
Abstract
Corilagin is a member of the tannin family and has been isolated from traditional Chinese medicinal plants, such as Phyllanthus spp. Corilagin has anti-inflammatory, antioxidative, antiatherogenic, and antihypertensive effects in various experimental models. In this research, we aimed to investigate for the first time whether corilagin had apoptotic and genomic effects in ovarian cancer treatment in the same study. The potential apoptotic of corilagin was investigated using a WST1 cell proliferation test, caspase 3, and mitochondrial membrane potential JC1 assays in a time- and dose-dependent manner. Genomic changes in expression levels against corilagin treatment were measured using an Illumina human HT-12V4 BeadChip microarray. Bioinformatic data analyses were performed using GenomeStudio and Ingenuity Pathway Analysis software. The data of our study demonstrated that there were statistically significant time- and dose-dependent increases in caspase 3 enzymatic activity and loss of mitochondrial membrane potential in line with decreases in cancer cell proliferation. According to gene-ontology analysis, we found that adherens junctions, antigen processing and presentation, and the phosphatidylinositol signaling system were the most statistically significant networks in response to corilagin treatment on SKOV3 cells, in a time- and dose-dependent manner. The apoptotic and genome-wide effects of corilagin on ovarian cancer cells were examined in detail for the first time in the literature. The results of our study suggest that corilagin might have the potential to be used as a new treatment option for epithelial ovarian cancer.
Introduction
Ovarian cancer is one of the leading causes of cancer-related deaths in women around the world.Citation1,Citation2 Epithelial ovarian cancer accounts for ~90% of ovarian cancer cases, with four major subtypes: endometrioid, clear, mucinous, and serous cell.Citation3,Citation4 According to recent Globocan (2012) statistical data, 238,700 people were diagnosed as having epithelial ovarian cancer and there were 151,900 deaths worldwide.Citation5 Despite various current surgical and therapeutic methods, the 5-year survival ratio of ovarian cancer is nearly 30%–40%, because of the absence of significant symptoms at earlier stages and late diagnosis at advanced or metastatic stages.Citation6–Citation10 In order to improve the overall survival ratio of patients with ovarian cancer, recent studies have targeted the finding of novel, effective, and nontoxic treatment strategies, as well as nutraceuticals, which are physiologically bioavailable and also cost-effective.Citation11
Corilagin belongs to the tannin family, which is extracted from different medicinal plants, including Phyllanthus amarus, P. urinaria, Geranium thunbergii, G. carolinianum, and Dimocarpus longan.Citation12–Citation15 Recent studies have reported that corilagin has beneficial properties in cardiovascular diseases with its antihypertensive, thrombolytic, and antiatherogenic features.Citation16–Citation18 It has also been demonstrated that corilagin can serve as a hepatoprotective radical scavenger for superoxide anions and peroxyls.Citation19 In addition, recent studies have shown that corilagin has antiproliferative, antitumoral, and anti-inflammatory effects.Citation20–Citation23 Although the potential therapeutic use of corilagin has already been identified, the underlying molecular mechanism of corilagin remains unclear.
To the best of our knowledge, there are no data about the genomic effects of corilagin in epithelial ovarian carcinoma. In this study, we aimed to investigate antiproliferative and apoptotic effects of corilagin on epithelial ovarian carcinoma using cell culture methods.
Materials and methods
Cell culture and treatment
SKOV3 cells line (HTB77) commercially purchased from the American Type Culture Collection were cultured in McCoy’s 5A modified medium supplemented with fetal bovine serum (heat-inactivated, 10%) and penicillin–streptomycin (1%) at 37°C in 5% CO2. SKOV3 cells were treated with vehicle 1% dimethyl sulfoxide (DMSO) alone or 5, 10, 25, 50, and 100 µM of corilagin dissolved in 1% DMSO.
Chemicals and reagents
McCoy’s 5A medium, heat-inactivated fetal bovine serum, and penicillin–streptomycin were purchased from Thermo Fisher Scientific (Waltham, MA, USA). Corilagin was obtained from Sigma-Aldrich (St Louis, MO, USA). The WST1 cell proliferation assay was obtained from Hoffman-La Roche (Basel, Switzerland). The caspase 3 assay was purchased from BioVision (Milpitas, CA, USA). The JC1 mitochondrial membrane potential (MMP) detection assay was obtained from Cayman Chemical (Ann Arbor, MI, USA).
Cell viability assay
The cell viability effect of corilagin was determined using the WST1 assay. SKOV3 cells were seeded at a density of 104 cells/well in 96-well plates. After 24 hours’ incubation, cells were treated with 5, 10, 25, 50, and 100 µM corilagin or vehicle (1%, 10 µL DMSO) alone as a control for different time periods (24–72 hours). WST1 colorimetric cell proliferation assays were performed after each incubation period. WST1 solution (10 µL) was added to each well and incubated for 4 hours at 37°C. Color development was measured at 450 nm using a microplate reader (Thermo Fisher Scientific). Cell viability was determined as a percentage of proliferation versus untreated cells (control, 100%).
Caspase 3 enzymatic activity assay
Determination of changes in caspase 3 enzymatic activity is known to be one of the important indicators of apoptosis. We used a caspase 3 colorimetric assay to examine apoptotic changes against corilagin treatment. SKOV3 cells were seeded at a density of 5×105 cells/well in 96-well plates. After a 24-hour incubation period, cells were treated with 50 and 100 µM corilagin or vehicle (1% DMSO) alone as a control for different time periods (48–72 hours). After each incubation period, the cells were lysed by adding 50 µL chilled cell lysis buffer and incubated on ice for 10 minutes before centrifugation. Supernatant samples were then mixed by adding 50 µL reaction buffer, 50 µL sample, and 5 µL DEVD-pNA substrate, and incubated for 2 hours. The reactions were read at 405 nm as described in the previous section.
MMP assay
We used a JC1 assay kit to investigate the loss of MMP in response to corilagin treatment (50 and 100 µM) or vehicle (1% DMSO) for 48–72 hours in SKOV3 cells (Cayman Chemical). In brief, cells (5×105 cells/2 mL) were collected by centrifugation at 1,000 rpm for 10 minutes. After removal of supernatant samples, pellets were homogenized in 200 µL medium, and 20 µL JC1 dye was added to the cells. The cells were then incubated at 37°C for 30 minutes, centrifuged, and 200 µL assay buffer added to the resulting pellets. All pellets were resuspended in 320 µL assay buffer, and 100 µL of each was added to a 96-well plate in triplicate. The plate was read using a fluorescence enzyme-linked immunosorbent assay plate reader. Green/red (485 nm/560 nm) values were calculated to determine changes in MMP.
RNA extraction
Total RNA was isolated from SKOV3 cells in response to 100 µM hesperidin treatment for 72 hours using a “high-pure” RNA isolation kit (Hoffman-La Roche) in accordance with the manufacturer’s instructions. DNA concentrations were qualified with NanoDrop spectrophotometry (260:280 nm ratios), and an A260:A280 ratio between 1.8 and 2 was considered suitable for use.
Microarray hybridization
An HT-12 v4 BeadChip microarray platform of ~47,000 probes was used to investigate whole-genome expression profile changes in response to corilagin treatment (Illumina, San Diego, CA, USA). After conversion of the totally amplified 500 ng total RNA to complementary DNA, fragmentation and biotin-labeling steps were assessed using a TotalPrep RNA Amplification Kit (Thermo Fisher Scientific). After that, the biotin-labeled complementary RNA sample was hybridized to microarray bead chips as per the manufacturer’s protocol. The bead chips were scanned using an Illumina BeadArray Reader. Images of bead chips were converted into signal intensities using Illumina GenomeStudio software. This software was also used to detect the quality controls.
Statistical analysis
Background noise coming from signal intensities of microarray bead chips was corrected and imported into text files using Illumina GenomeStudio. Hierarchical clustering analysis was assessed in data sets to evaluate the proximity between the genes. Hierarchical clusters were constructed using the statistically significant (P<0.05) genes. The related genes were identified as differentially expressed when logarithmic ratios in three different and independent hybridization experiments showed >1.5-fold difference in expression level and when the P-values were <0.05. The Kyoto Encyclopedia of Genes and Genomes, Ingenuity Pathway Analysis (IPA) tools, gene ontology, and pathway analyses were used to ascertain biologic meanings for the differentially expressed genes between the data points.
Results
Corilagin inhibited SKOV3-cell proliferation in a time- and dose-dependent manner
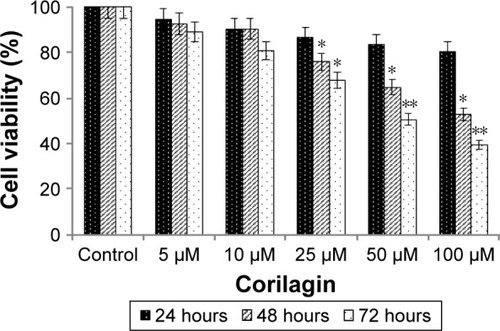
We used the WST1 colorimetric cell proliferation assay to determine cytotoxic effects of corilagin on SKOV3 ovarian cancer cells. The cells were incubated with increasing concentrations (5–100 µM) of corilagin for 24, 48, and 72 hours, and then treated with the WST1 assay after each incubation period. The results showed that 50 and 100 µM corilagin treatment had significant effects in a time-dependent manner (P<0.05, P<0.01). Half-maximal inhibitory-concentration values of corilagin for 24, 48, and 72 hours were calculated from both cell proliferations and found to be 0.7 mM ().
Figure 1 Effects of corilagin on cell proliferation.
Corilagin increased caspase 3 enzymatic activity in a time- and dose-dependent manner
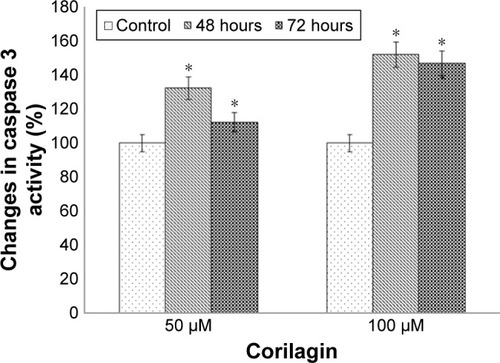
To determine the apoptotic effects of corilagin on SKOV3 cells, cells were incubated with 50 or 100 µM of corilagin for 48–72 hours, and then changes in caspase 3 enzymatic activity were determined. There was a 1.47-fold increase in caspase 3 enzymatic activity in response to 48 hours with 50 µM corilagin. We also found that there was a 2.58-fold increase at 48 hours and a 2.08-fold increase at 72 hours in caspase 3 enzymatic activity in response to treatment with 100 µM corilagin ().
Figure 2 Effects of corilagin on caspase 3 enzymatic activity in SKOV3 cells.
Corilagin induced loss of MMP in a time- and dose-dependent manner
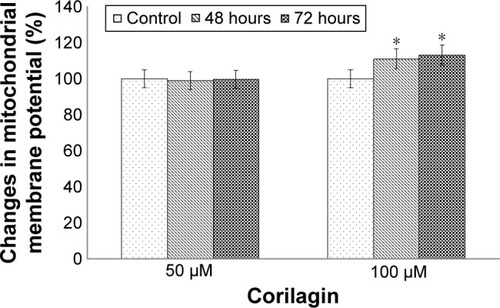
To determine the loss of MMP, cells were incubated with 50 or 100 µM of corilagin for 48–72 hours. The results showed that there was a 1.89-fold increase in the loss of MMP in response to 50 µM corilagin compared with untreated cells. We also found that there was a 2.1-fold increase at 48 hours and 1.89-fold increase at 72 hours in caspase 3 enzymatic activity in response to treatment with 100 µM corilagin ().
Figure 3 Effects of corilagin on membrane potential in SKOV3 cells.
Corilagin induced changes in gene expression profiles
To investigate changes in whole-genome expression levels in response to corilagin treatment (72 hours, 100 µM), we used HT-12 v4 BeadChip microarray profiling. After analyzing the most statistically significant transcripts, we clustered the data based on their expression patterns. In addition, biofunctional annotation analysis was performed to find the downstream pathways in response to corilagin treatment. We also used IPA analysis to understand the relationship between pathways in accordance with the literature ( and ). Based on IPA analysis, three major groups of pathways were shown as statistically significant in corilagin-induced SKOV3 cells: adherens junctions, antigen processing and presentation, and the phosphatidylinositol signaling system ().
Figure 4 Top networks and gene connections.
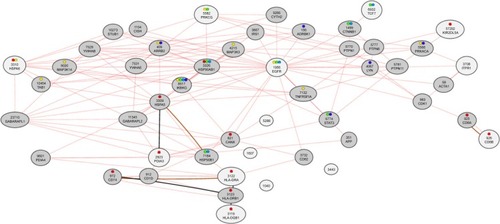
Table 1 Top networks and gene connections
Table 2 Genes involved in top network connections
Discussion
The antiproliferative, apoptotic, and genomic effects of corilagin in epithelial ovarian cancer were examined in detail for the first time in the literature. The results of our study suggest that corilagin has an antitumor effect on ovarian cancer cells and has potential for use as a new treatment for epithelial ovarian cancer.
Hau et al investigated the antitumoral effects of corilagin on HEP3B hepatocellular carcinoma cell lines in an athymic xenograft mouse model. They found significant inhibition in tumor growth in response to corilagin treatment.Citation20 Furthermore, they also indicated that corilagin had no adverse effects on the liver, based on alanine transaminase and aspartate aminotransferase tests. Gambari et al also investigated the sensitization of HEP3B cells to corilagin-induced cisplatin and doxorubicin effects, and showed that using low-dose corilagin could be used in combination with cisplatin and doxorubicin to increase antitumoral effects of chemotherapy treatment.Citation21 In addition to these studies, Ming et al also examined the effects of corilagin on the relationship between antitumoral mechanism and cell cycle proteins using BEL7402 and SMMC7721 hepatocellular carcinoma cells.Citation22 They showed that corilagin-induced SMMC7721 cells arrested at the G2/M phase in response to corilagin via downregulation of the pAkt pathway and upregulation of p53. Together, these data indicate that corilagin has potential for use as a novel antitumoral drug for hepatocellular cancer treatment.Citation22
In addition to its antitumoral effects, two reports emphasized the reparative effects of corilagin for lung injury caused by tobacco products and aerosols. Muresan et al demonstrated that cigarette smoking stimulated the loss of intercellular junctions of CALU3 lung epithelial cells, and corilagin reversed these changes by stimulating expression of connexins.Citation23 Wang et al investigated the effects of corilagin in a pulmonary fibrosis model, and found that high-dose corilagin could downregulate expressions of malondialdehyde, IKKα, pIKKα, NFκB p65, TNFα, and IL-1β to abolish the adverse effects of bleomycin exposure.Citation24 They also showed that TGFβ production and α-SMA expression were inhibited by corilagin treatment. In light of these data, it could be speculated that corilagin could repair lung epithelial injury by affecting cytokine release and the TGFβ1 signaling pathway.Citation24
To the best of our knowledge, no reports have investigated the apoptotic and genomic effects of corilagin on ovarian cancer cells in the same study. However, one report showed the anticarcinogenic effect of corilagin on SKOV3IP, HEY, and HO8910PM ovarian cancer cell lines and a xenograft model using flow cytometry and protein arrays.Citation25 The authors found that corilagin-induced ovarian cancer cells arrested at the G2/M cell cycle point, which correlates with the data from Ming et al. Furthermore, they also showed that levels of cyclin B1, Myt1, and phospho-CDC2 were downregulated in response to corilagin treatment using immunoblotting. In addition, supernatant samples of corilagin-induced ovarian cancer cells have shown inhibition for TGFβ secretion and blockage for TGFβ-induced Snail stabilization. According to that study, corilagin specifically targeted TGFβ secretion to block both canonical SMAD and noncanonical ERK–Akt pathway activation.
In this study, we investigated the apoptotic and genomic effects of corilagin on SKOV3 epithelial ovarian carcinoma cells. We performed WST1, caspase 3, and JC1 assays to understand the dose- and time-dependent effects of corilagin on SKOV3 cancer-cell proliferation and viability. Our results showed correlation with studies that used hepatocellular and ovarian cancer cell lines. We also reported that corilagin-induced inhibition of SKOV3 cell proliferation showed correlation with increased apoptotic cancer cells. In addition, we found that corilagin stimulated the MAPK and phosphatidylinositol signaling system to show its antitumoral effects on downstream pathways. These data show that corilagin has potential anticarcinogenic and antitumoral effects, and thus has potential for use as a novel drug for ovarian cancer treatment. Further studies will help to understand the mechanism of the corilagin-induced effect on cellular metabolism.
Acknowledgments
This work was funded by Istanbul University Scientific Research Project 33202. We thank Mr David F Chapman for editing the manuscript.
Disclosure
The authors report no conflicts of interest in this work.
References
- JemalASiegelRXuJWardECancer statistics, 2010CA Cancer J Clin201060527730020610543
- RenFShenJShiHHornicekFJKanQDuanZNovel mechanisms and approaches to overcome multidrug resistance in the treatment of ovarian cancerBiochim Biophys Acta20161866226627527717733
- CannistraSACancer of the ovaryN Engl J Med2004351242519252915590954
- DengJWangLChenHTargeting epithelial-mesenchymal transition and cancer stem cells for chemoresistant ovarian cancerOncotarget2016734557715578827304054
- TorreLABrayFSiegelRLFerlayJLortet-TieulentJJemalAGlobal cancer statistics, 2012CA Cancer J Clin20156528710825651787
- CakmakogluBAAttarRKahramanOTCyclooxygenase-2 gene and epithelial ovarian carcinoma riskMol Biol Rep20113853481348621107726
- BaldwinLAHuangBMillerRWTen-year relative survival for epithelial ovarian cancerObstet Gynecol2012120361261822914471
- QinDWangWLeiHCDDO-Me reveals USP7 as a novel target in ovarian cancer cellsOncotarget2016747770967710927780924
- VaughanSCowardJIBastRCJrRethinking ovarian cancer: recommendations for improving outcomesNat Rev Cancer2011111071972521941283
- QinDJTangCXYangLHsp90 is a novel target molecule of CDDO-Me in inhibiting proliferation of ovarian cancer cellsPLoS One2015107e013233726134508
- BanerjeeSKayeSBNew strategies in the treatment of ovarian cancer: current clinical perspectives and future potentialClin Cancer Res201319596196823307860
- HauDKGambariRWongRSPhyllanthus urinaria extract attenuates acetaminophen induced hepatotoxicity: involvement of cytochrome P450 CYP2E1Phytomedicine200916875176019386480
- ShenZQDongZJPengHLiuJKModulation of PAI-1 and tPA activity and thrombolytic effects of corilaginPlanta Med200369121109111214750026
- SudjaroenYHullWEErbenGIsolation and characterization of ellagitannins as the major polyphenolic components of longan (Dimocarpus longan Lour) seedsPhytochemistry20127722623722277734
- OkabeSSuganumaMImayoshiYTaniguchiSYoshidaTFujikiHNew TNF-α releasing inhibitors, geraniin and corilagin, in leaves of Acer nikoense, megusurinokiBiol Pharm Bull200124101145114811642320
- DuanWYuYZhangLAntiatherogenic effects of Phyllanthus emblica associated with corilagin and its analogueYakugaku Zasshi2005125758759115997216
- ChengJTLinTCHsuFLAntihypertensive effect of corilagin in the ratCan J Physiol Pharmacol19957310142514298748933
- DuanWGShenZQYanMYunYZhangLYCorilagin, a promising natural product to treat cardiovascular diseasesSinghVKMishraSKGovilJNPhytopharmacology and Therapeutic Values IIHoustonStudium Press2008163172
- KinoshitaSInoueYNakamaSIchibaTAniyaYAntioxidant and hepatoprotective actions of medicinal herb, Terminalia catappa L. from Okinawa Island and its tannin corilaginPhytomedicine2007141175576217293097
- HauDKZhuGYLeungAKIn vivo anti-tumour activity of corilagin on HEP3B hepatocellular carcinomaPhytomedicine2010181111521036022
- GambariRHauDKWongWYChuiCHSensitization of HEP3B hepatoma cells to cisplatin and doxorubicin by corilaginPhytother Res201428578178323913631
- MingYZhengZChenLCorilagin inhibits hepatocellular carcinoma cell proliferation by inducing G2/M phase arrestCell Biol Int201337101046105423686743
- MuresanXMCervellatiFSticozziCThe loss of cellular junctions in epithelial lung cells induced by cigarette smoke is attenuated by corilaginOxid Med Cell Longev2015201563175825802682
- WangZGuoQYZhangXJCorilagin attenuates aerosol bleomycin-induced experimental lung injuryInt J Mol Sci20141569762977924886817
- JiaLJinHZhouJA potential anti-tumor herbal medicine, corilagin, inhibits ovarian cancer cell growth through blocking the TGF-β signaling pathwaysBMC Complement Altern Med2013133323410205