
Abstract
Lung cancer has the highest morbidity and mortality in the world, and non-small cell lung carcinomas (NSCLC) account for 80% of cases of lung cancer. The mechanism of NSCLC is still largely unknown, and finding novel targets is of great importance for the treatment of NSCLC. The current study was designed to evaluate the role of miR-148a in NSCLC cell proliferation and invasion and to investigate the possible molecular mechanisms. We found that miR-148a expression was decreased in NSCLC tissues and cell lines. Upregulation of miR-148a significantly decreased A549 cell proliferation, and downregulation of miR-148a significantly increased A549 cell proliferation. Upregulation of miR-148a markedly increased apoptotic cell death and inhibited cell invasion potential. Upregulation of miR-148a significantly decreased signal transducer and activator of transcription 3 (STAT3) expression and 3′-untranslated region luciferase activity. Downregulation of miR-148a significantly increased STAT3 expression. Overexpression of STAT3 significantly inhibited the effect of miR-148a on cell viability and invasion potential. In conclusion, we found that miR-148a inhibited NSCLC cell proliferation and invasion potential through the inhibition of STAT3. Our findings highlight miR-148a/STAT3 axis as a novel therapeutic target for the inhibition of NSCLC growth.
Introduction
It is well known that lung cancer has the highest morbidity and mortality in the world.Citation1,Citation2 It has been reported that lung cancer resulted in 160,000 deaths in the United States in 2014, which accounted for 20% of all cancer-caused deaths.Citation3 Among those, ~80% of lung cancers are classified as non-small cell lung carcinomas (NSCLC) with a fairly low survival rate (5-year survival rate is less than 15%).Citation3–Citation5 Adenocarcinoma and squamous cell carcinoma are the main types of NSCLC.Citation6 Despite great breakthrough in the development of therapeutic techniques, patients with advanced NSCLC have a poor prognosis, and the 5-year survival rate of patients with NSCLC remains poor.Citation7,Citation8 The mechanism of NSCLC is still largely unknown, and finding novel targets is of great importance for the treatment of NSCLC.Citation9
MicroRNAs (miRNAs or miRs) are a class of small non-coding RNAs (20–24 nucleotides) that have been implicated in various biological processes and diseases.Citation10,Citation11 miRNAs function to modulate protein expression through mRNA destabilization and degradation via directly binding the 3′-untranslated region of the target mRNA.Citation12,Citation13 One-third of all mammalian genes have been estimated to be directly or indirectly regulated by miRNAs. It is reported that miRNAs play a role in the regulation of cell growth, proliferation, apoptosis, differentiation, migration, metabolism, etc.Citation14 In particular, dysregulation of miRNAs is believed to function as an important regulator in the pathogenesis of various types of tumors.Citation15,Citation16 Investigation of miRNAs in the development of tumor is useful for diagnosis and therapy of cancer.Citation17 Numerous miRNAs have been found to be dysregulated under the pathological condition of NSCLC.Citation18,Citation19 Recently, studies have found that miR-148a functions to suppress NSCLC.Citation20–Citation22 However, the mechanism underlying the inhibitory effect of miR-148a on NSCLC is still not completely known.
The present study was designed to investigate the mechanism underlying the antitumor effect of miR-148a in NSCLC. We found that miR-148a expression was decreased in NSCLC tissues and cell lines and functioned to suppress NSCLC cell proliferation and invasion through inhibition of signal transducer and activator of transcription 3 (STAT3).
Materials and methods
Chemicals and materials
β-actin was purchased from Santa Cruz Biotechnology Inc. (Santa Cruz). Bax, Bcl-2, STAT3, E-cadherin, and Snail antibodies were obtained from Cell Signaling Technology (CST; Danvers, MA, USA). All of the other chemicals used were of the highest grade available commercially.
Ethics statement and clinical specimens
The study was approved by the Ethical Review Committee of Shanxi Provincial People’s Hospital and complied with the Declaration of Helsinki. Twenty-one cases of NSCLC were retrieved from the Pathology Archive of Shanxi Provincial People’s Hospital in 2015. All patients provided their written informed consent. The NSCLC was diagnosed by two independent pathologists according to the World Health Organization classification. Clinical specimens were collected when the surgery was conducted in patients who had not received any chemotherapy or radiotherapy before surgery.
Cell culture
Normal human bronchial epithelial (HBE) and NSCLC cell lines (A549, H1299, H1650, and H460) were purchased from the American Type Culture Collection (Manassas, VA, USA). Cells were cultured in RPMI-1640 medium supplemented with 10% fetal bovine serum, 100 μg/mL streptomycin, and 100 U/mL penicillin in an incubator with 5% CO2 at 37°C.
Cell transfection
miR-148a mimics, miR-148a inhibitors (anti-miR-148a), and negative controls were synthesized by GeneBioPharma Company (Shanghai, People’s Republic of China). The STAT3 plasmid and empty vectors were purchased from Santa Cruz Biotechnology Inc. (Santa Cruz). Transient transfection was performed using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocols.
RNA isolation and real-time polymerase chain reaction (PCR)
Total RNA was isolated using Trizol reagent (Invitrogen). Total RNA of 500 ng was reversely transcribed using a commercial kit according to the manufacturer’s instructions (Takara, Tokyo, Japan). One microliter of cDNA was used for the performance of real-time PCR with SYBR Green PCR kit (Thermo Scientific, IL, USA). The PCR reaction conditions were as follows: initial denaturation at 95°C for 10 min followed by 30 cycles at 95°C for 1 min, annealing at 53°C for 1 min, extension at 72°C for 1 min, and final extension at 72°C for 5 min. The PCR was conducted using a CFX96 real-time PCR system (BioRad, Hercules, CA, USA). β-actin and U6 small nuclear RNAs were used as internal controls. Results were expressed as folds of control.
Cell proliferation
Cell Counting Kit-8 assay kit was used to determine cell proliferation according to the manufacturer’s protocols. Absorbance at 450 nm was measured using a microplate reader (BioRad).
Luciferase reporter assay
Cells were seeded in 24-well plates and then co-transfected with miR-148a mimics or negative control and the mutant or wild type 3′UTR of STAT3 using Lipofectamine 2000. Forty-eight hours after the transfection, cells were collected, and then relative luciferase activity was measured using the Dual Luciferase Reporter Assay System (Promega, Madison, WI, USA) according to the manufacturer’s protocols.
Western blot
Cells were lysed using RIPA lysis buffer (Thermo Scientific), and proteins were extracted. Protein concentration was determined by bicinchoninic acid (BCA) assay (Thermo Scientific). Twenty micrograms of protein was loaded and separated with 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis. After separation, proteins were transferred onto PVDF membrane (Millipore, MA, USA), and then blocked using 8% nonfat milk. Membranes were incubated overnight at 4°C with primary antibodies (β-actin: Santa Cruz, 1:1000; STAT3: CST, 1:1000; Bax: CST, 1:1000; Bcl-2: CST, 1:1000; E-cadherin: CST, 1:1000; Snail: CST, 1:1000). After four-time washing, the membranes were incubated with a horseradish peroxidase-conjugated secondary antibody (Thermo Scientific) at 37°C for 30 min. Protein bands were visualized with the enhanced chemiluminescence and captured using BioRad Imaging Systems (BioRad).
Apoptosis
After the treatment, cells were collected and apoptosis was determined using TdT-mediated dUTP nick end labeling (TUNEL) assay kit (Roche, Basel, Switzerland) according to the manufacturer’s protocols. Percentage of apoptotic cells were analyzed using flow cytometry. Results were shown as folds of control.
Caspase-3 and caspase-9 activities
After the treatment, cells were collected and pellets were suspended in chilled cell lysis buffer. Colorimetric assay kits were used to analyze the activities of caspase-3 and caspase-9 (BioVision, Milpitas, CA, USA). In brief, cytosolic protein from the cells was extracted and protein concentration was determined by BCA assay (Thermo Scientific). In each assay, 50 μL cell lysis buffer containing 50–200 μg protein was used. At the end, absorbance at 405 nm was measured using a microplate reader (BioRad).
Statistical analysis
The data were shown as the mean ± SEM, and data analysis was performed using SPSS 16.0 software (Chicago, IL). Significance between more than two groups was analyzed using one-way analysis of variance. Significance of differences between two groups was analyzed by Student’s t-test. P<0.05 was considered to be statistically significant.
Results
miR-148a reduced in NSCLC tissues and cell lines
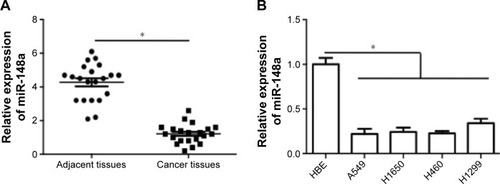
To confirm the expression pattern of miR-148a in NSCLC, we first assessed the expression of miR-148a in NSCLC tissues and cell lines. As shown in , miR-148a expression was reduced in samples of NSCLC tissues, compared with that in the corresponding adjacent normal lung tissues. In addition, in all the NSCLC cell lines, including A549, H1299, H1650, and H460 cell lines, miR-148a expression was markedly lower than that in the HBE cell line ().
Figure 1 Relative expression of miR-148a in non-small cell lung cancer tissues and cell lines.
Abbreviations: HBE, human bronchial epithelial; NSCLC, non-small cell lung carcinomas.
miR-148a reduced cell proliferation in A549 cells
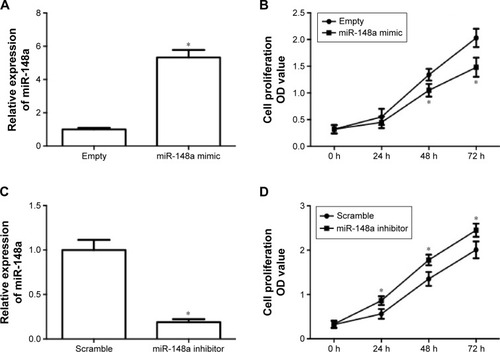
In order to evaluate the role of miR-148a in the growth of NSCLC, the effect of upregulation and downregulation of miR-148a on cell proliferation in A549 cells was examined. A549 cells were transfected with miR-148a mimic or inhibitor and then real-time PCR was conducted to detect the miR-148a expression. In , the results showed that transfection of miR-148a mimic induced a significant increase in miR-148a expression. Upregulation of miR-148a expression significantly suppressed A549 cell proliferation (). In , the results showed that transfection of miR-148a inhibitor induced a significant decrease in miR-148a expression. Downregulation of miR-148a expression significantly increased A549 cell proliferation (). The results demonstrated that miR-148a reduced cell proliferation in A549 cells.
Figure 2 miR-148a reduced cell proliferation in A549 cells.
miR-148a promoted apoptosis in A549 cells
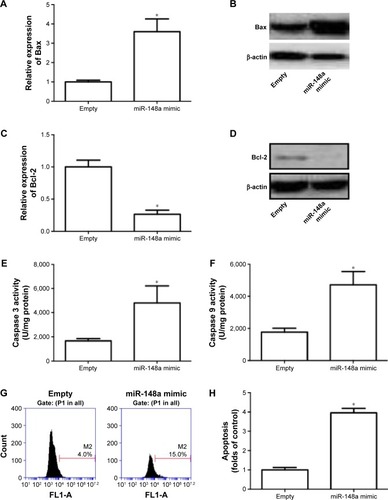
Role of miR-148a in apoptotic cell death was evaluated in A549 cells. In , we showed that transfection of miR-148a mimic significantly increased the mRNA and protein expression of Bax and decreased the mRNA and protein expression of Bcl-2. Moreover, miR-148a mimic notably increased the activities of caspases 3 and 9 (). The percentage of TUNEL-positive cells in A549 cells was significantly increased by upregulation of miR-148a (). The results indicated that miR-148a promoted apoptosis, which may contribute to the decrease of cell proliferation in A549 cells.
Figure 3 miR-148a promoted apoptosis in A549 cells.
Abbreviations: P1, population 1; PCR, polymerase chain reaction.
miR-148a decreased the invasion potential in A549 cells
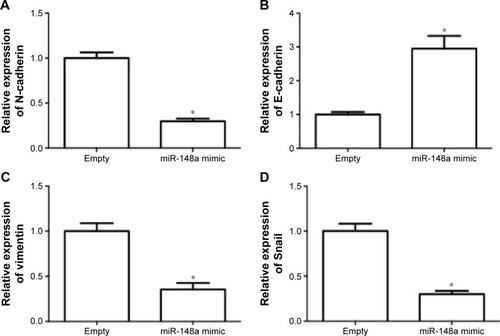
The effect of miR-148a on invasion potential in A549 cells was examined. As shown in , the expression of N-cadherin, vimentin, and Snail was notably reduced by miR-148a mimic. The expression of E-cadherin was markedly increased by miR-148a mimic. The results indicated that miR-148a exhibited an inhibitory effect against invasion potential in A549 cells.
Figure 4 miR-148a decreased the invasion potential in A549 cells.
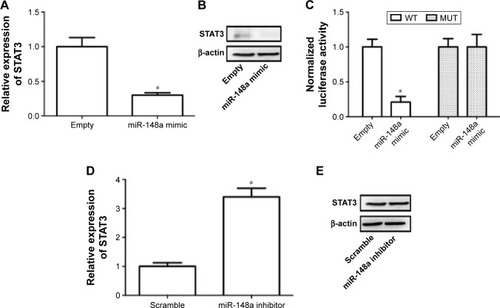
Downregulation of STAT3 required for miR-148a-induced inhibition of cell proliferation and invasion in A549 cells
Previous literature has reported that STAT3 is a potential target of miR-148a in gastric tumor.Citation23 In the current study, we also examined whether regulation of STAT3 was involved in the effect of miR-148a on cell proliferation and invasion in A549 cells. We showed that miR-148a mimic significantly decreased the mRNA and protein expression of STAT3 (). miR-148a mimic markedly decreased the luciferase activity of 3′-untranslated region of STAT3 (). miR-148a inhibitor notably increased the mRNA and protein expression of STAT3 (). The results indicated that STAT3 may play a role in the regulation of proliferation and invasion.
Figure 5 STAT3 was a target gene of miR-148a.
Abbreviations: MUT, mutant; STAT3, signal transducer and activator of transcription 3; WT, wild type.
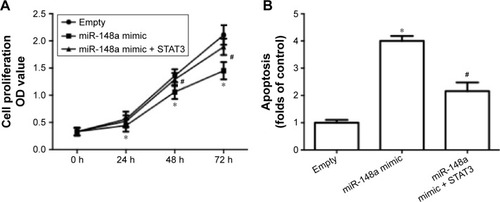
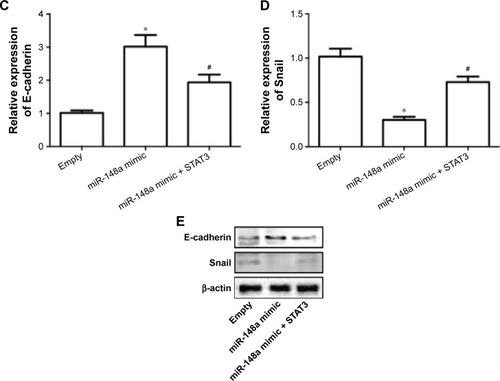
To further confirm the extract role of STAT3, we transfected cells with plasmid carrying STAT3. The results showed that overexpression of STAT3 significantly inhibited miR-148a mimic-induced decrease of cell proliferation (), increase of apoptosis () and E-cadherin expression (), and decrease of Snail expression (). The results indicated that downregulation of STAT3 was involved in miR-148a mimic-induced effect on cell proliferation and invasion potential in A549 cells.
Figure 6 Upregulation of STAT3 inhibited miR-148a mimic-induced inhibition of cell proliferation and invasion in A549 cells.
Abbreviations: CCK-8, Cell Counting Kit-8; PCR, polymerase chain reaction; OD, optical density; STAT3, signal transducer and activator of transcription 3; TUNEL, TdT-mediated dUTP nick end labeling.
Discussion
In the present study, we investigated the role of miR-148a in NSCLC cell proliferation and invasion potential and clarified the possible molecular mechanisms. Previous literature has shown that miR-148a functions to regulate the growth of several types of tumors.Citation24 For example, Xu et al found that miR-148a functioned to suppress metastasis and served as a prognostic indicator in triple-negative breast cancer.Citation20 Downregulation of miR-148a was significantly associated with shorter overall survival of gastric cancer patients.Citation25 In contrast, decreased expression of miR-148a was found to predict poor prognosis in ovarian cancer and to be associated with tumor growth and metastasis.Citation26 miR-148a was reported to serve as predictive biomarker for early diagnosis of laryngeal carcinoma.Citation27 Huang reported that circulating miR-148a expression level in plasma samples of NSCLC patients was decreased.Citation28 Yang et al believed that miR-148a could function as a novel biomarkers in NSCLC screening.Citation29 Moreover, Joshi et al found that miR-148a reduced tumorigenesis and increased TNF-related apoptosis-inducing ligand-induced apoptosis in NSCLC.Citation30 These literatures suggest a potent role of miR-148a in the development of NSCLC. However, the exact role and molecular mechanism of miR-148a-induced regulation of NSCLC is far from completely understood.
In the current study, we found that miR-148a expression was decreased in NSCLC tissues and cell lines. Moreover, we showed that upregulation of miR-148a expression decreased cell proliferation in A549 cells and downregulation of miR-148a expression showed an opposite effect. Furthermore, upregulation of miR-148a expression activated apoptotic pathway and promoted apoptosis. The results demonstrated that miR-148a inhibited A549 cell proliferation and downregulation of miR-148a may be required for the development of NSCLC. Furthermore, the effect of miR-148a on invasion potential was evaluated. The results suggested that miR-148a inhibited invasion potential in A549 cells since upregulation of miR-148a decreased N-cadherin, vimentin, and Snail expression and increased E-cadherin.
In the next step, we examined the potential targets that mediated the antitumor effect of miR-148a in NSCLC. The STAT signaling pathway is one of the most important intracellular signaling networks.Citation31 As one of the family members of STATs, STAT3 transduces signals from activated receptors or intracellular kinases to the nucleus, thus activating and regulating gene transcription.Citation31,Citation32 In response to interleukin-6 signal, STAT3 could be activated and is involved in various types of pathological conditions. STAT3 plays important roles in various types of cancer.Citation33–Citation35 Yu et al found that activated STAT3 was correlated with prognosis of NSCLC.Citation36 Disruption of STAT3 by niclosamide reversed radioresistance of human lung cancer.Citation37 Suppression of JAK/STAT3 was found to exert antitumor activity in NSCLC.Citation38 Moreover, miR-148a was identified to function as a tumor suppressor via inactivating STAT3 in human gastric cancer.Citation23
In the present study, we also examined the role of STAT3 in the antitumor effect of miR-148a in NSCLC. We showed that STAT3 was a target of miR-148a in A549 cells. Upregulation of STAT3 significantly suppressed the effect of miR-148a on cell proliferation and invasion potential. The results indicated that downregulation of STAT3 was required for miR-148a-induced inhibition of cell proliferation and invasion potential in A549 cells.
In conclusion, we found that miR-148a inhibited NSCLC cell proliferation and invasion potential through inhibition of STAT3. Our findings identify miR-148a/STAT3 axis as a novel therapeutic target for the inhibition of NSCLC growth.
Disclosure
The authors report no conflicts of interest in this work.
References
- SiegelRNaishadhamDJemalACancer statistics, 2013CA Cancer J Clin201363113023335087
- SiegelRNaishadhamDJemalACancer statistics, 2012CA Cancer J Clin201262102922237781
- HerbstRSHeymachJVLippmanSMLung cancerN Engl J Med2008359131367138018815398
- ClaassensLvan MeerbeeckJCoensCHealth-related quality of life in non-small-cell lung cancer: an update of a systematic review on methodologic issues in randomized controlled trialsJ Clin Oncol201129152104212021464420
- HuZChenXZhaoYSerum microRNA signatures identified in a genome-wide serum microRNA expression profiling predict survival of non-small-cell lung cancerJ Clin Oncol201028101721172620194856
- ShtivelmanEHensingTMolecular pathways and therapeutic targets in lung cancerOncotarget201451392143324722523
- ScagliottiGVNovelloSAdjuvant therapy in completely resected non-small-cell lung cancerCurr Oncol Rep2003531832512781075
- SandlerAGrayRPerryMCPaclitaxel-carboplatin alone or with bevacizumab for non-small-cell lung cancerN Engl J Med2006355242542255017167137
- SocinskiMASeeking new options for the treatment of small-cell lung cancer. Lung cancer (Amsterdam, Netherlands)Lung Cancer200550Suppl 1S25S26
- RanganathanKSivasankarVMicroRNAs – biology and clinical applications. Journal of oral and maxillofacial pathologyJ Oral Maxillofac Pathol201418222923425328304
- LiMHFuSBXiaoHSGenome-wide analysis of microRNA and mRNA expression signatures in cancerActa Pharmacol Sin201536101200121126299954
- WinterJJungSKellerSGregoryRIDiederichsSMany roads to maturity: microRNA biogenesis pathways and their regulationNat Cell Biol200911322823419255566
- BartelDPMicroRNAs: genomics, biogenesis, mechanism, and functionCell2004116228129714744438
- RottiersVNajafi-ShoushtariSHKristoFMicroRNAs in metabolism and metabolic diseasesCold Spring Harb Symp Quant Biol20117622523322156303
- ZimmermanALWuSMicroRNAs, cancer and cancer stem cellsCancer Lett20113001101920965651
- KishikawaTOtsukaMOhnoMYoshikawaTTakataAKoikeKCirculating RNAs as new biomarkers for detecting pancreatic cancerWorld J Gastroenterol201521288527854026229396
- WangPZhuangLZhangJThe serum miR-21 level serves as a predictor for the chemosensitivity of advanced pancreatic cancer, and miR-21 expression confers chemoresistance by targeting FasLMol Oncol20137333434523177026
- WangXChenZMicroRNA-19a functions as an oncogenic microRNA in non-small cell lung cancer by targeting the suppressor of cytokine signaling 1 and mediating STAT3 activationInt J Mol Med201535383984625604748
- SunWMaYChenPWangDMicroRNA-10a silencing reverses cisplatin resistance in the A549/cisplatin human lung cancer cell line via the transforming growth factor-beta/Smad2/STAT3/STAT5 pathwayMol Med Rep20151153854385925586740
- XuXZhangYJasperJMiR-148a functions to suppress metastasis and serves as a prognostic indicator in triple-negative breast cancerOncotarget2016715203812039426967387
- LiJSongYWangYLuoJYuWMicroRNA-148a suppresses epithelial-to-mesenchymal transition by targeting ROCK1 in non-small cell lung cancer cellsMol Cell Biochem20133801–227728223670799
- ZhengBLiangLWangCMicroRNA-148a suppresses tumor cell invasion and metastasis by downregulating ROCK1 in gastric cancerClin Cancer Res201117247574758321994419
- YuBLvXSuLMiR-148a functions as a tumor suppressor by targeting CCK-BR via inactivating STAT3 and Akt in human gastric cancerPloS One2016118e015896127518872
- LiYDengXZengXPengXThe role of Mir-148a in cancerJ Cancer20167101233124127390598
- QiuXZhuHLiuSExpression and prognostic value of microRNA-26a and microRNA-148a in gastric cancerJ Gastroenterol Hepatol Epub2016816
- GongLWangCGaoYWangJDecreased expression of microRNA-148a predicts poor prognosis in ovarian cancer and associates with tumor growth and metastasisBiomed Pharmacother201683586327470550
- WuYYuJMaYWangFLiuHmiR-148a and miR-375 may serve as predictive biomarkers for early diagnosis of laryngeal carcinomaOncol Lett201612287187827446362
- HuangMXDown-expression of circulating micro ribonucleic acid (miRNA)-148/152 family in plasma samples of non-small cell lung cancer patientsJ Cancer Res Ther201612267167527461630
- YangJSLiBJLuHWSerum miR-152, miR-148a, miR-148b, and miR-21 as novel biomarkers in non-small cell lung cancer screeningTumour Biol20153643035304225501703
- JoshiPJeonYJLaganaAMicroRNA-148a reduces tumorigenesis and increases TRAIL-induced apoptosis in NSCLCProc Natl Acad Sci U S A20151128650865526124099
- KiuHNicholsonSEBiology and significance of the JAK/STAT signalling pathwaysGrowth Factors20123028810622339650
- HeinrichPCBehrmannIMuller-NewenGSchaperFGraeveLInterleukin-6-type cytokine signalling through the gp130/Jak/STAT pathwayBiochem J1998334Pt 22973149716487
- YehCMChangLYLinSHEpigenetic silencing of the NR4A3 tumor suppressor, by aberrant JAK/STAT signaling, predicts prognosis in gastric cancerSci Rep201663169027528092
- XiangMKimHHoVTGene expression-based discovery of atovaquone as a STAT3 inhibitor and anti-cancer agentBlood Epub2016816
- DuanJYueWYuJEIn vitro comparative studies of resveratrol and triacetylresveratrol on cell proliferation, apoptosis, and STAT3 and NFkappaB signaling in pancreatic cancer cellsSci Rep201663167227539371
- YuYZhaoQWangZLiuXYActivated STAT3 correlates with prognosis of non-small cell lung cancer and indicates new anticancer strategiesCancer Chemother Pharmacol201575591792225735252
- YouSLiRParkDDisruption of STAT3 by niclosamide reverses radioresistance of human lung cancerMol Cancer Ther201413360661624362463
- ZhuFDaiCFuYPhysalin A exerts anti-tumor activity in non-small cell lung cancer cell lines by suppressing JAK/STAT3 signalingOncotarget2016789462947626843613