
Abstract
Background
Chemoresistance is one of the major obstacles for cancer therapy in the clinic. Nuclear paraspeckle assembly transcript 1 (NEAT1) has been reported as an oncogene in most malignancies such as lung cancer, esophageal cancer, and gastric cancer. This study is designed to investigate the function of NEAT1 in paclitaxel (PTX) resistance of ovarian cancer and its potential molecular mechanism.
Patients and methods
The expressions of NEAT1 and miR-194 in ovarian cancer tissues and cells were estimated by quantitative real-time polymerase chain reaction (qRT-PCR). MTT, flow cytometry, and Western blot assays were used to assess the effect of NEAT1 on PTX resistance in PTX-resistant ovarian cancer cells. Luciferase reporter assay was applied to examine the association between NEAT1, zinc finger E-box-binding homeobox 1 (ZEB1) and miR-194. Xenograft tumor model was established to confirm the biological role of NEAT1 in PTX resistance of ovarian cancer in vivo.
Results
NEAT1 was upregulated, and miR-194 was downregulated in PTX-resistant ovarian cancer tissues and cells. Functionally, NEAT1 knockdown enhanced cell sensitivity to PTX via promoting PTX-induced apoptosis in vitro. NEAT1 was identified as a molecular sponge of miR-194 to upregulate ZEB1 expression. Mechanistically, NEAT1-knockdown-induced PTX sensitivity was mediated by miR-194/ZEB1 axis. Moreover, NEAT1 knockdown improved PTX sensitivity of ovarian cancer in vivo.
Conclusion
NEAT1 contributed to PTX resistance of ovarian cancer cells at least partly through upregulating ZEB1 expression by sponging miR-194, elucidating a novel regulatory pathway of chemoresistance in PTX-resistant ovarian cancer cells and providing a possible long noncoding RNA (lncRNA)-targeted therapy for ovarian cancer.
Keywords:
Introduction
Ovarian cancer is one of the most prevalent and deadly types of gynecological malignancy, with ~225,000 new diagnosed cases and an estimated 140,000 deaths worldwide.Citation1,Citation2 Due to untypical early symptoms and unreliable screening approaches, the majority of ovarian cancer patients are diagnosed at an advanced stage and present a poor response to currently available therapeutic interventions, with a 5-year survival rate of 15%–30%.Citation3,Citation4 Most ovarian cancer patients receive successful chemotherapy initially; however, tumor recurrence frequently occurs following the development of multidrug resistance (MDR).Citation5 Resistance to paclitaxel (PTX), a frontline chemotherapeutic agent with long-term clinical applications in the therapy of ovarian carcinoma, continues to be one of the primary causes of treatment failure.Citation6 Therefore, it is imperative to identify an effective therapeutic target that can sensitize ovarian cancer to PTX and elucidate the molecular mechanism of drug resistance in ovarian cancer.
Long noncoding RNAs (lncRNAs) are defined as a class of transcripts of >200 nucleotides in length and possess the capability of regulating gene expression at the transcriptional, posttranscriptional, and epigenetic level. It is becoming increasingly apparent that dysregulated lncRNAs are implicated in carcinogenesis and progression of numerous cancers, acting as either oncogenes or tumor suppressors.Citation7 Moreover, abnormality of lncRNA levels has been shown to be associated with the development of chemoresistance in various tumors, including ovarian cancer.Citation8,Citation9 For example, enrichment of lncRNA ROR promoted proliferation, migration, and chemoresistance in nasopharyngeal carcinoma.Citation10 Upregulation of lncRNA HOTTIP induced cellular resistance to cisplatin in osteosarcoma through activating the Wnt/β-catenin pathway.Citation11 LncRNA-TUG1 mediated cell growth and drug resistance of small cell lung cancer by regulating LIMK2b via EZH2.Citation12
Nuclear paraspeckle assembly transcript 1 (NEAT1), transcribed from the familial tumor syndrome multiple endocrine neoplasia type 1 locus on chromosome 11, has been identified as a nuclear-restricted lncRNA with the following two isoforms: 3.7 kb NEAT1-1 and 23 kb NEAT1-2.Citation13 NEAT1 has been documented as an oncogene to facilitate tumorigenesis in different types of solid tumors.Citation14 Of note, NEAT1 was reported to be upregulated in ovarian cancer and contributed to the carcinogenesis of ovarian cancer.Citation15,Citation16 However, the involvement of NEAT1 in PTX resistance of ovarian cancer has been poorly defined.
In recent years, the interaction between lncRNAs and miRNAs has drawn great attention. LncRNAs could function as miRNA sponges or decoys to downregulate the expressions and activities of miRNAs, eventually leading to the derepression of miRNA targets at posttranscriptional level.Citation17 A previous study showed that miR-194 was downregulated in PTX-resistant ovarian cancer cell lines and forced expression of miR-194 attenuated PTX resistance in ovarian cancer cells.Citation18 Hence, this study was designed to explore whether the influence of NEAT1 on PTX resistance is mediated by miR-194 in ovarian cancer.
In this study, we demonstrated that NEAT1 was upregulated and miR-194 was downregulated in PTX-resistant ovarian cancer tissues and cells. Furthermore, functional and mechanism analyses revealed that NEAT1 contributed to PTX resistance through upregulating zinc finger E-box-binding homeobox 1 (ZEB1) expression by sponging miR-194. This study first established a NEAT1-miR-194-ZEB1 regulatory network in ovarian cancer, hinting a promising therapy strategy for ovarian cancer with PTX resistance.
Patients and methods
Patients and tissue samples
This study was approved by the Ethical Committee of Huaihe Hospital of Henan University, and written informed consent was provided by every participant prior to surgery. Thirty-two ovarian carcinoma specimens were collected from ovarian cancer patients receiving oophorectomies between 2013 and 2015 at Huaihe Hospital. All biopsy samples were histopathologically examined by two independent pathologists. Samples were promptly frozen in liquid nitrogen and maintained at −80°C until use. Patient’s samples were divided into two groups based on the response to the first-line chemotherapy: 18 treatment-responsive patients and 14 treatment-resistant patients. According to the NCCN guidelines, intrinsically treatment-resistant tumors were regarded as those with persistent or recurrent disease within 6 months after the initiation of first-line taxol-platinum-based combination chemotherapy. Treatment-responsive tumors were classified as those with a complete response to chemotherapy and a platinum-free interval of >6 months.
Cell lines and culture
Human ovarian carcinoma cell lines SKOV3 and HeyA-8 were obtained from American Type Culture Collection (Manassas, VA, USA). The corresponding PTX-resistant ovarian cancer cells SKOV3/PTX and HeyA-8/PTX cells were established from the parental cell lines by stepwise exposure to escalating concentrations of PTX, as previously described.Citation19 Cells were cultured in RPMI-1640 medium (HyClone, Logan, UT, USA) with 10% (v/v) fetal bovine serum (Thermo Fisher Scientific, Waltham, MA, USA) and antibiotics (100 U/mL penicillin, 100 μg/mL streptomycin) (Sigma-Aldrich Co., St Louis, MO, USA) in a 95% air/5% CO2 atmosphere at 37°C. To maintain the PTX-resistant phenotype of SKOV3/PTX and HeyA-8/PTX cells, 5 nM PTX was additionally added into the culture medium.
Transient transfection
siRNA specially targeting NEAT1 (si-NEAT1), scrambled siRNA control (si-con), miR-194 mimic (miR-194), scrambled miRNA control (miR-con), miR-194 inhibitor (anti-miR-194), and inhibitor control (anti-miR-con) were purchased from RiboBio (Guangzhou, People’s Republic of China). Ectopic expression of NEAT1 was achieved by introducing NEAT1 sequence into pcDNA3.1 vector (Thermo Fisher Scientific). SKOV3/PTX and HeyA-8/PTX cells were seeded into six-well plates at a density of 5×104 cells/well and cultured overnight prior to transfection. Then, transient transfection with oligonucleotides or plasmids into SKOV3/PTX and HeyA-8/PTX cells was performed by Lipofectamine™ 2000 (Thermo Fisher Scientific). Cells were harvested 48 h posttransfection for subsequent analysis.
RNA isolation and quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNA from ovarian cancer tissues and cells was extracted with TRIzol reagent (Thermo Fisher Scientific). The first strand of cDNA for NEAT1 and miR-194 was synthesized from 1 μg of RNA using the High-Capacity cDNA Reverse Transcription Kits (Thermo Fisher Scientific) and MicroRNA Reverse Transcription Kit (Thermo Fisher Scientific), respectively. qRT-PCR was carried out using SYBR® Premix DimerEraser Kit (TaKaRa, Dalian, People’s Republic of China) on an Applied Biosystems 7500 Real-Time PCR System. The relative expressions of NEAT1 and miR-194 were calculated using 2−ΔΔCT method with normalization to GAPDH and U6 snRNA, respectively. The primers used were presented as below: NEAT1: 5′-CTTCCTCCCTT TAACTTATCCATTCAC-3′ (forward primer) and 5′-CTCTTCCTCCACCATTACCAACAATAC-3′ (reverse primer); miR-194: 5′-CAGGAGTTGTAAATCCGAGCCG-3′ (forward primer) and 5′-TTCATAGGTCAGAGCCCT GTGCA-3′ (reverse primer); GAPDH: 5′-TATGATGATATCAAGAGGGTAGT-3′ (forward primer) and 5′-TGTATCCAAACTCATTGTCATAC-3′ (reverse primer); U6: 5′-CTCGCTTCGGCAGCACA-3′ (forward primer) and 5′-AACGCTTCACGAATTTGCGT-3′(reverse primer).
Western blot
Protein samples from tissues or cells were subjected to 10% SDS-PAGE and transferred onto PVDF membranes. Following blocked in 5% skim milk for 2 h, the membranes were incubated overnight at 4°C with the primary antibodies against P-glycoprotein (P-gp) (1:1,000), glutathione S-transferase π (GST-π) (1:500), ZEB1 (1:2,000 dilution), and β-actin (1:5,000) from Santa Cruz Biotechnology Inc. (Dallas, TX, USA), followed by probed with horseradish peroxidase-conjugated goat anti-rabbit second antibody for 2 h at 1:5,000. The antigen–antibody complexes were visualized using chemiluminescence.
Drug resistance assay
PTX resistance was assessed by (4-5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay. SKOV3/PTX and HeyA-8/PTX cells were incubated for 24 h prior to exposing to various doses of PTX for 48 h. Subsequently, 20 μL of 5 mg/mL MTT (Sigma-Aldrich) was added to each well for another 4 h, followed by the addition of 150 μL of DMSO to dissolve the generated formazan crystals. The absorbance at a wavelength of 570 nm was detected using a microplate reader. The concentration of PTX causing 50% inhibition of growth (IC50) was determined by the relative survival curve.
Flow cytometry analysis of apoptosis
SKOV3/PTX and HeyA-8/PTX cells (3.5×105/well) were seeded into six-well plates and treated with PTX for 48 h. The ratio of apoptotic cells was detected using the Annexin V-FITC Apoptosis Detection Kit (Thermo Fisher Scientific) and analyzed using a FACSCalibur flow cytometer with CellQuest software (BD Biosciences, Franklin Lakes, NJ, USA).
Luciferase reporter assay
The NEAT1 full fragments or its mutant containing the putative miR-194-binding sites in NEAT1 were synthesized and cloned into downstream of firefly luciferase gene in pGL3 plasmids (Promega Corporation, Fitchburg, WI, USA), named as pGL3-NEAT1-wild type (Wt) and pGL3-NEAT1-mutant (Mut). Similarly, the 3′UTR of ZEB1 containing the predicted miR-194-binding sites or mutant sites was amplified by PCR and inserted into pGL3 plasmids, named as pGL3-ZEB1-3′UTR-Wt and pGL3-ZEB1-3′UTR-Mut. SKOV3/PTX and HeyA-8/PTX cells were maintained in 96-well plates and cotransfected with 400 ng of constructed luciferase reporter plasmids, 50 ng of renilla luciferase reporter vector (pRL-TK) and 50 nM of miR-194, miR-con, miR-194+ pcDNA-NEAT1 or miR-194+ vector using Lipofectamine™ 2000. Cells were harvested at 48 h after transfection, and luciferase activity was determined by using the Dual Luciferase Reporter Assay Kit (Promega Corporation). Renilla luciferase activities were used as the internal control for the normalization of firefly luciferase activity.
In vivo experiment
Lentiviral vector (Lenti-short hairpin [sh]-NEAT1) for stable NEAT1 overexpression was obtained from Genechem (Shanghai, People’s Republic of China). The animal experiments were approved by the Animal Care and Use Committee of Huaihe Hospital and were performed in accordance with the institutional guide for the care and use of laboratory animals. Six-week-old male BALB/c athymic nude mice (n=5 per group) were purchased from Shanghai Experimental Animal Center (Shanghai, People’s Republic of China). SKOV3/PTX cells (1×107) tansduced with sh-NEAT1 or sh-con were injected subcutaneously into the flank of the nude mice. After 6 days of injection, mice were intraperitoneally administrated with phosphate buffered solution (PBS) or 3 mg/kg PTX (Sigma-Aldrich Co.) every 3 days for five cycles. The tumor size was measured every 3 days with a caliper, and tumor volume was calculated according to the formula: volume = length × width2/2. All mice were sacrificed on day 21 after inoculation. The resected tumor masses were harvested for subsequent weight and qRT-PCR analysis.
Statistical analysis
All quantitative data were shown as mean ± standard deviation (SD) from three independent experiments. The comparisons between two or more groups were estimated by Student’s t-test or one-way analysis of variance (ANOVA). P<0.05 was considered to be statistically significant.
Results
NEAT1 was upregulated and miR-194 was downregulated in PTX-resistant ovarian cancer tissues and cells
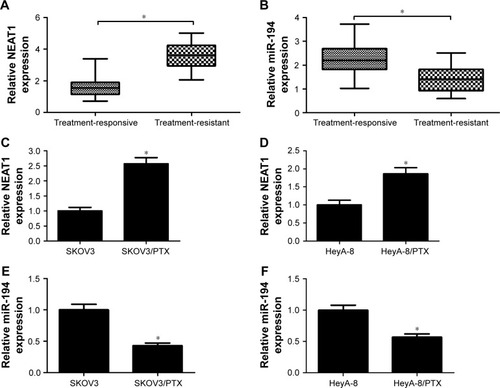
To examine the association of NEAT1 and miR-194 with PTX resistance in ovarian cancer, qRT-PCR was first performed to detect the expressions of NEAT1 and miR-194 in PTX-resistant and PTX-sensitive ovarian cancer tissues. As shown in , a significant increase in NEAT1 expression and a marked decrease in miR-194 expression were observed in treatment-resistant patients compared to that in treatment-responsive patients. Next, the expressions of NEAT1 and miR-194 in PTX-resistant ovarian cancer cells (SKOV3/PTX and HeyA-8/PTX) were further explored. Similarly, higher expression of NEAT1 () and lower expression of miR-194 () were discovered in PTX-resistant ovarian cancer cells in comparison with their parental cells SKOV3 and HeyA-8. These results demonstrated that dysregulation of NEAT1 and miR-194 may be associated with PTX resistance in ovarian cancer.
Figure 1 Expressions of NEAT1 and miR-194 in PTX-resistant ovarian cancer tissues and cells.
Abbreviations: NEAT1, nuclear paraspeckle assembly transcript 1; PTX, paclitaxel; qRT-PCR, quantitative real-time polymerase chain reaction.
NEAT1 knockdown enhanced PTX sensitivity in PTX-resistant ovarian cancer cells
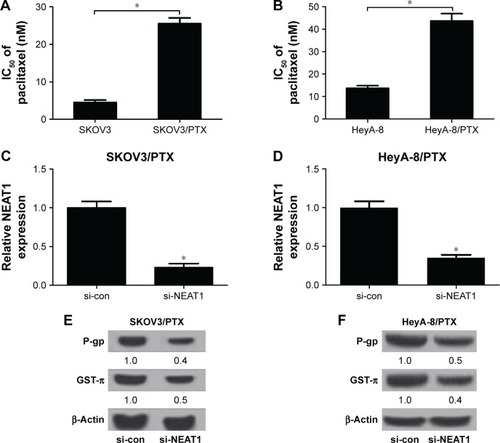
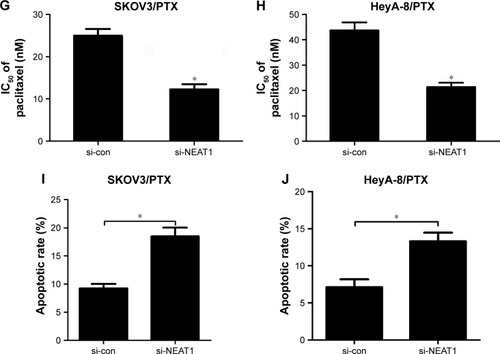
Based on the above results, the effects of NEAT1 knockdown on PTX resistance in ovarian cancer cells were further explored. First, cells were incubated with different concentrations of PTX for 48 h and, then, IC50 value was detected by MTT assay. As expected, IC50 value of PTX in SKOV3/PTX () and HeyA-8/PTX () cells was significantly higher than that in SKOV3 and HeyA-8 cells, confirming the production of PTX resistance in SKOV3/PTX and HeyA-8/PTX cells. Next, the role of NEAT1 in the acquisition of PTX resistance was evaluated by using loss-of-function experiments. qRT-PCR analysis confirmed the knockdown efficiency of NEAT1, demonstrated by the downregulation of NEAT1 expression in si-NEAT1-transfected SKOV3/PTX () and HeyA-8/PTX () cells. Since P-gp and GST-π, as MDR-associated marker proteins, were reported to play an important role in drug resistance of ovarian cancer,Citation20 we investigated the effect of NEAT1 knockdown on the protein levels of P-gp and GST-π by Western blot. The results exhibited that the protein levels of P-gp and GST-π were remarkably reduced in response to transfection of si-NEAT1 in SKOV3/PTX () and HeyA-8/PTX () cells. In addition, IC50 determination showed that NEAT1 knockdown significantly decreased PTX resistance in SKOV3/PTX () and HeyA-8/PTX () cells. To observe whether NEAT1-mediated alteration of PTX resistance was related to apoptosis, SKOV3/PTX and HeyA-8/PTX cells transfected with si-con or si-NEAT1 were treated with PTX for 48 h. Flow cytometry analysis revealed that PTX-induced apoptosis was obviously enhanced after introduction with si-NEAT1 in SKOV3/PTX () and HeyA-8/PTX () cells. Taken together, these data suggested that NEAT1 knockdown sensitized SKOV3/PTX and HeyA-8/PTX cells to PTX.
Figure 2 NEAT1 knockdown-sensitized SKOV3/PTX and HeyA-8/PTX cells to PTX.
Abbreviations: GST-π, glutathione S-transferase π; IC50, the concentration of PTX causing 50% inhibition of growth; MTT, (4-5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide; NEAT1, nuclear paraspeckle assembly transcript 1; P-gp, P-glycoprotein; PTX, paclitaxel; qRT-PCR, quantitative real-time polymerase chain reaction; si-con, siRNA control.
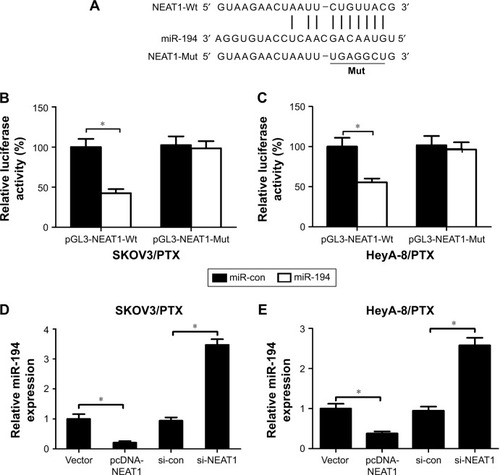
NEAT1 negatively regulated the expression of miR-194
IncRNAs have been reported as competing endogenous RNAs (ceRNAs) that share miRNAs recognition sites with mRNAs or compete for miRNAs binding, thus resulting in changes of target mRNAs expression.Citation21 Through web-based tool Starbase 2.0 (http://starbase.sysu.edu.cn/mirLncRNA.php), miR-194 was predicted to have complementary bases pairing with NEAT1 (). Accordingly, luciferase reporter vectors (pGL3-NEAT1-Wt or pGL3-NEAT1-Mut) containing the wild-type or mutated miR-194-binding sites in NEAT1 were established and co-transfected with miR-194 mimic or miR-con into SKOV3/PTX and HeyA-8/PTX cells. The results showed that ectopic expression of miR-194 dramatically suppressed the luciferase activity of pGL3-NEAT1-Wt reporter but not that of pGL3-NEAT1-Mut reporter in SKOV3/PTX () and HeyA-8/PTX () cells. To further evaluate the regulatory impact between NEAT1 and miR-194, SKOV3/PTX and HeyA-8/PTX cells were transfected with pcDNA-NEAT1, si-NEAT1, or matched controls. It was observed that miR-194 expression was significantly downregulated in pcDNA-NEAT1-transfected SKOV3/PTX and HeyA-8/PTX cells and was exceptionally upregulated in si-NEAT1-transfected SKOV3/PTX and HeyA-8/PTX cells (). All these data indicated that NEAT1 suppressed the expression of miR-194.
Figure 3 NEAT1 inhibited miR-194 expression.
Abbreviations: miR-con, miRNA control; NEAT1, nuclear paraspeckle assembly transcript 1; PTX, paclitaxel; qRT-PCR, quantitative real-time polymerase chain reaction; si-con, siRNA control; Wt, wild type; Mut, mutant.
NEAT1 knockdown improved PTX sensitivity in PTX-resistant ovarian cancer cells by negatively regulating miR-194
In view of the regulatory role of NEAT1 in miR-194 expression in SKOV3/PTX and HeyA-8/PTX cells, whether the influence of NEAT1 in PTX resistance was correlative with miR-194 was further explored. SKOV3/PTX and HeyA-8/PTX cells were transfected with miR-194, si-NEAT1, si-NEAT1 + anti-miR-194, or matched controls. Western blot analysis demonstrated that miR-194 overexpression or NEAT1 knockdown led to an obvious reduction of P-gp and GST-π expressions in SKOV3/PTX () and HeyA-8/PTX () cells, while anti-miR-194 transfection significantly reversed si-NEAT1-mediated reduction of P-gp and GST-π expressions. Drug resistance assay showed that miR-194 upregulation or NEAT1 deficiency effectively enhanced PTX sensitivity in SKOV3/PTX () and HeyA-8/PTX () cells; however, anti-miR-194 introduction greatly abolished si-NEAT1-triggered increase in PTX sensitivity. Meanwhile, flow cytometry analysis revealed that forced expression of miR-194 or NEAT1 silence dramatically promoted PTX-induced apoptosis in SKOV3/PTX () and HeyA-8/PTX () cells, whereas anti-miR-194 treatment markedly abated the promotive effect of si-NEAT1 on PTX-induced apoptosis. These results illustrated that miR-194 downregulation partially overturned NEAT1-knockdown-induced PTX sensitivity in PTX-resistant ovarian cancer cells.
Figure 4 NEAT1 knockdown increased PTX sensitivity in PTX-resistant ovarian cancer cells by repressing miR-194.
Abbreviations: GST-π, glutathione S-transferase π; IC50, the concentration of PTX causing 50% inhibition of growth; MTT, (4-5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide; miR-con, miRNA control; NEAT1, nuclear paraspeckle assembly transcript 1; P-gp, P-glycoprotein; PTX, paclitaxel; si-con, siRNA control.
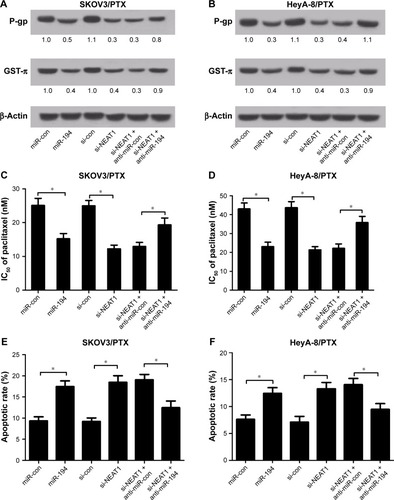
NEAT1 upregulated ZEB1 expression by sponging miR-194
TargetScan 6.2 (www.targetscan.org) and miRanDa (www.microrna.org) were used to predict the potential targets of miR-194. ZEB1-3′UTR was found to contain the complementary sequence for miR-194, as depicted in . To confirm the prediction, the 3′UTR sequence of ZEB1 and its mutated sequence were cloned into the luciferase reporter vectors, named as pGL3-ZEB1-3′UTR-Wt and pGL3-ZEB1-3′UTR-Mut, respectively. Luciferase reporter assay results showed that exogenetic expression of miR-194 significantly blocked luciferase activity of pGL3-ZEB1-3′UTR-Wt reporter, whereas NEAT1 overexpression remarkably abrogated the inhibitory effect in SKOV3/PTX () and HeyA-8/PTX () cells. However, the luciferase activity in pGL3-ZEB1-3′UTR-Mut reporter was not affected by any treatment (). Subsequently, we assessed the protein level of ZEB1 in SKOV3/PTX and HeyA-8/PTX cells introduced with si-NEAT1, miR-194, miR-194+ pcDNA-NEAT1, or matched controls. As shown in , NEAT1 knockdown or miR-194 overexpression markedly suppressed the protein level of ZEB1, while revert of NEAT1 expression attenuated miR-194-induced decrease in ZEB1 expression. Taken together, these data demonstrated that NEAT1 positively regulated ZEB1 expression through miR-194.
Figure 5 NEAT1 upregulated ZEB1 expression via sponging miR-194.
Abbreviations: miR-con, miRNA control; NEAT1, nuclear paraspeckle assembly transcript 1; PTX, paclitaxel; si-con, siRNA control; ZEB1, zinc finger E-box-binding homeobox 1; Wt, wild type; Mut, mutant.
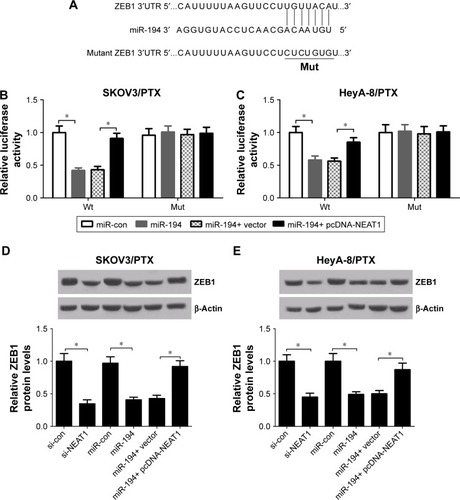
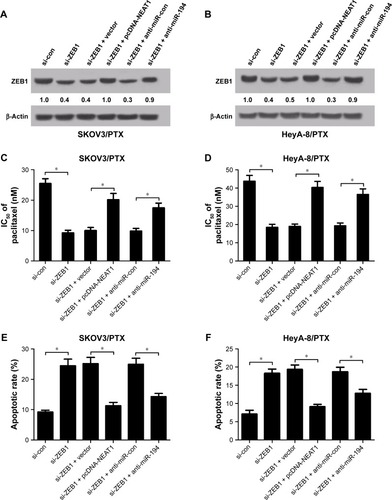
NEAT1 contributed to PTX resistance of PTX-resistant ovarian cancer cells through elevating ZEB1 expression by sponging miR-194
To investigate the regulatory effect of NEAT1 and miR-194 on ZEB1 function, SKOV3/PTX and HeyA-8/PTX cells were transfected with si-ZEB1, si-ZEB1 + pcDNA-NEAT1, si-ZEB1 + anti-miR-194, or matched controls. The protein level of ZEB1 was apparently reduced in si-ZEB1-treated SKOV3/PTX () and HeyA-8/PTX () cells, while NEAT1 overexpression or miR-194 downregulation weakened this effect. IC50 determination showed that ZEB1 knockdown dramatically attenuated PTX resistance in SKOV3/PTX () and HeyA-8/PTX () cells, which was significantly relieved by reintroduction of pcDNA-NEAT1 or anti-miR-194. Furthermore, ZEB1 knockdown resulted in a marked promotion of PTX-induced apoptosis in SKOV3/PTX () and HeyA-8/PTX () cells, which was strikingly suppressed by NEAT1 up-regulation or miR-194 suppression, as demonstrated by flow cytometry analysis. These results indicated that NEAT1 overexpression or miR-194 downregulation reversed ZEB1-knockdown-induced sensitivity of PTX in ovarian cancer cells.
Figure 6 ZEB1-knockdown-induced sensitivity of PTX was suppressed by reintroduction of pcDNA-NEAT1 or anti-miR-194.
Abbreviations: IC50, the concentration of PTX causing 50% inhibition of growth; miR-con, miRNA control; NEAT1, nuclear paraspeckle assembly transcript 1; PTX, paclitaxel; si-con, siRNA control; ZEB1, zinc finger E-box-binding homeobox 1.
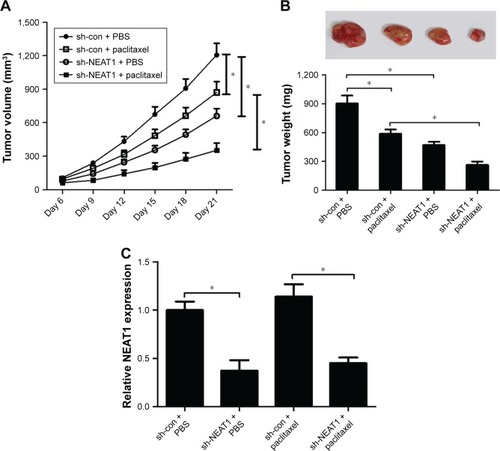
NEAT1 knockdown improved PTX sensitivity of ovarian cancer in vivo
To confirm the functional effect of NEAT1 on PTX resistance in vivo, a xenograft tumor mouse model was established. SKOV3/PTX cells transfected with sh-con or sh-NEAT1 were subcutaneously injected into nude mice. After 6 days, the mice were administrated intraperitoneally with PBS or 3 mg/kg PTX every 3 days. As presented in , PTX treatment or NEAT1 knockdown significantly blocked tumor growth (decreased tumor volume and weight), while combined sh-NEAT1 and PTX led to more obvious inhibition on tumor growth. qRT-PCR analysis showed that NEAT expression was prominently lower in tumor tissues from sh-NEAT1 group than sh-con group (). Thus, it is concluded that NEAT1 knockdown improved PTX sensitivity of ovarian cancer in vivo.
Figure 7 NEAT1 knockdown enhanced PTX sensitivity of ovarian cancer cells in vivo.
Abbreviations: NEAT1, nuclear paraspeckle assembly transcript 1; PTX, paclitaxel; sh, short hairpin; PBS, phosphate buffered solution.
Discussion
Although PTX-based chemotherapy is particularly effective in most cancer cases, the development of chemoresistance is becoming a major impediment to the successful treatment of cancer patients.Citation22 Convincing evidence has indicated the essential roles of lncRNAs in drug resistance of diverse tumors.Citation23 In this study, we focused on the function and mechanism of NEAT1 in PTX resistance in ovarian cancer. The results showed that NEAT1 expression was upregulated in PTX-resistant ovarian cancer tissues and cells. Functional and mechanistic studies manifested that NEAT1 contributed to PTX resistance of ovarian cancer cells possibly through upregulating ZEB1 expression by sponging miR-194.
It is well documented that aberrant expression of NEAT1 plays a crucial role in the development of drug resistance in human cancers. For example, NEAT1 was upregulated in adriamycin-resistant gastric cancer cells and functioned as an oncogene to contribute to chemotherapy resistance of gastric cancer cells.Citation24 NEAT1 expression level was downregulated in leukemia patients and cells and overexpression of NEAT1-mitigated MDR induced by cytotoxic agent through inhibition of ATP-binding cassette G2 (ABCG2) in leukemia.Citation25 Another study reported that NEAT1 enhanced cisplatin sensitivity in non-small-cell lung cancer through functioning as a ceRNA to upregulate EGCG-induced cisplatin transporter CTR1 (copper transporter 1) by sponging miR-98-5p.Citation26 In the present study, it is demonstrated that NEAT1 was upregulated in PTX-resistant ovarian cancer tissues and cells. The efflux pump P-glycoprotein (P-gp), an important member of the ATP-binding cassette (ABC) transporters that facilitate the removal of anticancer drugs from cancer cells, is identified as an important mechanism of resistance to drug treatment.Citation27,Citation28 GST-π, as one of the members of GST family, has been shown to affect drug metabolism and result in drug sequestration in intracellular vesicles.Citation29 Our results demonstrated that NEAT1 knockdown suppressed P-gp and GST-π levels in PTX-resistant ovarian cancer cells, suggesting that NEAT1 knockdown may attenuate drug resistance of ovarian cancer cells. Functional analysis further revealed that NEAT1 knockdown improved PTX sensitivity and promoted PTX-induced apoptosis in PTX-resistant ovarian cancer cells in vitro. Moreover, chemoresistance of NEAT1 was validated on ovarian cancer xenografts in nude mice. These results demonstrated that NEAT1 conferred PTX resistance in PTX-resistant ovarian cancer cells in vitro and in vivo. In agreement with our data, NEAT1 was highly expressed in ovarian cancer tissues and cells and knockdown of NEAT1 elicited suppression on proliferation and invasion in ovarian cancer.Citation16
Recently, a competing endogenous RNAs (ceRNAs) hypothesis proposed that lncRNAs may exert their biological function through acting as molecular sponge for miRNAs, thus in turn leading to derepression of miRNA targets.Citation30 In the present study, miR-194 was demonstrated to be down-regulated in PTX-resistant ovarian cancer tissues and cells. Interestingly, NEAT1 was first demonstrated to interact with miR-194 and negatively regulate its expression. Moreover, enforced expression of miR-194 attenuated PTX resistance in PTX-resistant ovarian cancer cells, consistent with a previous report.Citation18 Furthermore, inhibition of miR-194 reversed NEAT1-knockdown-mediated increase in PTX sensitivity. The inductive effect of miR-194 on drug sensitivity was also reported in non-small-cell lung cancerCitation31 and laryngeal squamous cell carcinoma.Citation32
Additionally, our results demonstrated that ZEB1 was a direct target of miR-194 and NEAT1 derepressed ZEB1 expression by suppressing miR-194. Mechanistic analysis further revealed that miR-194 inhibition or NEAT1 overexpression abolished ZEB1-knockdown-mediated improvement of PTX sensitivity. ZEB1, as a epithelial–mesenchymal transition (EMT) inducing transcription factor, is involved in drug-resistant phenotype.Citation33 A previous document demonstrated that increased expression of ZEB1 contributed to the development of drug resistance in ovarian cancer cells.Citation34 From all these data, it is concluded that NEAT1 acted as a ceRNA to positively regulate ZEB1 expression by sponging miR-194, thus contributing to PTX resistance of ovarian cancer cells. Moreover, it has been reported that TGF-β1 is involved in EMT and cell invasion and NEAT1 knockdown remarkably reduced the expression of TGF-β1 in ovarian cancer cells.Citation35 In the future research, we will focus on whether the regulatory effect of NEAT1/miR-194 on PTX resistance was mediated by TGF-β1 signaling pathway.
Conclusion
This study identified the involvement of NEAT1 in PTX resistance of ovarian cancer cells. NEAT1 enhanced PTX resistance in ovarian cancer cells by working as a ceRNA to sponge miR-194, thus reinforcing the protein level of ZEB1. Therefore, our study contributes to better understanding of the molecular mechanism of drug resistance in ovarian cancer, providing a promising lncRNA-targeted therapy for ovarian cancer.
Disclosure
The authors report no conflicts of interest in this work.
References
- FerlayJShinHRBrayFFormanDMathersCParkinDMEstimates of worldwide burden of cancer in 2008: GLOBOCAN 2008Int J Cancer2010127122893291721351269
- SiegelRLMillerKDJemalACancer statistics, 2015CA Cancer J Clin201565152925559415
- PuvanenthiranSEssapenSSeddonAMModjtahediHImpact of the putative cancer stem cell markers and growth factor receptor expression on the sensitivity of ovarian cancer cells to treatment with various forms of small molecule tyrosine kinase inhibitors and cytotoxic drugsInt J Oncol20164951825183827599579
- DillmanROMcClayEFBarthNMDendritic versus tumor cell presentation of autologous tumor antigens for active specific immunotherapy in metastatic melanoma: impact on long-term survival by extent of disease at the time of treatmentCancer Biother Radiopharm201530518719426083950
- MantiasmaldoneGMEdwardsRPVladAMTargeted treatment of recurrent platinum-resistant ovarian cancer: current and emerging therapiesCancer Manag Res20113253821734812
- WeiWBirrerMJSpleen tyrosine kinase confers paclitaxel resistance in ovarian cancerCancer Cell20152817926175410
- TangYCheungBBAtmadibrataBThe regulatory role of long noncoding RNAs in cancerCancer Lett2017391121928111137
- XiaHHuiKMMechanism of cancer drug resistance and the involvement of noncoding RNAsCurr Med Chem201421263029304124735364
- ZhangJLiuJXuXLiLCurcumin suppresses cisplatin resistance development partly via modulating extracellular vesicle-mediated transfer of MEG3 and miR-214 in ovarian cancerCancer Chemother Pharmacol201779347948728175963
- LiLGuMYouBLong non-coding RNA ROR promotes proliferation, migration and chemoresistance of nasopharyngeal carcinomaCancer Sci201610791215122227311700
- LiZZhaoLWangQOverexpression of long non-coding RNA HOTTIP increases chemoresistance of osteosarcoma cell by activating the Wnt/β-catenin pathwayAm J Transl Res2016852385239327347346
- NiuYFengMHuangWLong non-coding RNA TUG1 is involved in cell growth and chemoresistance of small cell lung cancer by regulating LIMK2b via EZH2Mol Cancer2017161528069000
- SunwooHDingerMEWiluszJEAmaralPPMattickJSSpectorDLMEN epsilon/beta nuclear-retained non-coding RNAs are up-regulated upon muscle differentiation and are essential components of paraspecklesGenome Res200919334735919106332
- YuXLiZZhengHChanMTWuWKNEAT1: a novel cancer-related long non-coding RNACell Prolif2017502
- ChenZJZhangZXieBBZhangHYClinical significance of up-regulated lncRNA NEAT1 in prognosis of ovarian cancerEur Rev Med Pharmacol Sci201620163373337727608895
- ChaiYLiuJZhangZLiuLHuR-regulated lncRNA NEAT1 stability in tumorigenesis and progression of ovarian cancerCancer Med2016571588159827075229
- YoonJHAbdelmohsenKGorospeMFunctional interactions among microRNAs and long noncoding RNAsSemin Cell Dev Biol201434491424965208
- NakamuraKSawadaKKinoseYHashimotoKMabuchiSKimuraTAbstract 4386: identification of microRNA which regulates paclitaxel resistance of ovarian cancer cells – a potential of miR-194 by attenuating paclitaxel resistance through the down-regulation of oncogene BMI-1Cancer Res201474Suppl 19 Abstract nr 4386
- LiZHuSWangJMiR-27a modulates MDR1/P-glycoprotein expression by targeting HIPK2 in human ovarian cancer cellsGynecol Oncol2010119112513020624637
- LuDShiHCWangZXGuXWZengYJMultidrug resistance-associated biomarkers PGP, GST-pi, Topo-II and LRP as prognostic factors in primary ovarian carcinomaBrJ Biomed Sci2011682697421706917
- PilyuginMIrminger-FingerILong non-coding RNA and microRNAs might act in regulating the expression of BARD1 mRNAsInt J Biochem Cell Biol201454835637625008968
- StordalBPavlakisNDaveyRA systematic review of platinum and taxane resistance from bench to clinic: an inverse relationshipCancer Treat Rev200733868870317881133
- ChenQNWeiCCWangZXSunMLong non-coding RNAs in anti-cancer drug resistanceOncotarget2017811925193627713133
- ZhangJZhaoBChenXWangZXuHHuangBSilence of long noncoding RNA NEAT1 inhibits malignant biological behaviors and chemotherapy resistance in gastric cancerPathol Oncol ResIn press2017
- GaoCZhangJWangQRenCOverexpression of lncRNA NEAT1 mitigates multidrug resistance by inhibiting ABCG2 in leukemiaOncol Lett20161221051105727446393
- JiangPWuXWangXHuangWFengQNEAT1 upregulates EGCG-induced CTR1 to enhance cisplatin sensitivity in lung cancer cellsOncotarget2016728433374335127270317
- AndorferPRothenederHRegulation of the MDR1 promoter by E2F1 and EAPPFEBS Lett2013587101504150923542036
- XingAYShiDBLiuWRestoration of chemosensitivity in cancer cells with MDR phenotype by deoxyribozyme, compared with ribozymeExp Mol Pathol201394348148523531420
- ZhangYZhouTDuanJXiaoZLiGXuFInhibition of P-glycoprotein and glutathione S-transferase-pi mediated resistance by fluoxetine in MCF-7/ADM cellsBiomed Pharmacother201367875776223731711
- TayYRinnJPandolfiPPThe multilayered complexity of ceRNA crosstalk and competitionNature2014505748334435224429633
- ZhuXLiDYuFmiR-194 inhibits the proliferation, invasion, migration, and enhances the chemosensitivity of non-small cell lung cancer cells by targeting forkhead box A1 proteinOncotarget2016711131391315226909612
- LiPYangYLiuHMiR-194 functions as a tumor suppressor in laryngeal squamous cell carcinoma by targeting Wee1J Hematol Oncol20171013228122647
- ZhangPSunYMaLZEB1: at the crossroads of epithelial–mesenchymal transition, metastasis and therapy resistanceCell Cycle201514448148725607528
- ZouJLiuLWangQDownregulation of miR-429 contributes to the development of drug resistance in epithelial ovarian cancer by targeting ZEB1Am J Transl Res2017931357136828386361
- LiPZhangXLinLChenGChenJSilencing of the long non-coding RNA NEAT1 suppresses ovarian cancer cell proliferation, migration and invasionInt J Clin Exp Pathol20169551385147