
Abstract
FH535 is a small-molecule inhibitor of the Wnt/β-catenin signaling pathway, which a substantial body of evidence has proven is activated in various cancers, including pancreatic cancer. Activation of the Wnt/β-catenin pathway plays an important role in tumor progression and metastasis. We investigated the inhibitory effect of FH535 on the metastasis and growth of pancreatic cancer cells. Western blotting and luciferase reporter gene assay indicated that FH535 markedly inhibited Wnt/β-catenin pathway viability in pancreatic cancer cells. In vitro wound healing, invasion, and adhesion assays revealed that FH535 significantly inhibited pancreatic cancer cell metastasis. We also observed the inhibitory effect of FH535 on pancreatic cancer cell growth via the tetrazolium and plate clone formation assays. Microarray analyses suggested that changes in the expression of multiple genes could be involved in the anti-cancer effect of FH535 on pancreatic cancer cells. Our results indicate for the first time that FH535 inhibits pancreatic cancer cell metastasis and growth, providing new insight into therapy of pancreatic cancer.
Introduction
Pancreatic cancer is one of the most aggressive human malignancies worldwide. Despite improvements in surgical and chemotherapeutic approaches over the past decades, the prognosis of pancreatic cancer remains dismal; the average overall 5-year survival rate is <5%.Citation1 The reasons for this are the challenges associated with diagnosis, which tends to be late and uncertain; more importantly, therapeutic options are limited. Even with early diagnosis and surgical resection with curative intention, nearly all patients develop local recurrence or distant metastases following surgery and eventually succumb to the debilitating effects of metastatic growth.Citation2,Citation3 Conventional chemotherapy is rarely curative for metastatic pancreatic cancer. In recent years, there have been important advances in the organization of care for patients with pancreatic cancer; these advances have also resulted in more focused studies on surgical, oncological, and immunological treatment.
The Wnt/β-catenin pathway is a genetically conserved signaling pathway associated with a variety of human conditions such as birth defects and tumors. Abnormal Wnt/β-catenin pathway activation is closely related to the development of many cancers.Citation4,Citation5 An increasing amount of evidence demonstrates that both the β-catenin-dependent (canonical) and β-catenin-independent (non-canonical) Wnt signaling pathways play a key role in regulating pathological processes by facilitating tumor growth, migration, and invasion. In canonical Wnt signaling, glycogen synthase kinase-3β (GSK-3β) phosphorylates β-catenin at certain key residues, leading to its ubiquitination and subsequent degradation.Citation5,Citation6 Non-phosphorylated β-catenin accumulates in the cytoplasm, and pathway activation leads to nuclear accumulation of β-catenin and interaction with T-cell factor (TCF) transcription factors, subsequently stimulating the downstream target genes, which include the genes participating in cell metastasis and proliferation.Citation7,Citation8
Abnormal Wnt/β-catenin pathway activation plays an important role in human pancreatic cancer, where it causes extracellular matrix degradation and uncontrolled cell proliferation and differentiation.Citation9 Recent studies have demonstrated that FH535 is a synthetic inhibitor of the canonical Wnt signaling pathway; it inhibits the growth of colon, lung, breast, and hepatocellular carcinoma lines,Citation10,Citation11 suggesting that small-molecule targeting of the Wnt/β-catenin pathway could be a promising therapeutic approach for cancers in which this pathway is activated.
In this study, we investigated the anti-cancer effect of FH535 on pancreatic cancer and explored the mechanisms underlying the effect, providing a rationale for further development of FH535 as a promising therapeutic agent for treating pancreatic cancer.
Materials and methods
Cell cultures and reagents
The human pancreatic cancer cell lines PANC-1 and BxPC-3 were purchased from American Type Culture Collection (ATCC) (Manassas, VA, USA). The cells were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM; Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal calf serum (FCS), 100 U/mL penicillin, and 100 μg/mL streptomycin (Thermo Fisher Scientific) at 37°C in a 5% CO2 incubator under a humidified atmosphere; the cells were passaged every 2–3 days for exponential growth. FH535 was purchased from EMD Millipore (Billerica, MA, USA).
Western blotting
Total protein was extracted using a lysis buffer (50 mM Tris-HCl [pH 7.4], 150 mM NaCl, 1% Triton X-100, 0.1% sodium dodecyl sulfate [SDS], 1 mM EDTA) supplemented with a protease inhibitor cocktail kit and a phosphatase inhibitor cocktail kit (Hoffman-La Roche Ltd., Basel, Switzerland). The protein extracts were loaded, size-fractionated by SDS-polyacrylamide gel electrophoresis, and transferred to polyvinylidene difluoride membranes (Bio-Rad Laboratories Inc., Hercules, CA, USA). After blocking, the membranes were incubated with the primary antibodies mouse anti-β-catenin (Santa Cruz Biotechnology Inc., Dallas, TX, USA) and rabbit anti-β-actin (Proteintech Group Inc., Chicago, IL, USA) at 4°C overnight. Protein expression was determined using horseradish peroxidase-conjugated anti-mouse or anti-rabbit secondary antibodies, followed by detection using enhanced chemiluminescence (EMD Millipore). Band intensity was visualized using a JS-1035 image analysis scanning system (Shanghai Peiqing Science & Technology, Co., Ltd., Shanghai, People’s Republic of China).
Luciferase reporter assay
β-catenin is a dominant factor in the Wnt/β-catenin/TCF signaling pathway, which regulates gene transcription by binding β-catenin and TCF. The activity of this final step in the pathway can be precisely measured using a luciferase reporter construct. The reporter plasmid pTOPFLASH (TCF reporter plasmid; EMD Millipore) contains two sets (the second set is in the reverse orientation) of three copies of the TCF binding site (wild-type) upstream of the thymidine kinase minimal promoter and luciferase open reading frame. The internal control plasmid pRL-SV40 (Promega Corporation, Fitchburg, WI, USA) contains the Renilla luciferase gene. Cells were transiently cotransfected with pTOPFLASH plasmid (500 ng/well) and pRL-SV40 plasmid (100 ng/well) for 6 hours using Lipofectamine 2000 (Thermo Fisher Scientific) according to the manufacturer’s protocol. Then, the medium was renewed and FH535 was added. After 24 hours of treatment, cell lysates were subjected to the dual luciferase reporter assay according to the manufacturer’s recommendations; luciferase activity was measured using a luminometer (Turner Designs, Sunnyvale, CA, USA). The results are expressed as relative luciferase activity, ie, the ratio of firefly luciferase activity over Renilla luciferase activity.
Wound healing assay
Cells (1×104/well) were seeded in 96-well plates and grown to confluence. The monolayer culture was artificially scrape wounded with a sterile micropipette tip to create a denuded zone of constant width. Each well was washed with phosphate-buffered saline twice to remove the detached cells before FH535 treatment. Cell migration to the wounded region was observed using an XDS-1B inverted microscope (MIC Optical and Electrical Instrument, Chongqing, People’s Republic of China) and photographed (×40 magnification). Images were captured at 0, 8, and 12 hours to monitor the wound healing process. The wound areas were measured using ImageJ (NIH, Bethesda, MA, USA).
Transwell invasion assay
We used a 24-well Transwell plate with an 8 μm pore size polycarbonate filter membrane (Corning Incorporated, Corning, NY, USA). Cells (1×105) in 100 μL serum-free DMEM were added to the Matrigel-coated top chamber (BD Biosciences, San Jose, CA, USA); the bottom chamber contained DMEM with 10% FCS. The cells were incubated for 24 hours; cells that had invaded through the Matrigel-coated membrane were fixed and stained with crystal violet and counted under a light microscope in five random fields in a blinded fashion.
Adhesion assay
Cells were resuspended in complete medium and seeded in 24-well plates at 1×104 cells/mL. After 5-hour incubation, the unattached cells were removed to another well. The attached and unattached cells were evaluated using the 3-[4,5-dimethylthiazol-2-yl] 2,5-diphenyltetrazolium bromide (MTT) assay. The adhesion rate was calculated as follows: (absorbance of attached cells/[absorbance of attached cells + absorbance of unattached cells]) ×100%.
MTT assay
Cell growth was evaluated using the MTT assay. Cells (5×104/well) were seeded in 24-well tissue culture plates. Blank control was treated with DMSO. After FH535 treatment, MTT (Sigma-Aldrich Co., St Louis, MO, USA) was added to each well (final concentration, 0.5 mg/mL), followed by 4-hour incubation at 37°C. The medium was removed, and 800 μL of dimethyl sulfoxide was added to each well. The absorbance of the mixture was measured at 490 nm using a microplate enzyme-linked immunosorbent assay reader (Bio-Rad Laboratories Inc.). The relative cell viability was calculated as follows: relative cell viability = (mean experimental absorbance/mean control absorbance) ×100%.
Plate clone formation assay
Cells (200/well) were seeded in 24-well plates and treated after 12 hours. After 15 days, the cells were stained with 1% methylrosanilinium chloride, and the number of visible colonies was counted. The relative clone formation ability was calculated as follows: (mean experimental clone number/mean control clone number) ×100%.
Cell cycle analysis
Before treatment, the cells were serum starved for 24 hours to synchronize the cell cycle. Then, FCS was added to the cells, followed by various concentrations of FH535. Following 24 hours of FH535 treatment, the cells were fixed in 80% cooled ethanol and incubated with 0.5% Triton X-100 solution containing 1 mg/mL RNase A at 37°C for 30 minutes. Next, propidium iodide (Sigma-Aldrich Co.) was added to the wells (final concentration, 50 μg/mL), followed by 30-minute incubation in the dark. Cellular DNA content was analyzed using a fluorescence-activated cell sorter (Becton Dickinson, Franklin Lakes, NJ, USA). Data were processed using ModFit LT software (Verity Software House, Topsham, ME, USA).
Microarray assay
Sample preparation and processing were performed as described in the GeneChip Expression Analysis Manual (Agilent Technologies, Santa Clara, CA, USA). Differentially expressed genes were screened using Agilent 44K human whole-genome oligonucleotide microarrays. The selection criterion was greater than twofold difference in expression (difference in upregulated expression was greater than twofold; difference in downregulated expression was less than 0.5-fold). Hierarchical clustering of samples was performed using an average linkage algorithm using TIGR MultiExperiment Viewer (The Institute for Genomic Research, Rockville, MD, USA).
Statistical analysis
Each experiment was performed in at least triplicate. Results are expressed as the mean ± standard deviation. Statistical analysis was performed using an unpaired Student’s t-test. P<0.05 was considered significant.
Results
FH535 inhibited the β-catenin pathway in pancreatic cancer cells
Treatment with 20 μM FH535Citation12 did not affect nuclear or total β-catenin expression in the BxPC-3 cells, but downregulated nuclear and total β-catenin in the PANC-1 cells (). The luciferase reporter assay confirmed that FH535 suppressed TCF-dependent transcription, which may have led to dysregulation of the genes downstream of the β-catenin pathway (). To verify this, we performed microarray analyses to determine the mRNA expression changes in 138 genes downstream of the β-catenin pathway using Agilent 44K human whole-genome oligonucleotide microarrays (http://www.stanford.edu/group/nusselab/cgi-bin/wnt/target_genes); 20 μM FH535 upregulated or downregulated multiple genes (, ).
Figure 1 FH535 suppressed the Wnt/β-catenin pathway in pancreatic cancer cells.
Abbreviations: TCF, T-cell factor; h, hours.
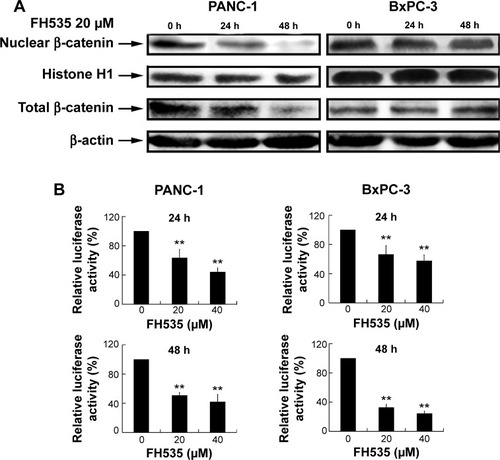

Table 1 Microarray analysis of expression regulation of genes downstream of the Wnt/β-catenin pathway upon 20 μM FH535 treatment
FH535 inhibited pancreatic cancer cell migration
In all, 20 μM FH535 inhibited pancreatic cancer cell migration in a time-dependent manner (). To investigate the mechanisms involved, we analyzed the microarray data to illustrate the expression of genes participating in focal adhesion (, ),Citation13,Citation14 adhesion junctions (, ),Citation15–Citation17 tight junctions (, ),Citation18–Citation23 and cell motility (, ).Citation24–Citation27
Figure 2 FH535 inhibited pancreatic cancer cell migration.
Abbreviation: h, hours.
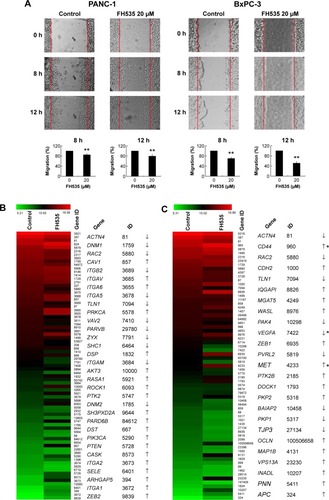
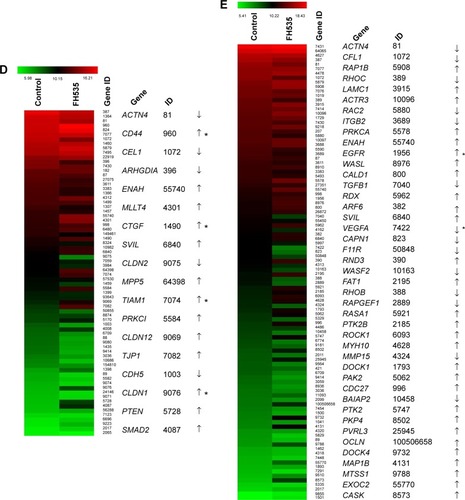
Table 2 Microarray analysis of focal adhesion–related gene expression regulation upon FH535 treatment
Table 3 Microarray analysis of adhesion junction–related gene expression regulation upon FH535 treatment
Table 4 Microarray analysis of tight junction–related gene expression regulation upon FH535 treatment
Table 5 Microarray analysis of cell motility–related gene expression regulation upon FH535 treatment
FH535 inhibited pancreatic cancer cell invasion
The Matrigel invasion assay revealed that FH535-treated cells had significantly decreased invasive capacity as compared with the control cells (), supporting the premise that FH535 inhibits pancreatic cancer cell invasion. Moreover, FH535 inhibited the adhesion ability of pancreatic cancer cells dose-dependently (). We also analyzed the microarray data to explore the changes in the expression of genes involved in the in vitro invasion process, including extracellular matrix degradation (, ), cell adhesion (, ),Citation28,Citation29 and epithelial–mesenchymal transition (EMT) (, ).Citation30–Citation33
Figure 3 FH535 inhibited pancreatic cancer cell invasion.
Abbreviation: EMT, epithelial–mesenchymal transition.
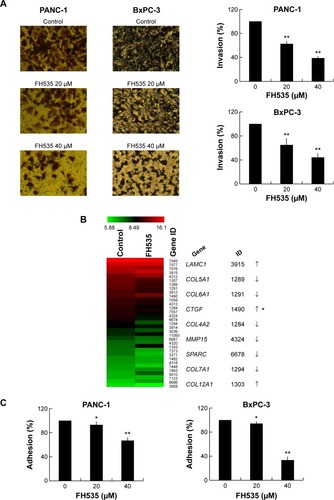
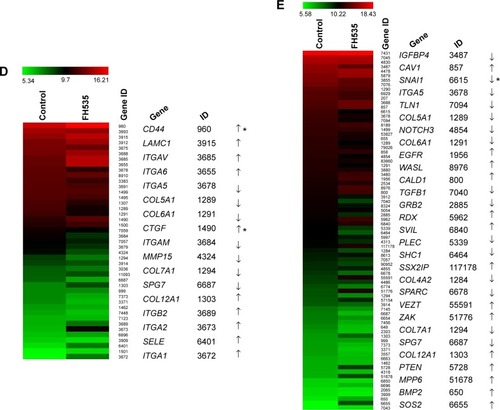
Table 6 Microarray analysis of extracellular matrix degradation–related gene expression regulation upon FH535 treatment
Table 7 Microarray analysis of adhesion molecule–related gene expression regulation upon FH535 treatment
Table 8 Microarray analysis of EMT-related gene expression regulation upon FH535 treatment
FH535 inhibited pancreatic cancer cell growth
Using MTT assay, we evaluated the inhibitory effect of FH535 on pancreatic cancer cell line growth. The proliferation of PANC-1 and BxPC-3 cells cultured for up to 48 hours with FH535 was significantly inhibited time-dependently and dose-dependently as compared to the control cells (). The clone formation assays confirmed the dose-dependent inhibitory effect of FH535 on pancreatic cancer cell growth (). We performed cell cycle analysis to confirm the antimitogenic effect of FH535. FH535 induced G2/M accumulation and decreased the cell population in the G0/G1 and S phases dose-dependently (). The expression profile of the cell cycle–related genes obtained from microarray analyses was analyzed (, ).Citation34
Figure 4 Inhibitory effect of FH535 on pancreatic cancer cell growth.
Abbreviation: h, hours.
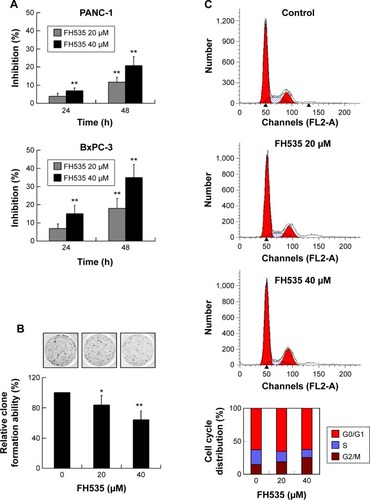

Table 9 Microarray analysis of cell cycle–related gene expression regulation upon 20 μM FH535 treatment
Discussion
It is widely acknowledged that the prognosis of pancreatic cancer is very poor. The canonical Wnt/β-catenin signaling pathway plays a key role in tumor development and dissemination. Classical Wnt signaling pathway causes accumulation of β-catenin in cytoplasm in complex with the transcription factor TCF/LEF that regulates target gene expression.Citation9,Citation35 Dysregulation of Wnt/β-catenin signaling and altered transcription of β-catenin/TCF-regulated genes are found in many cancers,Citation36 including pancreatic cancer.Citation37 In this regard, we focused on characterizing the mechanisms of the anti-tumor effect of FH535 on pancreatic cancer cells.
Western blotting revealed that FH535 did not affect β-catenin expression in BxPC-3 cells. Interestingly, FH535 downregulated the protein level of total β-catenin in the PANC-1 cells, which differed from the results of most previous studies.Citation10 This cell type–dependent downregulation of β-catenin could have been due to the stabilization of axin, which suppresses β-catenin.Citation11 Axin is characterized as a tumor-suppressor gene, and it plays a key role in inhibiting the canonical Wnt pathway by forming molecular complexes with other proteins such as GSK-3β and adenomatous polyposis coli (APC).Citation38 Whether or not β-catenin expression was inhibited, the luciferase reporter assay proved that transcriptional activity of β-catenin pathway was decreased, which was consistent with previous study findings.Citation10
Metastasis, the leading cause of cancer-related death, is a complex process comprising several steps, all of which we found were affected by FH535. First, FH535 inhibited pancreatic cancer cell migration. Microarray analyses revealed that FH535 altered the expression of several migration-related genes, which participate in focal adhesion, adhesion junctions, tight junctions, and/or motility regulation. Among these genes, the focal adhesion–related gene PTEN, considered “the most highly mutated tumor-suppressor gene in the post-p53 era”,Citation39 plays a role in controlling cell migration.Citation40 The loss of PTEN protein expression or function has been reported in many human cancers, including ovarian, endometrial, and prostate carcinoma; breast cancer; and primary gastrointestinal stromal tumor.Citation41,Citation42 We also found that FH535 downregulated the adhesion junction–related gene TLN1, which encodes a cytoskeletal protein that is concentrated in areas of cell–substratum and cell–cell contact. The encoded protein plays a significant role in actin filament assembly and in the spread and migration of various cell types.Citation43,Citation44 TLN1 is codistributed with integrins in the cell surface membrane, aiding the attachment of adherent cells to extracellular matrices and lymphocytes to other cells. In our study, tight junction protein 1 (TJP1), which plays a critical role in cell–cell interaction, proliferation, and differentiation, was upregulated. TJP1 is an important marker of tight junction integrity, which is disrupted in many highly invasive cancers; upregulated TJP1 correlates with favorable survival in breast cancer and gastrointestinal stromal tumor.Citation45,Citation46 The motility-related gene VEGFA significantly increases the motility of pancreatic cancer cells. The vascular endothelial growth factor/vascular endothelial growth factor receptor (VEGF/VEGFR) inhibitors bevacizumab and sunitinib significantly decrease pancreatic cancer cell motility.Citation47 In our study, FH535 not only suppressed VEGFA expression but also inhibited cell motility, suggesting the involvement of a similar mechanism.
To establish metastasis, tumor cells must traverse the basement membrane to reach the connective tissues. Accordingly, we investigated the anti-invasive effect of FH535. The Transwell assay proved that FH535 inhibited invasion. In vitro invasion can be divided into several steps, including matrix adhesion, matrix degradation, and EMT. We analyzed the expression of the genes involved in these steps using microarray and found that FH535 significantly downregulated the cell adhesion molecule ITGA5; ITGA5 knockdown results in decreased adhesion in pancreatic cancer cells.Citation48 The ability of matrix metalloproteinases (MMPs) to degrade extracellular matrix proteins has been well characterized; therefore, they have been studied extensively to elucidate their involvement in both tumor development and progression. Different MMPs play different roles in tumorigenesis. MMP15 appears to be upregulated during colorectal tumorigenesis, and past research has shown stromal localization of MMP15 in the early phases of neoplastic transformation in colorectal cancer.Citation49 In our study, FH535 downregulated MMP15. Epithelial cells are characterized by well-developed junctions and apical–basolateral polarization; on the contrary, mesenchymal cells lack polarization due to the loss of an organized junctional layer. Cell metastasis is correlated with EMT. In the present study, FH535 downregulated Snail, which is upregulated during EMT.Citation50 In human colorectal cancer cells, overexpression of Snail induces not only EMT but also a cancer stem cell–like phenotype, which enhances cell migration and invasion in vitro and increases metastasis formation in vivo.Citation51 Snail also plays an essential role in human pancreatic cancer progression and metastasis.Citation52,Citation53 In the clinical setting, overexpression of Snail was previously associated with poorer prognosis and a more invasive phenotype in many malignancies.Citation54–Citation56 We also detected the downregulation of TGFB1, a classic EMT stimulator.Citation57 TGFB1 overexpression is associated with early recurrence following resection and decreased survival;Citation58 consistent with our study, the suppression of TGFB1 activity in immune-deficient orthotopic mouse models of pancreatic cancer attenuated tumor growth and metastasis.Citation59,Citation60
Besides metastasis, FH535 also induced G2/M arrest and inhibited pancreatic cancer cell proliferation. FH535 significantly upregulated the G2/M regulator gene BCCIP while downregulating the cell cycle regulatory genes CCNG1 and SERTAD1. Human BCCIP, a protein that interacts with BRCA2 and CDKN1A (Cip1, p21), has been implicated in many cellular processes, including cell cycle regulation, DNA recombination and damage repair, telomere maintenance, embryonic development, and genomic stability.Citation61–Citation63 BCCIP knockdown and concomitant p53 deletion causes rapid development of medulloblastomas, which have a wide spectrum of alterations involving the Sonic hedgehog pathway, consistent with the caretaker responsibility of BCCIP in genomic integrity.Citation64 BCCIP expression is downregulated in human ovarian cancer, renal cell carcinoma, and colorectal cancer tissues, suggesting that the gene plays a role in the pathogenesis of these cancers.Citation63 The positive expression rate and intensity of CCNG1 in gastric carcinoma is significantly correlated with tumor differentiation. Elevated amounts of CCNG1 are frequently detected in malignant tissue tumors, including astrocytoma; melanoma; carcinoma of the esophagus, lung, and breast; and cancer of the cervix, uterus, and ovary.Citation65 It plays a pivotal role in hepatocellular carcinoma metastasis and may be a novel prognostic biomarker and therapeutic target.Citation66 SERTAD1 is involved in positive regulation of the cell cycle and proliferation;Citation67,Citation68 accordingly, its expression is upregulated in several tumor types.Citation69,Citation70 Studies indicate that SERTAD1 promotes proliferation by binding to the transcription factor E2F1 and by enhancing its transcriptional activity.Citation71 Experimental overexpression of SERTAD1 provoked hyperproliferation,Citation72 genomic instability,Citation68 and inhibition of apoptosis.Citation73
We demonstrated that FH535 significantly inhibits pancreatic cancer cell metastasis by suppressing migration, invasion, and adhesion and induces the accumulation of cells in the G2/M phase to suppress proliferation. These results suggest that FH535 is a potential candidate for pancreatic cancer treatment. Some of the identified genes that responded to FH535 are well-established direct targets of the Wnt/β-catenin pathway. However, it has not been proven that the other identified genes are located downstream of the pathway. FH535 might affect the expression of these genes through the Wnt/β-catenin pathway indirectly or in a β-catenin independent manner. In fact, FH535 not only antagonizes β-catenin/TCF-mediated transcription but also inhibits recruitment of the coactivators glucocorticoid receptor-interacting protein 1 (GRIP1) and β-catenin to peroxisome proliferator-activated receptor (PPAR)δ and PPARγ,Citation10 suggesting that these mechanisms could also be involved in the anti-cancer effect of FH535.
Acknowledgments
This study was supported by the National Natural Science Foundation of China (grant nos 81472296, 81101867, 81272542, 81200369, and 81372443), the CSPAC-Celgene Foundation, the China International Medical Foundation (grant no CIMF-F-H001-057), the Special Foundation of Clinical Medicine of Jiangsu Provincial Bureau of Science and Technology (grant no BL2014039), the Scientific Research Project of Jiangsu Provincial Bureau of Traditional Chinese Medicine (grant no L213236), the Medical Scientific Research Project of Jiangsu Provincial Bureau of Health (grant no Z201206), the Special Foundation of Wu Jieping Medical Foundation for Clinical Scientific Research (grant nos 320.6753.1225 and 320.6750.12242), the Science and Education for Health Foundation of Suzhou for Youth (grant nos SWKQ1003 and SWKQ1011), the Science and Technology Project Foundation of Suzhou (grant nos SYS201112, SYSD2012137, and SYS201335), the Science and Technology Foundation of Suzhou Xiangcheng (grant nos SZXC2012-70 and XJ201451), and a project founded by the priority academic program development of Jiangsu higher education institutions.
Disclosure
The authors report no conflicts of interest in this work.
References
- JemalASiegelRWardECancer statistics, 2006CA Cancer J Clin200656210613016514137
- LiDXieKWolffRAbbruzzeseJLPancreatic cancerLancet200436394141049105715051286
- BosettiCBertuccioPMalvezziMCancer mortality in Europe, 2005–2009, and an overview of trends since 1980Ann Oncol201324102657267123921790
- CaiHHSunYMMiaoYAberrant methylation frequency of TNFRSF10C promoter in pancreatic cancer cell linesHepatobiliary Pancreat Dis Int20111019510021269942
- IidaJWilhelmsonKLPriceMAMembrane type-1 matrix metalloproteinase promotes human melanoma invasion and growthJ Invest Dermatol2004122116717614962105
- EisenmannKMMcCarthyJBSimpsonMAMelanoma chondroitin sulphate proteoglycan regulates cell spreading through Cdc42, Ack-1 and p130casNat Cell Biol19991850751310587647
- VaidMPrasadRSunQKatiyarSKSilymarin targets beta-catenin signaling in blocking migration/invasion of human melanoma cellsPLoS One201167e2300021829575
- IidaJPeiDKangTMelanoma chondroitin sulfate proteoglycan regulates matrix metalloproteinase-dependent human melanoma invasion into type I collagenJ Biol Chem200127622187861879411278606
- CleversHWnt/beta-catenin signaling in development and diseaseCell2006127346948017081971
- HandeliSSimonJAA small-molecule inhibitor of Tcf/beta-catenin signaling down-regulates PPARgamma and PPARdelta activitiesMol Cancer Ther20087352152918347139
- IidaJDorchakJLehmanJRFH535 inhibited migration and growth of breast cancer cellsPLoS One201279e4441822984505
- RenJWangRSongHHuangGChenLSecreted frizzled related protein 1 modulates taxane resistance of human lung adenocarcinomaMol Med20142016417824643460
- HanniganGTroussardAADedharSIntegrin-linked kinase: a cancer therapeutic target unique among its ILKNat Rev Cancer200551516315630415
- LegateKRMontanezEKudlacekOFasslerRILK, PINCH and parvin: the tIPP of integrin signallingNat Rev Mol Cell Biol200671203116493410
- PalaciosFPriceLSchweitzerJCollardJGD’Souza-SchoreyCAn essential role for ARF6-regulated membrane traffic in adherens junction turnover and epithelial cell migrationEMBO J200120174973498611532961
- PeceSGutkindJSE-cadherin and Hakai: signalling, remodeling or destruction?Nat Cell Biol200244E72E7411944035
- D’Souza-SchoreyCDisassembling adherens junctions: breaking up is hard to doTrends Cell Biol2005151192615653074
- TsukitaSFuruseMItohMMultifunctional strands in tight junctionsNat Rev Mol Cell Biol20012428529311283726
- ChengCYMrukDDCell junction dynamics in the testis: sertoli-germ cell interactions and male contraceptive developmentPhysiol Rev200282482587412270945
- MatterKBaldaMSSignalling to and from tight junctionsNat Rev Mol Cell Biol20034322523612612641
- BaldaMSMatterKEpithelial cell adhesion and the regulation of gene expressionTrends Cell Biol200313631031812791297
- BazzoniGDejanaEEndothelial cell-to-cell junctions: molecular organization and role in vascular homeostasisPhysiol Rev200484386990115269339
- FuruseMTsukitaSClaudins in occluding junctions of humans and fliesTrends Cell Biol200616418118816537104
- LinderSThe matrix corroded: podosomes and invadopodia in extracellular matrix degradationTrends Cell Biol200717310711717275303
- ChhabraESHiggsHNThe many faces of actin: matching assembly factors with cellular structuresNat Cell Biol20079101110112117909522
- McEverRPZhuCRolling cell adhesionAnnu Rev Cell Dev Biol20102636339619575676
- ParsonsJTHorwitzARSchwartzMACell adhesion: integrating cytoskeletal dynamics and cellular tensionNat Rev Mol Cell Biol201011963364320729930
- MullinsRFSkeieJMFolkJCEvaluation of variants in the selectin genes in age-related macular degenerationBMC Med Genet2011125821521525
- CirizaJGarcia-OjedaMEExpression of migration-related genes is progressively upregulated in murine lineage-Sca-1+c-Kit+ population from the fetal to adult stages of developmentStem Cell Res Ther2010121420637061
- KaartinenVVonckenJWShulerCAbnormal lung development and cleft palate in mice lacking TGF-beta 3 indicates defects of epithelial-mesenchymal interactionNat Genet19951144154217493022
- TimmermanLAGrego-BessaJRayaANotch promotes epithelial-mesenchymal transition during cardiac development and oncogenic transformationGenes Dev20041819911514701881
- Moreno-BuenoGCubilloESarrióDGenetic profiling of epithelial cells expressing E-cadherin repressors reveals a distinct role for snail, slug, and E47 factors in epithelial-mesenchymal transitionCancer Res200666199543955617018611
- YangJWeinbergRAEpithelial-mesenchymal transition: at the crossroads of development and tumor metastasisDev Cell200814681882918539112
- HoffmanAEZhengTBaYPhenotypic effects of the circadian gene cryptochrome 2 on cancer-related pathwaysBMC Cancer20101011020334671
- TetsuOMcCormickFBeta-catenin regulates expression of cyclin D1 in colon carcinoma cellsNature1999398672642242610201372
- FearonERPARsing the phrase “all in for axin” – Wnt pathway targets in cancerCancer Cell200916536636819878868
- WhiteBDChienAJDawsonDWDysregulation of Wnt/beta-catenin signaling in gastrointestinal cancersGastroenterology2012142221923222155636
- MacDonaldBTTamaiKHeXWnt/beta-catenin signaling: components, mechanisms, and diseasesDev Cell200917192619619488
- Di CristofanoAPandolfiPPThe multiple roles of PTEN in tumor suppressionCell2000100438739010693755
- WaiteKAEngCProtean PTEN: form and functionAm J Hum Genet200270482984411875759
- ChalhoubNBakerSJPTEN and the PI3-kinase pathway in cancerAnnu Rev Pathol2009412715018767981
- WangHChenPLiuXXPrognostic impact of gastrointestinal bleeding and expression of PTEN and Ki-67 on primary gastrointestinal stromal tumorsWorld J Surg Oncol2014128924712384
- TangHYaoLTaoXmiR-9 functions as a tumor suppressor in ovarian serous carcinoma by targeting TLN1Int J Mol Med201332238138823722670
- SakamotoSMcCannRODhirRKyprianouNTalin1 promotes tumor invasion and metastasis via focal adhesion signaling and anoikis resistanceCancer Res20107051885189520160039
- SommersCLByersSWThompsonEWTorriJAGelmannEPDifferentiation state and invasiveness of human breast cancer cell linesBreast Cancer Res Treat1994312–33253357881109
- ZhuHLuJWangXUpregulated ZO-1 correlates with favorable survival of gastrointestinal stromal tumorMed Oncol201330363123820955
- DoiYYashiroMYamadaNAmanoRNodaSHirakawaKVEGF-A/VEGFR-2 signaling plays an important role for the motility of pancreas cancer cellsAnn Surg Oncol20121982733274322207048
- WalshNClynesMCrownJO’DonovanNAlterations in integrin expression modulates invasion of pancreatic cancer cellsJ Exp Clin Cancer Res20092814019825166
- SenaPMarianiFMarzonaLMatrix metalloproteinases 15 and 19 are stromal regulators of colorectal cancer development from the early stagesInt J Oncol201241126026622576687
- WuYZhouBPSnail: more than EMTCell Adh Migr20104219920320168078
- FanFSamuelSEvansKWOverexpression of snail induces epithelial-mesenchymal transition and a cancer stem cell-like phenotype in human colorectal cancer cellsCancer Med20121151623342249
- HotzBArndtMDullatSBhargavaSBuhrHJHotzHGEpithelial to mesenchymal transition: expression of the regulators snail, slug, and twist in pancreatic cancerClin Cancer Res200713164769477617699854
- von BurstinJEserSPaulMCE-cadherin regulates metastasis of pancreatic cancer in vivo and is suppressed by a SNAIL/HDAC1/HDAC2 repressor complexGastroenterology20091371e361e365
- ShinNRJeongEHChoiCIOverexpression of snail is associated with lymph node metastasis and poor prognosis in patients with gastric cancerBMC Cancer20121252123151184
- KuoKTChouTYHsuHSChenWLWangLSPrognostic significance of NBS1 and snail expression in esophageal squamous cell carcinomaAnn Surg Oncol201219suppl 3S549S55721881923
- van NesJGde KruijfEMPutterHCo-expression of SNAIL and TWIST determines prognosis in estrogen receptor-positive early breast cancer patientsBreast Cancer Res Treat20121331495921796367
- WuYZhouBPNew insights of epithelial-mesenchymal transition in cancer metastasisChin J Biochem Biophys2008407643650
- FriessHYamanakaYBüchlerMEnhanced expression of transforming growth factor beta isoforms in pancreatic cancer correlates with decreased survivalGastroenterology19931056184618568253361
- Rowland-GoldsmithMAMaruyamaHKusamaTRalliSKorcMSoluble type II transforming growth factor-beta (TGF-beta) receptor inhibits TGF-beta signaling in COLO-357 pancreatic cancer cells in vitro and attenuates tumor formationClin Cancer Res2001792931294011555612
- MelisiDIshiyamaSSclabasGMLY2109761, a novel transforming growth factor beta receptor type I and type II dual inhibitor, as a therapeutic approach to suppressing pancreatic cancer metastasisMol Cancer Ther20087482984018413796
- MisraSSharmaSAgarwalACell cycle-dependent regulation of the bi-directional overlapping promoter of human BRCA2/ZAR2 genes in breast cancer cellsMol Cancer201095020202217
- LuHYueJMengXNickoloffJAShenZBCCIP regulates homologous recombination by distinct domains and suppresses spontaneous DNA damageNucleic Acids Res200735217160717017947333
- LiuXCaoLNiJDifferential BCCIP gene expression in primary human ovarian cancer, renal cell carcinoma and colorectal cancer tissuesInt J Oncol20134361925193424101097
- HuangYYDaiLGainesDBCCIP suppresses tumor initiation but is required for tumor progressionCancer Res201373237122713324145349
- CuiXYuLWangYThe relationship between cyclin G1 and survival in patients treated surgically for HCCHepatogastroenterology20136012115315922835824
- WenWDingJSunWCyclin G1-mediated epithelial-mesenchymal transition via phosphoinositide 3-kinase/Akt signaling facilitates liver cancer progressionHepatology20125561787179822271581
- LiJMuscarellaPJooSHDissection of CDK4-binding and transactivation activities of p34(SEI-1) and comparison between functions of p34(SEI-1) and p16(INK4A)Biochemistry20054440132461325616201750
- TangDJHuLXieDOncogenic transformation by SEI-1 is associated with chromosomal instabilityCancer Res200565156504650816061626
- van DekkenHAlersJCRiegmanPHRosenbergCTilanusHWVissersKMolecular cytogenetic evaluation of gastric cardia adenocarcinoma and precursor lesionsAm J Pathol200115861961196711395372
- TangTCShamJSXieDIdentification of a candidate oncogene SEI-1 within a minimal amplified region at 19q13.1 in ovarian cancer cell linesCancer Res200262247157716112499249
- HsuSIYangCMSimKGHentschelDMO’LearyEBonventreJVTRIP-Br: a novel family of PHD zinc finger- and bromodomain-interacting proteins that regulate the transcriptional activity of E2F-1/DP-1EMBO J20012092273228511331592
- SugimotoMNakamuraTOhtaniNRegulation of CDK4 activity by a novel CDK4-binding protein, p34(SEI-1)Genes Dev199913223027303310580009
- HongSWKimCJParkWSp34SEI-1 inhibits apoptosis through the stabilization of the X-linked inhibitor of apoptosis protein: p34SEI-1 as a novel target for anti-breast cancer strategiesCancer Res200969374174619176394