
Abstract
Introduction
Wilms’ tumor (WT), the most common childhood tumor, occurs in sporadic or familial forms. Recent findings reported that abnormal expression in microRNA (miRNA) suggests an important role of miRNAs during WT progress. MiRNAs are endogenous short-chain noncoding RNAs, which have been reported as key biomarkers for detecting tumor onset and progression. However, the functional role of miR-1180 in WT has remained unknown.
Materials and methods
MTT and clonogenic survival assays were used to detect WT cell proliferation. Flow cytometry Annexin V-FITC was used to measure apoptosis. In addition, proteins expressions in the cells were determined by Western blotting.
Results
In the present study, we demonstrated that miR-1180 is upregulated in WT when compared with adjacent tissues by quantitative reverse-transcription polymerase chain reaction. In addition, the inhibition of miR-1180 induced apoptosis in SK-NEP-1 cell line in vitro. Moreover, luciferase reporter assay showed that p73 protein was the target of miR-1180, which was confirmed by the results of Western blotting. Finally, in vivo data indicated that the tumor growth in mice was significantly inhibited by miR-1180 inhibitor.
Conclusion
Our results indicate that miR-1180 might serve as a therapeutic target for future WT therapy.
Introduction
Wilms’ tumor (WT) is one of the most common pediatric renal tumors, and it affects 1 in 10,000 children of Western descent,Citation1 with a peak incidence at 3 years of age. Remarkable progress has been made in the treatment of children with WT. The overall survival rates exceeded 90% in 2009,Citation2 compared with about 30% in the 1930s.Citation3 The large proportion of affected children are cured with modern multidisciplinary therapy and with information from multi-institutional clinical trials.Citation4 However, about 25% of the patients cannot be cured by current treatments.Citation5,Citation6 It is reported that chromosomal abnormalities stimulate the uncontrolled growth of these undifferentiated cells and promote the formation of WT.Citation7,Citation8 Although there have been advances in understanding the pathogenesis of WT, the molecular mechanisms responsible for this differentiation failure are not completely understood.
MicroRNAs (miRNAs) are endogenous short-chain noncoding RNAs (20–24 nucleotides) that regulate gene function by direct binding of target mRNAs and then participating in multiple biological aspects of cancer progression, such as cell proliferation, cell invasion, cell migration, and cell angiogenesis.Citation9,Citation10 MiRNAs have emerged as novel biomarkers for diagnosis and potential therapeutic targets for therapy of human cancers,Citation11,Citation12 including WT.Citation13–Citation17 MiRNAs control gene expression either by degradation of target mRNAs or posttranscriptional repression through targeting the 3′-untranslated region (3′-UTR) in a sequence-specific manner.Citation10,Citation18 It is reported that miR-483-5p could predict an adverse prognosis in WT.Citation19 MiR-204Citation20 and miR-185Citation21 were also found to be significantly decreased in WT. These reports demonstrate that miRNAs play important roles in the cancer progression of WT. MiR-1180 was confirmed to be upregulated in hepatocellular carcinoma (HCC) clinical tissues and cell lines.Citation22 It is reported that miR-1180 promoted apoptotic resistance to human HCC via activation of NF-κB signaling pathway.Citation23 However, the functional role of miR-1180 in WT remains unknown.
Recently, p73 gene has been identified as a candidate gene for tumor suppression, and it plays an important role in the development of many tumors.Citation24,Citation25 The transcription factor p73 is homologous to the well-known tumor suppressor p53, which is the most frequently mutated gene in cancer. The p53-regulated networks are often impaired in cancer.Citation26 p73 has attracted increasing attention, because it may substitute for impaired p53-regulated networks which are commonly perturbed in cancer.Citation27 p73 mediates cell migration and restores anti-oncogenic responses, as well as epithelial cell mesenchyme transformation (EMT)-related mRNA co-targets (vimentin/β-catenin/lef1).Citation28 It was recently shown that the p73-dependent target miR-3158 was found to induce anti-oncogenic/anti-invasive responses in p53-mutant cancer cells.Citation28 However, the relationship of p73 and miR-1180 remain unknown.
In the present study, we demonstrate the upregulation of miR-1180 in WT. We find that the ectopic expression of miR-1180 has an antiapoptotic effect in HCC, while its inhibition increases cell apoptosis. Moreover, we found that miR-1180 might regulate WT cell proliferation by targeting p73 protein. Our results present novel insights into understanding the molecular mechanism of WT tumorigenesis and provide a future direction for interventional therapeutic targets for WT.
Materials and methods
Ethics statement
This study was approved by the Ethics Committee of Zhoukou Central Hospital. The specimen collection and the whole procedure were harmless to the patients, and written informed consent was obtained from the guardians of the child patients. The entire investigation conforms to the principles outlined in the Declaration of Helsinki.
Primary tumors
From March 2014 to December 2016, WT tissues (30 cases) and paired homolateral adjacent non-tumorous renal tissues (≥3 cm away from the tumor, with pathological confirmation that no cancer cells were found in tangent lines) were obtained from 30 patients who underwent tumor nephrectomy at the Zhoukou Central Hospital. Every nephrectomy specimen was immediately snap-frozen in liquid nitrogen and stored at −80°C until RNA and protein extraction was performed.
RNA extraction and quantitative real time polymerase chain reaction (qRT-PCR)
Total RNA from the tissue samples was extracted with Trizol reagent (Thermo Fisher Scientific, Waltham, MA, USA). cDNA was synthesized by reverse transcription using TaqMan® MicroRNA Reverse Transcription Kit (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. Products were amplified by PCR using a TaqMan Universal PCR Master Mix Kit (Thermo Fisher Scientific, Waltham, MA, USA). The relative levels of miR-1180 in WT and in adjacent non-tumorous tissues were calculated with the 2−ΔΔCt method. The primers were purchased from RiboBio Co., Ltd, Guangzhou, China. MiR-1180 forward: 5′-AACTACCTGGACCGCTTCCT-3′; reverse: 5′-CCACTTGAGCTTGTTCACCA-3′. GAPDH forward: 5′-TATGATGATATCAAGAGGGTAGT-3′; reverse: 5′-TGTATCCAAACTCATTGTCATAC-3′.
Cell lines and transfection
WT cell line SK-NEP-1, purchased from American Type Culture Collection (ATCC, Rockville, MD, USA), was routinely cultured in RPMI-1640 containing 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Waltham, MA, USA) with 100 units/mL penicillin and 100 μg/mL streptomycin (Sigma, St-Louis, MO, USA) in a humidified incubator at 37°C containing 5% CO2. All cell lines used were between passages 3 and 8 for each experiment. Cells were plated to about 60% confluency in medium without antibiotics, the day before transfection. MiR-1180 inhibitor and scramble, which were labeled with green fluorescence protein (GFP), were purchased from Genepharma (Suzhou, China). Cell transfection was performed using Lipofectamine RNAiMax (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. The medium was replaced 8 hours after transfection and then changed daily.
MTT assay
A colorimetric assay with tetrazolium salt was used to assess the antiproliferative effects. Twenty-four hours after transfection, cells were seeded in 96-well plates (2,000 cells/well) and allowed to grow for 48 hours. Then 10 μL of MTT solution (5 mg/mL) in PBS was added to each well and incubated at 37°C for 4 hours. The formazan crystals were dissolved with dimethyl sulfoxide (DMSO), and the absorbance of the wells at 570 nm was measured with a microplate reader. All experiments were carried out more than three times, and the data are presented as means ± standard deviation. Statistical analysis of the data was performed using Excel program.
Clonogenic survival assay
After transfection for 24 hours, cells were trypsinized, counted, and reseeded in six-well plates at the density of 1,000 cells/well for 8 days with complete medium. The medium was changed every 3 days to maintain the cell growth. The colonies were fixed and stained with 0.5% crystal violet (Sigma, St Louis, MO, USA) for 30 minutes at room temperature. After 8 days of culturing, the number of colonies was counted under an inverted microscope.
Flow cytometry Annexin V-FITC Binding Assay
Flow cytometry Annexin V-FITC Binding Assay (R&D, Minneapolis, MN, USA, Cat No TA4638) was used to assay apoptosis and necrosis in WT cells after transfection. The cells were separated with 0.25% trypsin and 0.02% ethylenediaminetetraacetic acid and centrifuged at 1,000 rpm for 5 minutes. Then, the cell pellet was washed and resuspended in binding buffer in a microfuge tube. Five microliters of fluorescein isothiocyanate (FITC)-conjugated Annexin V and 10 μL of propidium iodide (PI) were added to the tubes, mixed and incubated in the dark for 10 minutes. The labeled cells were tested using the FACS Calibur system (BD Biosciences, Franklin Lake, NJ, USA), and the percentages of early apoptotic, late apoptotic, and necrotic cells were determined. Results are representative of three independent experiments with triplicate samples.
Western blot analysis
About 5×106 cells were gathered after transfection for 24 hours and lysed in RIPA buffer (Cell Signaling Technology, Danvers, MA, USA) in the presence of protease inhibitor (phenylmethylsulfonyl fluoride) and phosphatase inhibitor (Na-orthovanadate and NaF). Nuclear proteins were prepared using a commercial kit (Thermo Fisher Scientific). Western blotting was performed as previously described.Citation29 In brief, equal amounts of total protein extract from cultured cells and tissues were fractionated by 8%–10% SDS-PAGE and electrically transferred onto polyvinylidene difluoride (PVDF) membranes. Mouse or rabbit primary antibodies and appropriate horseradish peroxidase-conjugated secondary antibodies were used to detect the designated proteins. The bound secondary antibodies on the PVDF membrane were reacted with enhanced chemiluminescent (ECL) detection reagents (Thermo Scientific) and exposed to X-ray films. Results were normalized to the internal control β-actin.
Luciferase reporter assay
The pGL3-luciferase reporter gene plasmid pGL3-P70 3′ UTR was transfected into the cells with miR-1180 using Lipofectamine 2000 Reagent (Thermo Fisher Scientific). After 48 hours of transfection, the cells were collected, and the luciferase activities were measured using the Dual-Luciferase Reporter Assay System (Promega, Madison, WI, USA).
Subcutaneous tumor model
BALB/c-nu mice (5–6 weeks old, 18–20 g) were purchased from Vital River (Beijing, China) and housed in barrier facilities in a 12 hour light/dark cycle. The mice were randomly assigned to three groups (six mice/group). All mice in groups were inoculated subcutaneously with SK-NEP-1 cells (5×106 cells per mouse) transfected with miR-1180 inhibitor or control vector in the left dorsal flank. Tumor volumes were examined weekly – length, width, and thickness were measured with calipers, and tumor volumes were calculated. Tumor volume was calculated using the equation (L×W2)/2. Four weeks later, the animals were euthanized, and the tumors were excised and weighed. This study was performed in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Committee on the Ethics of Animal Experiments of the Zhoukou Central Hospital University.
Statistical analysis
Statistical analysis was conducted using GraphPad Prism 7 (GraphPad Software, Inc., La Jolla, CA, USA). All the data are expressed as mean values ± SD. Comparisons among two groups were made with Student’s t-test. Comparisons among multiple groups were made with one-way analysis of variance (ANOVA) followed by Dunnett’s test. p<0.05 was used for statistical significance. All experiments were performed in triplicates.
Results
Association between miR-1180 expression and pathological variables
summarizes the correlation of miR-1180 and age, gender, histopathological type, NWTS (National Wilms’ Tumor Studies) stage, and lymphatic metastasis. The median expression level of miR-1180 in all 30 patients with WT was 2.847. We divided the patients into two groups according to their miR-1180 expression levels using the median score as a cutoff: the high miR-1180 expression group (2−ΔΔCt ≥2.847, n=13) and the low miR-21 expression group (2−ΔΔCt <2.847, n=17). ANOVA followed by Dunnett’s test showed that no significant correlation existed between age (p=0.742), gender (p=0.628), and the expression of miR-1180. However, there was a clear significant correlation between the expression of miR-1180 and histopathological type (p=0.045) as well as NWTS stage (p=0.034) and lymphatic metastasis (p=0.026). Together, these results indicated that WT patients with more aggressive tumors exhibited significantly higher miR-1180 expression compared with that of patients with less aggressive tumors.
Table 1 Correlation of miR-1180 expression levels with clinicopathological parameters of Wilms’ tumor patients
MiR-1180 expression in WT tissues was significantly higher than that in adjacent non-tumorous tissues
The relative expression levels of miR-1180 in the WT and in the non-tumorous tissues were 3.50 and 2.19, respectively (). Compared with the non-tumorous tissues, the expression level of miR-1180 in WT tissues was significantly elevated.
Figure 1 MiR-1180 is upregulated in WT, and the inhibition of miR-1180 suppressed WT cell proliferation.
Abbreviations: DAPI, 4′,6-diamidino-2-phenylindole; DMSO, dimethyl sulfoxide; GFP, green fluorescence protein; NC, normal control; qRT-PCR, quantitative reverse-transcription polymerase chain reaction; WT, Wilms’ tumor.
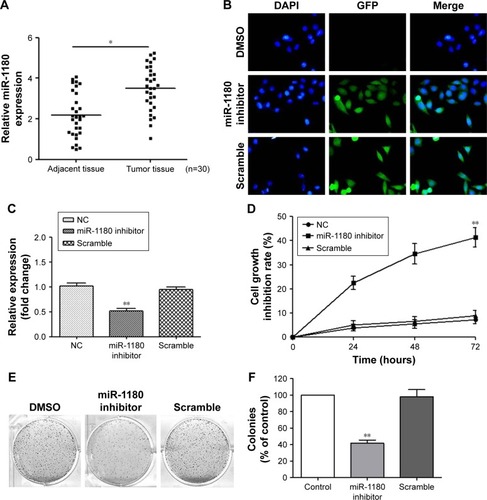
The proliferation of SK-NEP-1 cells was significantly inhibited after transfection with miR-1180 inhibitor
To analyze the effect of miR-1180 on the proliferation of SK-NEP-1 WT cells, SK-NEP-1 WT cells were transfected with miR-1180 inhibitor. The luciferase assay and qRT-PCR were used to verify the transfection efficiency (). To further explore the role of miR-1180 in WT progression, we first monitored the impact of miR-1180 inhibitor on cell viability using the MTT assay. MiR-1180 inhibitor significantly increased cell growth inhibition rate compared with the scramble (SK-NEP-1 WT cells + transfection reagent + miR-1180 inhibitor negative control reagent) and normal control (NC) group (SK-NEP-1 WT cells + transfection reagent + PBS) (p<0.01; ). Furthermore, the colony formation assays showed that miR-1180 inhibitor led to less and smaller-sized colonies compared with control and scramble cells ().
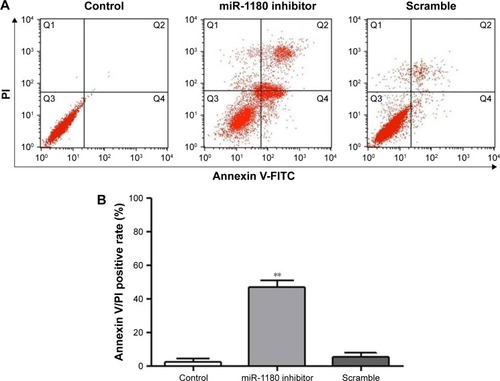
The apoptosis of SK-NEP-1 WT cells was promoted after transfection with miR-1180 inhibitor
The results of flow cytometry showed that after being transfected with miR-1180 inhibitor for 48 hours, the Annexin-V-FITC-positive rate (%) of the miR-1180 inhibitor group (47.01±3.74) was significantly higher than the control (2.38±1.77) and scramble groups (5.39±2.42) (p<0.01, ). No significant difference in the apoptotic rate was detected between the control and scramble groups (p>0.05).
Figure 2 The percentages of apoptotic cells in WT were determined in flow cytometry experiments using Annexin V/PI double staining.
Abbreviations: FITC, fluorescein isothiocyanate; PI, propidium iodide; WT, Wilms’ tumor.
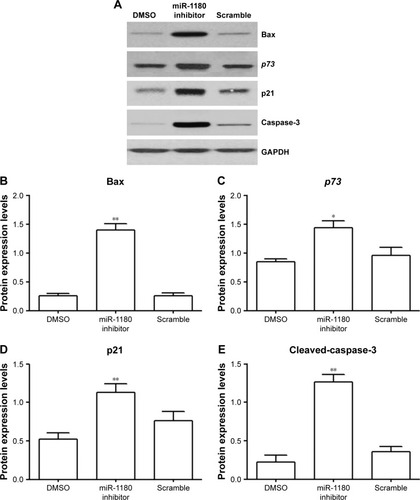
MiRNA-1180 regulated WT apoptosis via a p73-dependent mechanism
In order to investigate the effect of miR-1180 inhibitor on the apoptosis of WT, the expression levels of related proteins were examined. MiR-1180 led to an increase in Bax, p73, p21, and cleaved-caspase-3 expressions (). p73, a member of p53 tumor suppressor family, plays a crucial role in tumor suppression.Citation30 The increased expression of p73, as well as its downstream factors, p21 and the pro-apoptotic Bax, indicated that miRNA-1180 might have an antiapoptotic role in cleaved-caspase-3 expression.
Figure 3 MiRNA-1180 regulates WT apoptosis via a p73-dependent mechanism.
Abbreviations: DMSO, dimethyl sulfoxide; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; WT, Wilms’ tumor.
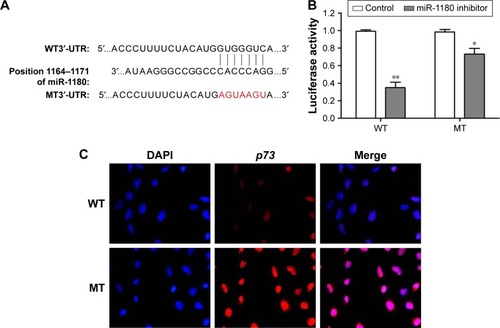
MiRNA 1180 regulated WT apoptosis by targeting p73
Comparison of the miRNA-1180 and p73 3′-UTR sequences indicated that p53 may be a direct target of miRNA-1180 (). Luciferase reporter assays were used to determine the molecular mechanism underlying the effects of miRNA-1180 on WT. As shown in , the luciferase activity of the 3′-UTR of p73 was downregulated in the presence of miR-1180 (p<0.01; ). These results indicated that miR-1180 directly targeted p73 in WT cells.
Figure 4 Validation of the direct interaction between miRNA-1180 and p73.
Abbreviations: 3′-UTR, 3′-untranslated region; DAPI, 4′,6-diamidino-2-phenylindole; MT, mutant; WT, Wilms’ tumor.
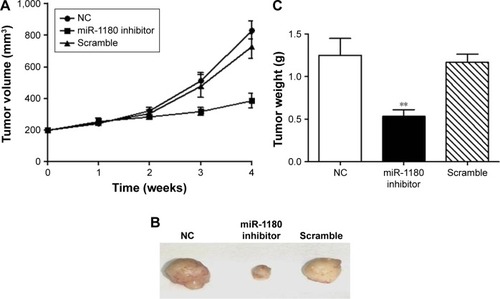
Inhibition of miR-1180 suppressed WT xenograft tumor growth in nude mice
To evaluate the antitumor effect of miR-1180 in vivo, we screened SK-NEP-1-pooled clones with G418 to gain the stable expression of miR-1180 inhibitor (SK-NEP-1/miR-1180 inhibitor) and established a WT xenograft model by inoculating indicated cells in the left dorsal flanks of nude mice. After 4 weeks, compared with the control group, the volume of nodules was smaller in the SK-NEP-1/miR-1180 inhibitor group than in the scramble group (). The tumor weight of the SK-NEP-1/miR-1180 inhibitor group was 0.53 g, which was also much smaller than the control or scramble group (). The in vivo result was consistent with in vitro data, which confirmed the antiapoptosis role of miR-1180 in WT.
Figure 5 Inhibition of miR-1180 suppresses SK-NEP-1 tumor growth in vivo.
Abbreviation: NC, normal control.
Discussion
In the present study, we showed that miR-1180 was upregulated in both clinical tissues. MTT and clonogenic survival assay performed by transfecting miR-1180 inhibitor into SK-NEP-1 cells illustrated that miR-1180 was capable of inducing the cell growth of WT. Furthermore, miR-1180 inhibitor regulated the apoptosis of the WT. We further examined this process and found that p73 and the targets played important roles in the regulation. The in vivo experiment indicated that miR-1180 inhibitor inhibited WT growth. These results revealed that miR-1180 might induce apoptosis of WT by targeting p73.
Previous studies have indicated that miR-1180 was implicated in diverse biological processes.Citation23 MiR-1180 was reported to be upregulated in HCC clinical tissues and cell lines.Citation22 MiR-1180 significantly regulated TNIP2 expression and then regulated the expression of NF-κB downstream target genes, Cyclin D1, Myc, and p-Rb.Citation22 Suppressing miR-1180 expression exerted anticancer effects on pancreatic cancer cells via regulation of the TNIP2/NF-κB signaling pathway.Citation31 In accordance with these reports, our results showed that miR-1180 was upregulated and acted as a tumor promoter in the development of WT. The current study also showed that miRNA-1180 regulated WT apoptosis via targeting p73.
The tumor suppressor p73 is known to function as a transcription factor. The identification and characterization of the p53/p63/p73 network provides evidence of a tight link between developmental processes and tumorigenesis.Citation32 p73 can induce cell growth arrest and apoptosis in response to stress signals, two functional properties that it shares with p53. It is reported that the tumor suppressors p53, p63, and p73 function as both positive and negative regulators of the miRNA processing components.Citation33 p53, p63 and p73 appear to regulate the processing of miRNAs, such as let-7, miR-200c, miR-143, miR-107, miR-16, miR-145, miR-134, miR-449a, miR-503, and miR-21.Citation33 On the other hand, p73 was regulated by multiple mechanisms, including transcription and mRNA and protein stability.Citation34 MiRNAs such as miRNA-193a-5p is implicated in p73 regulation. Inhibiting the miR-193a-5p increases the expression of Tap73β.Citation35 Together, it is conjectured that miRNAs and p73 play important roles and regulate each other in tumor progression, but this requires more genetic and biochemical experiments.
In conclusion, the current study showed that miR-1180 was markedly upregulated in WT patients. Moreover, the inhibition of miR-1180 upregulated the expression of p73 and induced apoptosis in WT cell line. These findings collectively implicated that miR-1180 might serve as a therapeutic target for future WT therapy.
Acknowledgments
This work was supported by Ke Wu, who proofread the manuscript and offered kind suggestions.
Disclosure
The authors report no conflicts of interest in this work.
References
- GrovasAFremgenARauckAThe National Cancer Data Base report on patterns of childhood cancers in the United StatesCancer19978012232123329404710
- DavidoffAMWilms’ tumorCurr Opin Pediatr200921335736419417665
- FarberSChemotherapy in the treatment of leukemia and Wilms’ tumorJAMA196619888268364288581
- DomeJSGrafNJamesIAdvances in Wilms Tumor Treatment and Biology: progress through international collaborationJ Clin Oncol201533272999300726304882
- MalogolowkinMCottonCAGreenDMTreatment of Wilms tumor relapsing after initial treatment with vincristine, actinomycin D, and doxorubicin. A report from the National Wilms Tumor Study GroupPediatr Blood Cancer200850223624117539021
- SpreaficoFPritchard JonesKMalogolowkinMHTreatment of relapsed Wilms tumors: lessons learnedExpert Rev Anticancer Ther20099121807181519954292
- HohensteinPPritchard-JonesKCharltonJThe yin and yang of kidney development and Wilms’ tumorsGenes Dev201529546748225737276
- SpreaficoFCiceriSGambaBChromosomal anomalies at 1q, 3, 16q, and mutations of SIX1 and DROSHA genes underlie Wilms tumor recurrencesOncotarget2016788908891526802027
- AmbrosVThe functions of animal microRNAsNature2004431700635035515372042
- BartelDPMicroRNAs: genomics, biogenesis, mechanism, and functionCell2004116228129714744438
- HernandoEmicroRNAs and cancer: role in tumorigenesis, patient classification and therapyClin Transl Oncol20079315516017403626
- OsadaHTakahashiTMicroRNAs in biological processes and carcinogenesisCarcinogenesis200728121217028302
- HeJGuoXSunLWangKYaoHNetworks analysis of genes and microRNAs in human Wilms’ tumorsOncol Lett20161253579358527900039
- YuXLiZChanMTWuWKThe roles of microRNAs in Wilms’ tumorsTumor Biol201637214451450
- YuXLiZChanMTWuWKThe roles of microRNAs in Wilms’ tumorsTumour Biol20163721445145026634744
- WegertJIshaqueNVardapourRMutations in the SIX1/2 pathway and the DROSHA/DGCR8 miRNA microprocessor complex underlie high-risk blastemal type Wilms tumorsCancer Cell201527229831125670083
- LudwigNNourkami-TutdibiNBackesCCirculating serum miRNAs as potential biomarkers for nephroblastomaPed Blood Cancer201562813601367
- FabbriMCaloreFPaoneAGalliRCalinGAEpigenetic regulation of miRNAs in cancerAdv Exp Med Biol201375413714822956499
- WatsonJABryanKWilliamsRmiRNA profiles as a predictor of chemoresponsiveness in Wilms’ tumor blastemaPLoS One201381e5341723308219
- KollerKPichlerMKochKNephroblastomas show low expression of microR-204 and high expression of its target, the oncogenic transcription factor MEIS1Pediatr Dev Pathol201417316917524617557
- ImamJSBuddavarapuKLee-ChangJSMicroRNA-185 suppresses tumor growth and progression by targeting the Six1 oncogene in human cancersOncogene201029354971497920603620
- ZhouXZhuHQMaCQMiR-1180 promoted the proliferation of hepatocellular carcinoma cells by repressing TNIP2 expressionBiomed Pharmacother20167931532027044843
- TanGWuLTanJMiR-1180 promotes apoptotic resistance to human hepatocellular carcinoma via activation of NF-kappaB signaling pathwaySci Rep201662232826928365
- GuoHYangSXuLAssociation between the p73 gene G4C14-to-A4T14 single nucleotide polymorphism and risk of cervical cancer by high resolution melting and PCR with confronting two-pair primers in a Chinese populationOncol Lett201612172172627347206
- NgKYChanLHChaiSTP53INP1 down-regulation activates a p73-dependent DUSP10/ERK signaling pathway to promote metastasis of hepatocellular carcinomaCancer Res201777174602461228674078
- VogelsteinBLaneDLevineAJSurfing the p53 networkNature2000408681030731011099028
- ChakrabortyJBanerjeeSRayPGain of cellular adaptation due to prolonged p53 impairment leads to functional switchover from p53 to p73 during DNA damage in acute myeloid leukemia cellsJ Biol Chem201028543331043311220675383
- GaltsidisSLogothetiSPavlopoulouAUnravelling a p73-regulated network: The role of a novel p73-dependent target, MIR3158, in cancer cell migration and invasivenessCancer Lett20173889610627919789
- ZhangHYangJYZhouFSeed Oil of Brucea javanica induces apoptotic death of acute myeloid leukemia cells via both the death receptors and the mitochondrial-related pathwaysEvid Based Complement Alternat Med2011201196501621760826
- ZhangYYoungAZhangJChenXP73 tumor suppressor and its targets, p21 and PUMA, are required for Madin-Darby canine kidney cell morphogenesis by maintaining an appropriate level of epithelial to mesenchymal transitionOncotarget2015616139941400426101856
- GuLZhangJShiMPengCThe effects of miRNA-1180 on suppression of pancreatic cancerAm J Transl Res2017962798280628670370
- ZaikaAIEl-RifaiWThe role of p53 protein family in gastrointestinal malignanciesCell Death Differ200613693594016557273
- BoominathanLThe tumor suppressors p53, p63, and p73 are regulators of microRNA processing complexPLoS One201055e1061520485546
- ZhangMZhangJYanWChenXp73 expression is regulated by ribosomal protein RPL26 through mRNA translation and protein stabilityOncotarget2016748782557826827825141
- JacquesCCallejaLRBaud’huinMmiRNA-193a-5p repression of p73 controls Cisplatin chemoresistance in primary bone tumorsOncotarget2016734545035451427486986