
Costello further criticized animal models, suggesting that avoidance models fail to account for the maladaptive nature of phobic behavior.Citation6 Costello suggested that one important feature of a phobia is that it interferes with another adaptive, healthy behavior.Citation6 Indeed, one can imagine that the fear of an object only reaches the level of pathology if that fear is preventing an otherwise valued, adaptive behavior. In the typical avoidance paradigm, an animal learns to prevent the onset of shock by making a specific response. There is absolutely nothing maladaptive about such a behavior. Rather, the prevention of a painful shock is in fact a valuable and adaptive response. Costello argued that for a conditioning model of phobia to truly capture the human pathology, the model must reflect the detrimental nature of phobias.Citation6
The model described in this paper was created partially in response to the criticisms leveled against avoidance models. The discriminated conditioned punishment (DCP) model argues that phobias do most of their harm as “punishers” because they interfere with otherwise rewarding behaviors. Similar to the argument first proposed by Dinsmoor, we argue that many of the weaknesses of conditioning models of fear and phobia stem from a mistaken focus on the avoidance response rather than punishment.Citation13 Indeed, careful inspection of Stampfl demonstrates this potential problem.Citation9 When developing an analogy to link animal models with phobia, Stampfl applied the terminology of punishment in the example of a person with a bridge phobia selecting one of two paths to a destination, one of which required the use of a bridge.Citation9 With two paths to the same goal (reinforcement) available, the person with the phobia chose to take the path that did not require crossing the bridge. In this fantasy scenario, the individual chose the response path that was rewarded without exposure to the feared stimulus: the bridge. The bridgeless path was simply reinforced whereas the alternative was both reinforced and punished by the bridge. Stampfl’s recognition of the potential role of punishers in the effect of phobias was unfortunately not pursued experimentally.Citation9 When devising an experiment to recreate this scenario in the lab, his experimental paradigm and his methodological language focused on avoidance, where animals were mechanically moved toward an aversive stimulus and were forced to respond to prevent exposure.Citation14
The DCP paradigm models the example put forth in Stampfl’s bridge example.Citation14 Specifically, phobic stimuli and the warning signals that predict them serve as punishers, suppressing behaviors that are otherwise adaptive and reinforced. Given an alternative response that provides reinforcement, individuals choose the response that is free of punishment. The increase in the frequency of the nonpunished response reflects the suppression of the punished response. The failure to perform the response that elicits the phobic stimuli can best be understood and studied as punishment rather than avoidance.
The current experiment compares an animal model of DCP with a traditional sequential SA model. The DCP model utilizes a concurrent schedule in which two alternatives lead to reinforcement. Additionally, one of those alternatives, assigned at random on each trial, is also punished via both shock and warning signals. We hypothesized that the DCP model better represents human phobia, and as such, would more successfully answer the aforementioned criticisms of avoidance models. Specifically, animals trained under DCP would demonstrate greater fear of the warning signals – evidenced by responding earlier in the sequence of warning signals – and also greater resistance to extinction than would be found with the SA model. Each of these features has long been held as an essential portion of an accurate animal model of phobia. We further argue that the DCP model better represents the irrational nature of phobias. In traditional SA models, the avoidance response to avoid shock is both reasonable and adaptive. Alternately, failure to respond in the DCP model prevents the presentation of the aversive stimulus but also interferes with an otherwise adaptive behavior, ie, a lever press that produces positive reinforcement via food presentation.
Material and methods
All procedures involving animals in this study were approved by the Providence College Institutional Animal Care and Use Committee and were in compliance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Subjects
Subjects were male outbred Sprague-Dawley® rats (N = 20) (Charles River Laboratories International, Inc, Wilmington, MA, USA), weighing 225–250 g at arrival. Upon arrival, the animals were housed in individual cages (25 × 25 × 35 cm). Having established a baseline weight with ad libitum feeding, animals were placed on a restricted diet, resulting in a 10% decrease in the ad libitum weight. Water was available ad libitum in the home cage for the duration of the experiment. The animals were kept under a day/night schedule (lights on from 8 am to 8 pm) in a temperature (20°C–24°C) and humidity (40%–60%) controlled environment. All animals were trained during the light portion of the light-dark schedule.
Apparatus
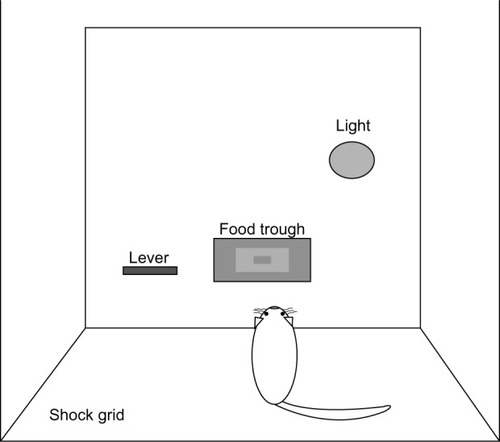
Ten sound-attenuating cubicles containing operant conditioning chambers (30.5 × 24.1 × 29.2 cm; Med Associates Inc, St Albans, VT, USA) were utilized for all operant experiments. As seen in , boxes were arranged as a function of the training paradigm. Chambers assigned to the DCP training were equipped with two levers, one on each side of the food delivery trough. Two warning lights, one on each side of the chamber, were placed above the response levers. The remaining chambers, illustrated in , were used for the SA training. SA boxes were identical in organization but contained just one lever arranged to the left of the food delivery trough and a single warning light placed on the opposite side of the food trough. The floors of all chambers consisted of stainless steel grids, allowing for the delivery of scrambled shock.
Figure 1 Discriminated conditioned punishment (DCP) operant chamber arrangement.
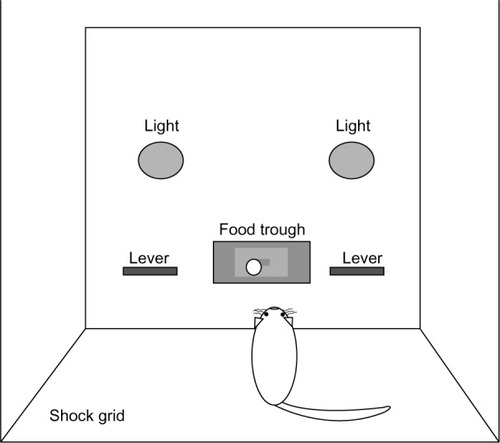
Figure 2 Signaled avoidance (SA) operant chamber arrangement.
Procedure
Discriminated conditioned punishment
Following magazine training, animals were shaped to respond for continuous reinforcement (45 mg pellet) on both the left and right operandum. Once responding was established, animals were then exposed to a free-choice, forced-choice schedule utilizing both levers. Initially, reinforcement was available on both levers (free-choice). Following reinforcement, the lever previously utilized was no longer reinforced, forcing animals to respond on the opposite operandum (forced choice). Having now responded on each operandum, this procedure repeated itself, again providing a free-choice. This training procedure was intended to test for and eliminate any inherent operandum preferences in the animals that might have interfered with later conditioning. Two animals demonstrated very strong lever preferences and as a result were dropped from further training. Once responding on both levers was asymptotic, ratios were slowly increased until animals were responding on a fixed-ratio 20 (FR-20) schedule on both levers. Reliable responding on the FR-20 schedule represented the final stage of pretraining and served as the starting point for exposure to the DCP paradigm.
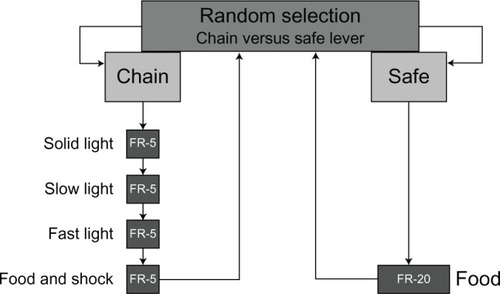
The DCP training program, as seen in , consisted of a concurrent schedule. The first contingency, a simple FR-20 (safe) schedule for food, was assigned to one of the two levers at random at the beginning of each trial. The 20th response on the safe operandum produced food reinforcement (45 mg). The second schedule, assigned to the remaining lever, was a chained procedure in which a series of warning signals was initiated (steady light, slow flashing light, fast flashing light) when the animals were responding on the chained schedule (FR-5, FR-5, FR-5, FR-5). Each subsequent warning signal was presented for every five lever presses, with the 20th response producing both food reinforcement (45 mg) and a 1-second shock (0.4 mA). The animals continued on this paradigm until 120 trials were completed or until 2 hours and 20 minutes of time had passed.
Figure 3 DCP contingency.
Signaled avoidance
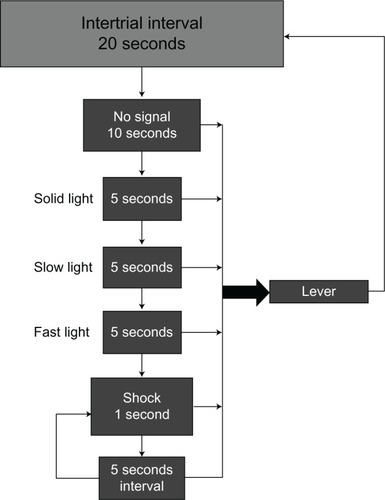
Training of the animals assigned to the SA paradigm began with the procedure illustrated in . The initial training on this procedure included the shaping of the avoidance response by the researchers via a remote hand operandum. Shaping of the response was terminated following the first sign of avoidance responding, and animals were continued on the training paradigm without additional shaping. After a 20-second intertrial interval (ITI), each trial began with a 10-second period during which no signals were presented. This was followed by a sequence of signals (steady light, slow flashing light, fast flashing light) each lasting 5 seconds, preceding the presentation of a 1-second shock (0.4 mA). The 1-second shock was repeated following an additional 5-second interval if the animal failed to respond. Responding at any point following the ITI, whether it was in the sequence of signals or following the presentation of shock, reset the 20-second ITI, and the process was repeated. Animals continued under this contingency for 1 hour per day or until 360 trials were completed. Two animals failed to demonstrate avoidance responding via this training method and were dropped from further participation in the experiment.
Figure 4 Signaled avoidance (SA) contingency.
Results
The outcome measures, “point of responding” and “percentage of shock received,” served as the dependent variables. Percentage of shock represented the proportion of trials in which the animals failed to prevent shock by making an appropriate response during the sequence of warning signals (switching levers from chain to safe in DCP, lever press in SA). Trials in which animals were not exposed to warning signals (ie, they chose the safe lever prior to exposure to response-initiated warning signals in DCP; responses during the 10-second prewarning signal interval in SA) were not included in further analyses. Point of responding was calculated as the period in the sequence of stimuli in which the animal responded (switched levers in the DCP paradigm or a lever press in the SA paradigm). A “1” indicated a response in the first stimulus, a “2” to the second, etc, with a “4” indicating the animal failed to respond and was shocked. A 2 × (3) mixed model analysis of variance (ANOVA) was implemented for each DV (dependent variable), with the training condition (DCP, SA) and phase of training (training, asymptote, extinction) serving as the between- and within-subject variables, respectively. The phase of training represented the progression of animals across time under each of the operant paradigms. Training included data from the initial 8 days of exposure to the DCP and SA paradigms. The asymptotic and extinction phases consisted of data from the final 16 days of training. Once the point of responding and percentage of shocks became stagnant (on the ninth day), the asymptotic phase began and continued for an additional 6 days of training. Extinction began on 16th day and continued for an additional 8 days. Under these paradigms, extinction consisted of the removal of the aversive stimulus (shock) from the contingencies. It is important to note that for analyses of shock percentage during the extinction phase, no shocks were actually received and instead, the value represented shocks that would have been received if the shock generator had not been removed. Other details of the contingencies, including the warning signals, were left untouched during the extinction phase. All analyses of the phase of training DV consisted of these 24 days of data. For all figures, error bars represent the standard error.
Point of responding
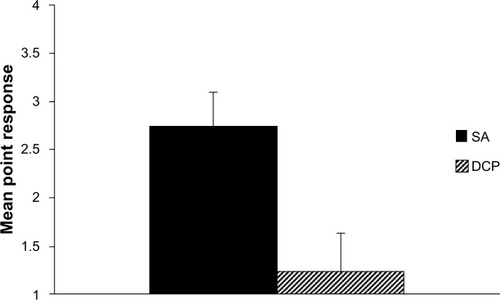
Tests of the main effect for the between-subject factor, training condition, were found to be statistically significant (F[1,14] = 2762.55, P < 0.001). As seen in , inspection of the means further indicated that animals trained under the DCP paradigm responded significantly earlier in the sequence of stimuli than those trained under the SA paradigm (mean [M] = 1.23, standard deviation [SD] = 0.20; M = 2.73, SD = 0.66, respectively).
Figure 5 Main effect, training condition (DV point of responding).
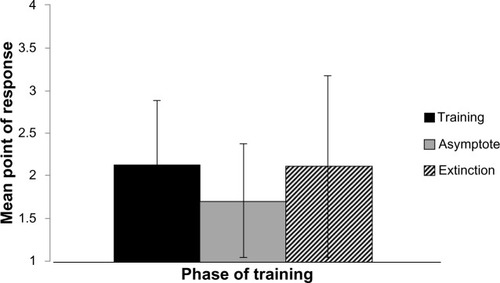
Tests of the main effect for the within-subject factor, phase of training, were found to be statistically significant (F[2,28] = 11.049, P < 0.001). Within-subjects t-tests served as post hoc analyses. These tests indicated that when collapsing across training condition, the mean point of responding during the training phase differed from asymptote, at a statistically significant level (t[15] = 3.90, P < 0.001). Additionally, the asymptote differed from extinction on this dependent measure, at a statistically significant level (t[15] = −3.66, P < 0.002). As seen in , animals responded later in the sequence of signals in the training and extinction phases than during the asymptotic phase DV (M = 2.13, SD = 0.75; M = 1.71, SD = 0.67; M = 2.11, SD = 1.1, respectively).
Figure 6 Main effect, phase of training (DV point of responding).
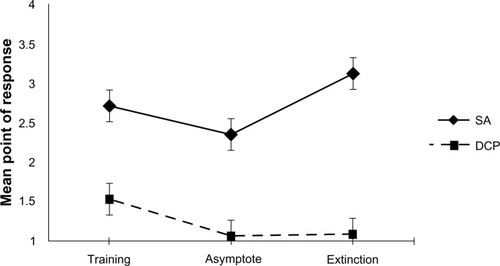
As illustrated in , tests of the interaction of the between-subject factor (training condition) and the within-subject factor (phase of training) were found to be statistically significant (F[1,14] = 10.679, P < 0.001). Test of the simple main effects revealed that animals trained on SA demonstrated a statistically significant difference on this DV between the training and asymptote phases (t[7] = 4.84, P < 0.002), with responding in the asymptote occurring earlier in the sequence of warning signals when compared with training. Additionally, the SA-trained animals demonstrated a statistically significant difference between the asymptote and extinction phases on this DV (t[7] = −7.4, P < 0.01), with responding during extinction occurring later in the sequence of warning signals. Animals trained on DCP, while demonstrating a difference between training and asymptote (t[7] = 2.29, P < 0.05), failed to demonstrate a difference in the point of responding between the asymptote and extinction phases of training (t[7] = −1.14, P > 0.05).
Figure 7 Interaction, training condition × phase of training (DV point of responding).
Percentage of shock
Tests of the main effect for the within-subject factor, phase of training, were found to be statistically significant (F[2,28] = 9.102, P = 0.001). Within-subjects t-tests served as post hoc analyses. These tests indicated that when collapsing across training condition, the percentage of shocks received during the training phase differed from the asymptote at a statistically significant level (t[15] = 2.88, P < 0.01). Additionally, the asymptote differed from extinction on this dependent measure, at a statistically significant level (t[15] = −3.72, P < 0.01). As seen in , animals received a higher percentage of shocks in training and in extinction than during the asymptotic phase DV (M = 24.01, SD = 6.32; M = 9.97, SD = 3.58; M = 27.97, SD = 7.25, respectively).
Figure 8 Main effect, phase of training (DV, % of shock).
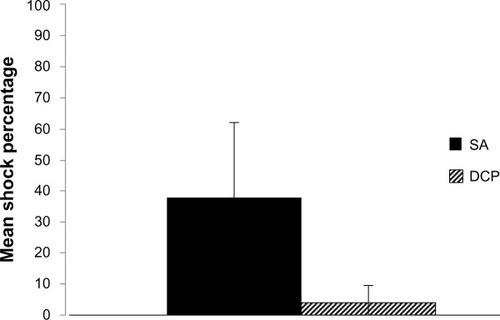
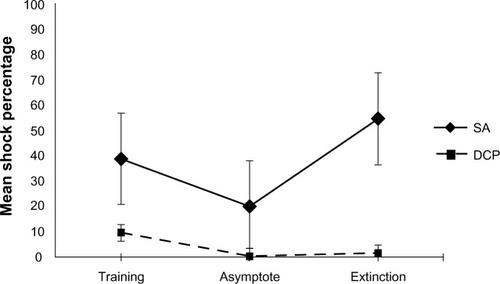
Tests of the main effect for the between-subject factor, training condition, were found to be statistically significant (F[1,14] = 62.462, P < 0.001). Inspection of the means, as demonstrated in , indicated that animals trained under the DCP paradigm were shocked at a lower percentage than those trained under the SA paradigm (M = 3.65, SD = 2.65; M = 37.65, SD = 14.54, respectively).
Figure 9 Main effect, training condition (DV, % of shock).
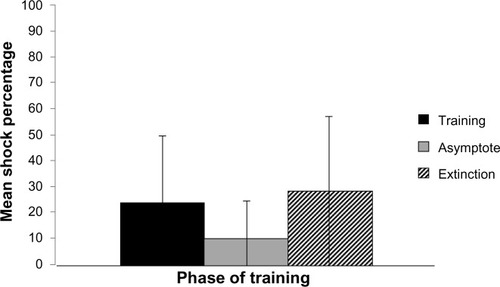
As illustrated in , tests of the interaction of the between-subject factor, training condition, and the within-subjects factor, phase of training, were found to be statistically significant (F[1, 14] = 7.720, P < 0.002). Test of the simple main effects revealed that animals trained on SA demonstrated a statistically significant difference between the training and asymptote phases on this DV (t[7] = 2.02, P < 0.05), with the percentage of shock decreasing from training to the asymptotic phase. Additionally, the SA trained animals demonstrated a statistically significant difference on this DV, between the asymptote and extinction phases (t[7] = −8.34, P < 0.001), with shock percentage during extinction being larger than during the training phase. Animals trained on DCP, while demonstrating a difference between training and asymptote (t[7] = 3.45, P < 0.05), failed to demonstrate a difference in shock percentage between the asymptote and extinction phases of training (t[7] = −1.09, P > 0.05).
Figure 10 Interaction, training condition × phase of training (DV, % of shock).
Discussion
Criticisms of conditioning models of phobia are nearly as old as the history of the models themselves.Citation2 Fear conditioning models of phobia focusing on SA have repeatedly failed to demonstrate a reliable fear of warning signals, a substantial factor expected in an accurate model of phobia.Citation6,Citation8,Citation9 The current study provides support for this criticism. Animals trained under a sequential SA paradigm failed to respond early in the sequence. As illustrates, when collapsing across the phase of training, animals under this paradigm typically responded late in the sequence of warning signals (M = 2.73), failing to demonstrate fear at the earlier warning signals. demonstrates that this trend continued across the training phases; during the asymptotic phase of training, animals responded late in the sequence (M = 2.35). In comparison, animals trained under the DCP model responded much earlier in the sequence. When collapsing across the phase of training, DCP trained animals responded (switching from the chain to the safe lever and receiving reinforcement) to the warning signals between the first and second warning signal (M = 1.23). This difference is even more apparent when one looks at responding during asymptotic performance. shows DCP animals during the asymptotic phase of training responding almost perfectly to the first warning signal (M = 1.06). When animals encountered the first warning signal (steady light) on the chain side, they chose to switch to the second lever, responding an additional 20 times for food. The demonstration of early responding to warning signals suggests that the DCP paradigm may answer a long-held criticism of models of phobia and supports that it may be a better means of representing the complexities of human phobia.
A feature of human phobia worth noting is that phobics are extremely effective in limiting their exposure to the fear-eliciting stimuli.Citation8 This was clearly demonstrated by the DCP-trained animals, as they were highly successful in limiting exposure to shock. During the asymptotic phase of training, DCP trained animals were nearly perfect in preventing shock exposure (M = 0.26%). The SA animals, though effective, were shocked much more frequently – during the asymptotic phase of training, the SA trained animals were shocked in nearly one fifth of trials (M = 19.68%). SA trained animals’ failure to prevent shock as successfully as the DCP animals serves as another example where the proposed model appears to better simulate human phobia.
Phobias have long been described as being extremely resistant to extinction.Citation15 Humans with a phobia continue to demonstrate fear of phobic stimuli (US) even when they have not been exposed to them for extended periods of time. The DCP and SA paradigms were compared regarding their resistance to extinction as a further test of the ability of each model to represent human phobia. Comparison of responding during the asymptotic phase and extinction phase revealed that animals trained under SA demonstrated extinction. demonstrates that the point of responding for SA animals drifted during extinction, with animals responding after the third warning signal on average (M = 3.13). Additionally, as seen in , failure to respond at all increased from nearly 20% of the trials to over 50% (M = 54.61). The change in both of these variables across the asymptotic and extinction phases demonstrates that when shock (US) was removed from the contingency, the animal response rates demonstrated extinction. However, the DCP animals demonstrated a much greater resistance to extinction. and indicate that the DCP animals showed very little change in responding following the removal of shock (US) from the contingency. Indeed, the change in both the point of responding and the percentage of shock received failed to demonstrate a statistically significant difference (t[7] = −1.14, P > 0.05; t[7] = −1.09, P > 0.05, respectively). DCP animals, like phobic humans, continued to respond to warning signals (CS) to prevent exposure to the phobic stimulus even after extended periods in which they failed to be exposed to shock (US).
Dinsmoor proposed that the anomalies often seen in avoidance could best be understood through the lens of punishment and urged researchers to make direct comparisons in studies of avoidance and punishment training.Citation13 Similarly, we suggest that the failure of previous animal models to accurately portray the human phobic scenario is, in part, a result of the confusion between punishment and avoidance that is best illustrated via the direct comparison of the paradigms. Though phobias are often discussed conceptually in the language of punishment, the experimental applications have represented avoidance instead. In avoidance, the absence of a response results in the presentation of the aversive stimuli. This is certainly true of the Stampfl model, in which animals pulled toward shock on a tread mill were forced to perform a behavior (ie, get off the tread mill) to avoid shock.Citation14 More standard avoidance paradigms use lever presses to recreate a similar contingency – shock is presented unless the organism performs a lever press; any behavior other than the lever press, including doing nothing, results in exposure to the aversive stimuli. The distinction between punishment and avoidance becomes more apparent when one considers the “do nothing” test. In punishment, an organism that does not respond is not exposed to warning signals or the aversive stimuli. Indeed, it is the behavior itself that results in the presentation of the aversive stimuli. By failing to do anything at all, the organism can successfully prevent the aversive stimuli. It is important to note, returning to the bridge example, that the person also fails to be reinforced for reaching their destination. The failure to perform a behavior, preventing exposure to aversive stimuli, is often referred to as “passive avoidance,” when in fact, it perfectly describes punishment. We maintain that reference to a behavior not performed as passive avoidance has, in part, potentiated the failure to recognize the role of the fear-eliciting stimuli as punishers of otherwise adaptive behavior. This reconceptualization provides new avenues into investigating the means by which phobias may develop and be maintained. Azrin and Holz summarized data showing that responses reinforced by food will be suppressed by a response-contingent shock, with the extent of the suppression being a function of the aversiveness, reliability, and immediacy of the shock.Citation16 Azrin and Holz also reported that when a brief electric shock punished one of the two alternative responses for producing food, responding was quickly restricted to the unpunished alternative.Citation16 Not surprisingly, given the option between a punished or nonpunished response for food, the nonpunished option is preferred. But what of warning signals predicting the punishment? The present study lends support to the notion that warning signals associated with shock via classical conditioning become aversive punishers themselves. Hake and Azrin showed that as long as the warning signal occasionally predicted shock, the warning signal could be used to punish a single rewarded response, with the amount of suppression being a function of the shock intensity paired with the warning signal.Citation17 To date, no one has reported the role of conditioned punishers in a concurrent schedule where CSs were used to punish one of two responses, both of which produced food. The DCP model demonstrates that in such a scenario, the conditioned punishers successfully suppress the punished alternative and increase responding on the nonpunished alternative.
The results of the current study appear to lend support to a reformulation of learning models of phobia. The DCP model of phobias appears to answer many of the criticisms and shortcomings of the SA models. Animals trained under a DCP paradigm demonstrate highly successful responding to prevent the presentation of shock. Additionally, animals respond early in a sequence of warning signals, providing evidence that warning signals themselves become fear evoking. Additionally, both of these effects appear to be highly resistant to extinction, remaining after the possibility of shock has been removed.
Aside from answering some of the experimental criticisms raised, the model additionally addresses a long-held conceptual problem with avoidance models of phobia. Whereas human phobias are viewed as maladaptive, interfering with healthy daily functioning, traditional active avoidance responding is an adaptive response. Animals trained to lever press to prevent the shock are behaving in an adaptive way. In this very basic conceptual way, avoidance models are doomed to fail in representing phobias. Animals trained under the DCP paradigm are potentially prevented from an adaptive behavior (food reinforcement) as a result of their conditioned fear. In human phobias, it is the fear-induced failure to respond, (eg, leave the house for work, fly to visit family, cross a bridge to go on vacation) that interferes with the pursuit of reinforcement in aspects of one’s life. As noted previously, CostelloCitation6 argued that a phobia model must reflect the detrimental nature of the behavior manifested by the organism. That detriment may manifest in a person needing to work harder to reach the same goal. For instance, a person scared to fly may have to drive 10 hours to get to their vacation spot. This additional work can be seen as detrimental even though the goal has been achieved. The DCP model provides a potential avenue for investigating this aspect of human phobia as well. Animals that chose to switch from the chain to the safe response lever following the occurrence of a warning signal were forced to do “more work” to receive reinforcement. For example, a rat responds five times on the chain side, eliciting the first (steady light) warning signal. Now, switching to the second lever, an additional 20 or 25 total responses, are required to receive reinforcement. Though a fairly modest amount of additional effort is required under the current paradigm, future work that experimentally manipulates the “work requirement” for switching to the safe response will investigate this aspect of the maladaptive nature of phobias.
By reformulating the conceptual and empirical model of phobia, a number of experimental questions can be better addressed. Further investigations into the variables that predict responding in DCP, such as shock magnitude, ratios of reinforcement, and reliability of warning signals, are all important areas to be addressed in future research. Additionally, the effects of anxiolytic medications and physiological measures of fear provide important avenues for further analysis of discriminated punishment as a model of human phobia.
DCP and SA result in very different behavioral profiles during both acquisition and extinction. Viewing phobias in light of these differences may lead to innovations in the treatment of this often debilitating condition.
Acknowledgments
We extend our sincere thanks to Nicole Eslinger, Ekaterina Protsenko, Kaitlyn Dahlborg, Sharon Longo, and Nick Salem for their help in data collection and manuscript preparation. Research reported in this Neuropsychiatric Disease and Treatment, Volume 9, was supported by an Institutional Development Award (IDeA) from the National Institute of General Medical Sciences of the National Institutes of Health und grant number 8 P20 GM103430-12.
Disclosure
The authors report no conflicts of interest in this work.
References
- KesslerRCBerglundPDemlerOJinRMerikangasKRWaltersEELifetime prevalence and age-of-onset distributions of DSM-IV disorders in the National Comorbidity Survey ReplicationArch Gen Psychiatry200562659360215939837
- MinekaSThe role of fear in theories of avoidance learning, flooding, and extinctionPsychol Bull19798659851010
- WatsonJBRaynerRConditioned emotional reactionsJ Exp Psychol192031114
- MaiaTVTwo-factor theory, the actor-critic model, and conditioned avoidanceLearn Behav2010381506720065349
- MowrerOHOn the dual nature of learning: a re-interpretation of “conditioning” and “problem-solving.”Harv Educ Rev194717102148
- CostelloCGDissimilarities between conditioned avoidance responses and phobiasPsychol Rev19707732502544916729
- SidmanMBorenJJThe relative aversiveness of warning signal and shock in an avoidance situationJ Abnorm Psychol195755333934413474911
- MinekaSAnimal models of anxiety-based disorders: their usefulness and limitationsHussainTAMaserJDAnxiety and the Anxiety DisordersHillsdale, NJLawrence Erlbaum Associates1985199244
- StampflTGTheoretical implications of the neurotic paradox as a problem in behavior theory: an experimental resolutionBehav Anal198710216117322477974
- BoydTLLevisDJThe effects of single-component extinction of a three-component serial CS on resistance to extinction of the conditioned avoidance responseLearn Motiv197674517531
- LevisDJBoydTLSymptom maintenance: an infrahuman analysis and extension of the conservation of anxiety principleJ Abnorm Psychol1979882107120571878
- KrasnegorNABradyJVFindleyJDSecond-order optional avoidance as a function of fixed-ratio requirementsJ Exp Anal Behav197115218118716811498
- DinsmoorJAPunishment: I. The avoidance hypothesisPsychol Rev1954611344613134415
- StampflTGAnalysis of aversive events in human psychopathology: fear and avoidanceDennyMRFear, Avoidance, and Phobias: A Fundamental AnalysisHillsdale, NJLawrence Erlbaum Associates1991363394
- McNallyRJPreparedness and phobias: a reviewPsychol Bull198710122833033562708
- AzrinNHHolzWCPunishmentHonigWKOperant Behavior: Areas of Research and ApplicationEnglewood Cliffs, NJPrentice Hall1966380447
- HakeDFAzrinNHConditioned punishmentJ Exp Anal Behav1965827929314342363