Abstract
Objectives
Although human respiratory metabolism is characterized by the mitochondrial electron transport chain, some organisms present a “branched respiratory chain.” This branched pathway includes both a classical and an alternative respiratory chain. The latter involves an alternative oxidase. Though the Plasmodium falciparum alternative oxidase is not yet identified, a specific inhibitor of this enzyme, salicylhydroxamic acid (SHAM), showed a drug effect on P. falciparum respiratory function using oxygen consumption measurements. The present study aimed to highlight the metabolic pathways that are affected in P. falciparum following SHAM exposure.
Design
A proteomic approach was used to analyze the P. falciparum proteome and determine the metabolic pathways altered following SHAM treatment. To evaluate the SHAM effect on parasite growth, the phenotypic alterations of P. falciparum after SHAM or/and hyperoxia exposure were observed.
Results
After SHAM exposure, 26 proteins were significantly deregulated using a fluorescent two dimensional-differential gel electrophoresis. Among these deregulated proteins, some were particularly involved in energetic metabolism. And the combinatory effect of SHAM/hyperoxia seems deleterious for the growth of P. falciparum.
Conclusion
Our results indicated that SHAM appears to activate glycolysis and decrease stress defense systems. These data provide a better understanding of parasite biology.
Background
The malaria parasite Plasmodium falciparum possesses a mitochondrion with specific structural and physiological features.Citation1–Citation4 Unlike most other eukaryotes, malarial mitochondrial electron transport chain (mETC) activity appears to be primarily involved in de novo pyrimidine biosynthesis through the activity of dihydroorotate dehydrogenase rather than through an adenosine triphosphate (ATP) source.Citation4–Citation6 Intraerythrocytic stages of malaria parasites have been considered for a long time to rely primarily on anaerobic glycolysis for ATP production. However, recently five subunits of the F1 subcomplex and four F0 subunits of the mitochondrial F0F1 ATP synthase have been reported in the P. falciparum genome, supporting the fact that oxidative phosphorylation could be functional.Citation7 As parasite respiratory metabolism is distinct from human respiration, diverse inhibitors of the major complexes of the mETC have been used in chemotherapyCitation8 and validated the importance of the mitochondrion in the growth of the asexual intraerythrocytic stage of P. falciparum.Citation9,Citation10 Moreover, the metabolic changes of the mitochondrion contribute to environmental adaptations of parasites during its life cycle.Citation11 Effectively, in vivo, P. falciparum life cycle involves two hosts: the mosquito and humans during which the parasite is subjected to varying oxygen levels (from 5% oxygen in human venous blood to 13% oxygen in the human lungs and 21% oxygen in mosquito salivary glands).
Study of the rotenone drug effect underlined the divergence of the first component (complex I) of the P. falciparum mETC.Citation12–Citation14 The eukaryotic complex I is classically composed of a rotenone-sensitive nicotinamide adenine dinucleotide hydride (NADH) dehydrogenase. In contrast, P. falciparum encodes a rotenone-insensitive alternative complex I (type II NADH dehydrogenase, PfNDH2). This complex I is comparable to type II NADH dehydrogenase found in plants. Not surprisingly, parasite mitochondria have an endosymbiotic origin probably resulting from algal symbiont into a protoeukaryote.Citation15–Citation17 Moreover, other mitochondrial drugs such as atovaquone inhibit electron transfer at the level of the bc1 complex (complex III) by interfering with the ubiquinol oxidation site of cytochrome b. The drug effect is lethal for the parasite, presumably by interruption of essential links to de novo pyrimidine biosynthesis and to the collapse of the mitochondrial transmembrane potential.Citation18,Citation19 This drug is already currently used in endemic regions in combination with proguanil (Malarone®).Citation20 Also, the combination of atovaquone and salicylhydroxamic acid (SHAM), a specific inhibitor of alternative oxidase (AOX), inhibits parasite growth synergistically.Citation21,Citation22 The AOX, which is absent from mammals, has been most extensively studied in higher plants. The plant respiratory pathway includes an alternative respiratory chain, cyanide-resistant, and the classical mETC, cyanide-sensitive. The AOX does not appear to contribute directly to mitochondrial transmembrane potential or the energy balance of the cell.Citation23,Citation24 However, it can contribute indirectly by accepting electrons from enzymes that provide electrons to coenzyme Q for the preservation of tricarboxylic acid cycle (TCA) turnover and the balance of carbon metabolism and electron transport. From the mitochondrial transmembrane potential and oxygen consumption measurements in presence of P. falciparum mETC inhibitors, the evidence of an alternative respiratory pathway in which electrons are transferred directly from coenzyme Q to oxygen was underlined.Citation25 This cyanide-resistant alternative respiratory pathway was inhibited by two inhibitors, propyl gallate and SHAM, which are specific inhibitors of AOX activity in other organisms.Citation26–Citation28 The AOX gene has been characterized in several organisms such as plants, yeast, bacterium, and notably from other parasites, Trypanosoma brucei and Cryptosporidium parvum.Citation28–Citation30 Nevertheless, until now, no gene encoding AOX has been detected in the P. falciparum genome but 60% of the predicted genes could not be assigned to orthologous functions.
In the present work, a two dimensional-differential gel electrophoresis (2D-DIGE) approach, combined with mass spectrometry (MS) analysis, was used to define the consequences of SHAM on the P. falciparum proteome. Studies were previously reported that the hyperoxia induces a cycle delay on P. falciparum of 4 hours with a biological adaptation of the parasite to next cycle.Citation11,Citation31 The deleterious effect of this drug on the in vitro parasite growth in hyperoxia, the involved metabolic pathways, and its mode of action on glycolysis are discussed, supporting the hypothesis that the energetic metabolism may indeed provide an attractive chemotherapeutic target.
Methods
SHAM or/and hyperoxia exposure of P. falciparum cultures
The in vitro antimalarial activity of SHAM was determined using the isotopic semi microtest method as previously described by Desjardins et al.Citation32 Drug concentrations were from 0.5 to 600 μM. The IC50, ie, the drug concentration corresponding to 50% of the uptake of [3H] hypoxanthine by the parasite in drug-free control wells, was determined by nonlinear regression analysis of log dose-response curves.
The 3D7 P. falciparum strain was cultured in human A+ erythrocytes, as previously described.Citation33 Cultures were tightly synchronized (with 4 hour intervals) using combined D-sorbitol and CS columns on a VarioMACS (Miltenyi Biotec, Paris, France) according to standard procedures.Citation34,Citation35
To evaluate the effect of SHAM, hyperoxia, and the combinatory effect, cultures were adapted to hyperoxic conditions (21% oxygen, 5% carbon dioxide, 74% nitrogen gas mixture) and regularly synchronized during six P. falciparum parasites’ cycles before the addition of SHAM. The effects of SHAM alone and combined SHAM/hyperoxia were observed at 0, 24, 32, 48, and 78 hours (0 hours corresponding to SHAM addition at the ring stage). Viability, parasitemia, and morphology were monitored daily by examining blood smears stained with RAL 555 (RAL Diagnostics, Martillac, France) from SHAM addition at the ring stage.
To test the SHAM effects on P. falciparum proteome (3% parasitemia and 6% hematocrit), the drug was added to 25 mL culture medium at IC50 and the parasites were further incubated for 12 hours and harvested. For each proteomic experiment, four biological replicates were performed.
Protein extraction
After SHAM pressure, parasitized erythrocytes (late ring stages aged 16–20 hours) were washed three times in phosphate buffered saline (PBS) medium (Invitrogen, Cergy Pontoise, France) and lysed in cold water-saponin (0.1%, Sigma, St Louis, MO) for 10 minutes. The lysate was then centrifuged at 1500 g for 5 minutes. The supernatant was discarded and the pellet containing free parasites was recovered by washing in cold PBS medium followed by a centrifugation step (1500 g for 5 minutes). The free parasites were washed until the supernatant became colorless. The pellet was then suspended in 4% (w/v) CHAPS (Sigma) and disrupted by ultrasonication (Vibracell 72412; Bioblock Scientific, Illkirch, France) 60 seconds on ice at maximum amplitude. The lysate was then centrifuged at 16,100 g for 15 minutes. The supernatant was further precipitated with 100% acetone (Sigma). The protein concentration for each sample was estimated using the BioRad Lowry-based DC assay (BioRad Laboratories, Hercules, CA), according to the manufacturer’s instructions. Total proteins were suspended in standard cell lysis buffer (7 M urea, 2 M thiourea, 4% CHAPS, 30 mM Tris base, pH 8.5 [Sigma]) to obtain a protein concentration adjusted to 2.5 μg/μL. Protein samples were minimally labeled with CyDye according to the manufacturer’s protocols (GE Healthcare, Piscataway, NJ).Citation36
Briefly, protein extracts (50 μg) were labeled with 400 pmol of CyDye, freshly dissolved in anhydrous dimethyl formamide (Sigma) and incubated on ice for 30 minutes in the dark. The reaction was quenched with 1 μL of free lysine (10 nM, Sigma) by incubating for 10 minutes on ice. An equal volume of 2× sample buffer (8 M urea, 2 M thiourea, 4% (w/v) CHAPS, 10 mM Dithiothreitol (DTT) and 1% (v/v) IPG buffer 3–10 [GE Healthcare]) was added to the CyDye-labeled sample. The mixture of labeled proteins was then separated by 2D-DIGE.
2D electrophoresis of parasite proteins, image analysis, and in-gel digestion
Isoelectric focusing was performed on 18 cm pH 3–10 linear IPG strips (GE Healthcare). Destreak buffer containing 1% (v/v) IPG buffer pH 3–10 was used for overnight rehydration of IPG strips. The samples were applied at the acidic end of the IPG strip using a cup-loading technique. Isoelectric focusing was carried out on an Ettan IPGphor II (GE Healthcare) electrophoresis unit at 20°C for a total of 45 kVh (ramp to 300 V in 3 hours, ramp to 1000 V in 6 hours, ramp to 8000 V in 3 hours, hold at 8000 V for 4 hours). IPG strips were equilibrated in a equilibration buffer containing 50 mM Tris hydrochloride, pH 8.6, 6 M urea, 2% sodium dodecyl sulfate (SDS) and 30% glycerol supplemented with 1% (w/v) DTT for 15 minutes at room temperature, followed by protein alkylation (carbamidomethylation) in the same equilibration buffer containing 2.5% (w/v) iodoacetamide instead of DTT for 15 minutes at room temperature. IPG strips were then placed on the top of 10% uniform polyacrylamide gels. Strips were overlaid with 0.5% agarose in 1x running buffer containing bromophenol blue, and the proteins were further separated by SDS-polyacrylamide gel electrophoresis (10 W per gel) at 20°C in the Ettan DALTsix Electrophoresis System (GE Healthcare). After electrophoresis, the gels with CyDye-labeled proteins were directly imaged using a Typhoon™ Trio image scanner (GE Healthcare, Buckinghamshire, UK).
After 2D-DIGE, gels were scanned on the Typhoon™ Trio image scanner at different excitation wavelengths (Cy3, 580 BP 30/green [532 nm]; Cy5, 670 BP 30/red [633 nm]; Cy2, 520 BP 40/blue [488 nm]). The intensity was adjusted to ensure that the maximum volume of each image was within 60,000–80,000 U. Analysis of 2D-DIGE was performed using DeCyder 6.5 software (GE Healthcare), according to the manufacturer’s recommendations. Briefly, the differential in-gel analysis module was used to detect the intra-gel spots and the biological variation analysis module to match different gels using the in-gel standard. The paired t-test was used for statistical analysis of the data. Protein spots that were expressed differentially between two experimental conditions (|ratio| ≥ 1.5, P ≤ 0.05 t-test) were marked with master gel numbers. Based on DeCyder 6.5, analysis spots of interest were excised from gels using a Shimadzu Xcise automated gel processing platform (Shimadzu Biotech, Kyoto, Japan). Excised spots were prepared as described previously and then stored at −20°C before analysis by MS.Citation37
MS analysis
The samples were analyzed by nanoscale capillary liquid chromatography-tandem MS (nano LC-MS/MS). Purification and analysis were performed on a C18 capillary column using a CapLC system (Waters, Milford, MA) coupled to a hybrid quadrupole orthogonal acceleration time-of-flight tandem mass spectrometer (Q-TOF Ultima; Waters). Chromatographic separation was conducted on a reversed-phase capillary column (Atlantis™ dC18, 3 μm, 75 μm × 150 mm Nano Ease™; Waters) with a 180–200 nL/minute flow. The gradient profile consisted of a linear gradient from 95% A (water, 0.1% formic acid) to 60% B (80% acetonitrile, 0.1% formic acid) in 60 minutes followed by a linear gradient by 95% B in 10 minutes. Mass data acquisitions were piloted by MassLynx 4.0 software (Micromass Ltd, Manchester, UK) using automatic switching between MS and MS/MS modes. The internal parameters of Q-TOF were set as follows. The electrospray capillary voltage was set to 3.2 kV, the cone voltage was set to 30 V, and the source temperature was set to 80°C. The MS survey scan was m/z 400–1,300 with a scan time of 1 second and an interscan time of 0.1 second. When the intensity of a peak rose above a threshold of 15 counts, tandem mass spectra were acquired. Normalized collision energies for peptide fragmentation were set using the charge-state recognition files for +2 and +3 peptide ions. The scan range for MS/MS acquisition was from m/z 50 to 1500 with a scan time of 1 second and an interscan time of 0.1 second. Fragmentation was performed using argon as the collision gas and with the collision energy profile optimized for various mass ranges and charges of precursor ions. Mass data collected during a nano LC-MS/MS analysis were processed using ProteinLynx Global Server 2.2 software (Waters) with the following parameters: no background subtraction, smooth 3/2 Savitzky Golay, and no deisotoping to generate peak lists in the Micromass pkl format. Pkl files were then fed into a local search engine Mascot Daemon v2.2.2 (Matrix Science, London, UK). The data were searched against the Homo sapiens (218,356 sequences) and P. falciparum (13,110 sequences) National Center for Biotechnology Information nonredundant (NCBInr) protein databases (March 15, 2010). Search parameters allowed for one missed tryptic cleavage site, the carbamidomethylation of cysteine, and the possible oxidation of methionine; precursor and product ion mass error tolerance was <0.2 Da. All identified proteins had a Mascot score greater than 34 and 43 for P. falciparum and H. sapiens, respectively, corresponding to statistically significant identification (P < 0.05).
Results
SHAM and hyperoxia combinatory effect on the asexual blood cycle of P. falciparum
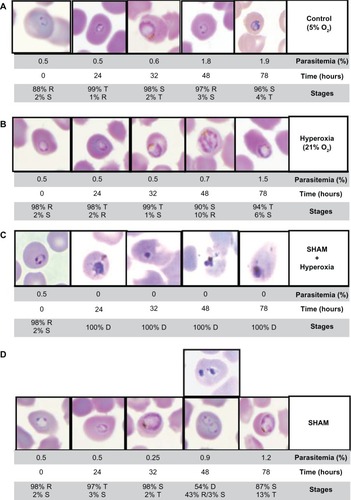
To study the deleterious effect of SHAM on asexual blood stages of P. falciparum in hyperoxia, 21% oxygen-adapted cultures of 3D7 were exposed to SHAM IC50. The parasitemia and percentages of the different stages of parasites were evaluated by blood smears at 0, 24, 32, 48, and 78 hours (0 hours corresponding to SHAM addition at the ring stage). In in vitro culture under hyperoxic conditions, the 3D7 P. falciparum strain had a life cycle of 49 hours against 45 hours in normal conditions (). Parasite exposure to 21% oxygen and SHAM pressure resulted in morphologic alterations involving cellular degeneration during the first 24 hours ( and ). These results suggest that SHAM addition to 3D7 P. falciparum strain under oxidative stress had lethal effects from the beginning of the parasitic cycle and that the life cycle does not continue beyond the ring stage. A previous study demonstrated that hyperoxia induces a delay of P. falciparum cycle of 4 hours with an energetic metabolism adaptation crucial to parasite survival.Citation11 Consequently, the SHAM/hyperoxia combinatory effect appeared to result in cellular death, suggesting that the adaptive pathway for hyperoxia was disturbed under SHAM pressure.
Figure 1 Morphologic alterations of Plasmodium falciparum following salicylhydroxamic acid (SHAM) pressure. Phenotypic changes of P. falciparum asexual blood stages were observed during two parasite cycles under (A) normoxia, (B) hyperoxia (ie, 21% oxygen), (C) SHAM/hyperoxia conditions, and (D) SHAM pressure. The parasitemia and parasitic stages were evaluated by blood smears at 0, 24, 32, 48, and 78 hours. After SHAM addition following hyperoxic exposure, significant morphological alterations were observed during the two cycles of parasites. The different intraerythrocytic stages and their corresponding percentages are indicated as follows: ring (R), trophozoites (T), schizonts (S), and parasite’s death (D).
Effect of SHAM on P. falciparum asexual blood cycle
To establish the experimental design for the proteome study, the SHAM IC50 and the action timing of the SHAM should be determined.
Consistent with previously published results,Citation21,Citation38 SHAM inhibited growth of the 3D7 strain with an average IC50 of 246 +/−4 μM.
To determine the SHAM effect on asexual blood stages of P. falciparum, the drug was added at the IC50 concentration on synchronous cultures of 3D7 at the ring stage. The parasitemia and parasitic stage percentages were monitored during two life cycles by blood smears at 0, 24, 32, 48, and 78 hours in three independent experiments (0 hours corresponding to SHAM addition at the ring stage). The parasitemia of SHAM-treated cultures was low at 32 hours compared to untreated parasites ( and , 0.6% parasitemia vs 0.25%, respectively). At the end of 48 hours under SHAM exposure, 54% of the parasites were degenerated but the parasites not affected by SHAM continued their cycle without phenotypic changes and lengthening of the parasitic cycle. Thus, the SHAM perturbed the parasite growth between the 24th and 32nd hours of the cycle at IC50 concentration.
SHAM-alteration to the P. falciparum proteome
To study the SHAM effect on P. falciparum, a proteomic approach was performed in synchronized parasites exposed to SHAM IC50. To identify altered P. falciparum proteins, 2D-DIGE experiments coupled to MS were performed. Four independent cultures of untreated and SHAM-treated P. falciparum were included in this analysis. After protein separation by 2D-DIGE using pH 3–10 IPG strips and homogeneous 10% SDS-polyacrylamide gel electrophoresis, each gel was individually imaged, and all gel images were analyzed using DeCyder 6.5 software. Among 1998 matched protein spots, 18 spots were differentially modulated (|fold change| ≥ 1.5, P ≤ 0.05 t-test) between the control and SHAM treatment (seven and eleven spots were up- and down-modulated, respectively; ). The corresponding protein spots were excised from gels, analyzed by LC-MS/MS, and searched against P. falciparum and H. sapiens databases (NCBInr). Eight protein spots not fulfilling the selection criteria were still selected and so submitted to MS analysis. Their P values were significant or very close to significance with |FC| closed from 1.5 (). With this last selection, a total of 26 protein spots were identified by MS that correspond to 10 P. falciparum and 16 H. sapiens specific proteins (). Among these eight last selected proteins, six correspond to isoforms supporting the abundance variation of significant deregulated proteins. Thus, some proteins were detected in more than one spot (eg, four for glyceraldehyde-3-phosphate dehydrogenase, three for catalase), suggesting the presence of different deregulated isoforms. So, the number of distinct proteins identified was five for P. falciparum and nine for H. sapiens (). The importation of human proteins into the parasite during erythrocytic cycle is a well-known phenomenon, as previously described in other studies.Citation39 The deregulated protein spots need to be confirmed on other wild P. falciparum strains or isolates.
Figure 2 Salicylhydroxamic acid (SHAM) effect on Plasmodium falciparum 3D7 strain proteome. Representative data from a two dimensional-differential gel electrophoresis experiment using a 10% homogenous sodium dodecyl sulfate polyacrylamide gel with pH range 3–10 are shown. The proteins from untreated P. falciparum parasites or treated parasites with 250 μM SHAM were labeled with Cy3 and Cy5, respectively. As determined by DeCyder 6.5 software, protein spots that were up- and down-expressed on P. falciparum under SHAM treatment (|FC| ≥ 1.5, P ≤ 0.05 t-test) were marked with master numbers (). Bold and italicized numbers correspond, respectively, to identified proteins from P. falciparum and Homo sapiens.
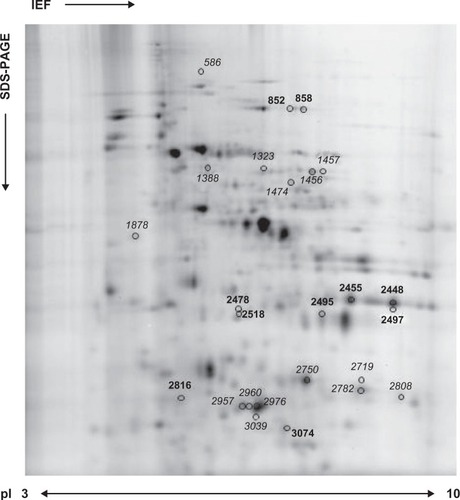
Table 1 Proteins identified from the two dimensional-differential gel electrophoresis analysis in salicylhydroxamic acid-treated Plasmodium falciparum
To determine the metabolic pathways perturbed under SHAM pressure, the identified proteins were classified using the NCBI Clusters of Orthologous Groups of proteins database (). Among the functional categories, some proteins were predicted to play a role in glycolysis, chaperone-assisted protein folding, and redox metabolism. Among the antioxidant proteins, human catalase, carbonic anhydrase I, and carbonic anhydrase II were altered in SHAM-treated P. falciparum.
Discussion
SHAM and hyperoxia exposure affects P. falciparum growth
Previous studies provided evidence for the existence of a P. falciparum AOXCitation40 and supported the ability of SHAM to potentiate the activity of atovaquone, a complex III inhibitor, in in vitro chemosusceptibility assays.Citation21 Other studies have examined the effects of cyanide, a complex IV inhibitor, on P. falciparum respiratory metabolism using a polarographic assay.Citation38 In the presence of high concentrations of cyanide, 25% of the parasite oxygen consumption was residual, suggesting that the parasite exploits a cyanide-resistant alternative respiratory pathway. This residual oxygen consumption was totally inhibited by SHAM (coenzyme Q analog), a specific inhibitor of AOX enzymatic activity involved in this alternative pathway.Citation13
Our results indicate that SHAM (IC50 250 μM) induced the death of effectively 54% of the parasites but in hyperoxic conditions and to the same IC50, parasite death is 100%. Therefore, in hyperoxic conditions, SHAM exposure has a deleterious effect on parasite survival. SHAM seems to inhibit the metabolic adaptation involved in hyperoxia, which is essential to parasite survival.
Consequently, as described in plants,Citation23,Citation24 SHAM action seems to prevent the decrease in the levels of reactive oxygen species (ROS), which are exceedingly produced in hyperoxia by the mETC.Citation42 Because the AOX activity reduces oxygen, its activity should be dependent on the oxygen concentration and also on the coenzyme Q concentration and its redox state.Citation23 This alternative pathway could provide a regulatory function for excess electrons when the mETC is saturated (as during hyperoxia) and thus, decrease ATP production (unpublished data). Additionally, this “electron leak” mechanism may improve potential repression of the TCA cycle by elevated levels of NADH and ATP.Citation24
SHAM alters the P. falciparum proteome
To investigate the metabolic pathways altered by the activity of SHAM, a comparative study on the P. falciparum proteome was performed using 2D-DIGE coupled to MS for identification. The experimental design (parasite stage, drug dose, incubation time) was chosen based on our preliminary results. In our study, SHAM was exposed at the IC50 concentration to ring stage parasites for 12 hours.
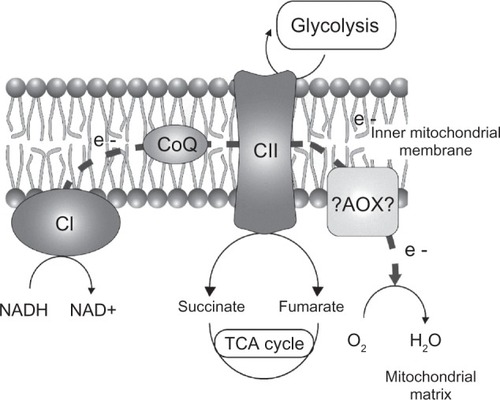
Most SHAM-altered proteins are involved in glycolysis and redox metabolism. The major implication of glycolytic enzymes (glyceraldehyde-3-phosphate dehydrogenase, L-lactate dehydrogenase, enolase, and phosphoglycerate mutase) suggests the need to sustain the major energy dependence of the parasite on glycolysis. Although the function of the P. falciparum mitochondrion remains unclear, its contribution to ATP production is considered to be minimal in asexual stages. P. falciparum relies principally on anaerobic glycolysis for energy production.Citation4 Therefore, the parasites are dependent on glycolysis in a microaerophilic environment while in hyperoxia conditions; our previous results showed that the respiratory metabolism is favored at the expense of glycolysis.Citation11 This observation could explain the lethal effect of SHAM in hyperoxic conditions. The NADH generated during glycolysis are reoxidized through glycerol-3-phosphate in the cytosol and by PfNDH2 in the mitochondria.Citation43,Citation44 In plants, the alternative respiratory pathway involves an alternative NADH dehydrogenase, coenzyme Q, and AOX.Citation23 Mitochondrial AOX exists under two forms, covalent and noncovalent homodimers.Citation45 The oxidized covalent form of the enzyme is inactive, whereas reducing the disulfide bond generated by NADH activates the enzyme, possibly mediated by a glutathione/thioredoxin coupling system.Citation46 Interestingly, glycolysis activation under SHAM pressure could be the result of a metabolic adaptation in a microaerophilic environment. The end products of P. falciparum glycolysis are pyruvate, lactate, and glycerol. Firstly, pyruvate is an allosteric activator of AOX counterbalancing the AOX inhibition by SHAM. Under conditions of partial respiration, accumulation of pyruvate would result in increased NADH, which could potentially activate AOX.Citation2 Secondly, lactate, via L-lactate dehydrogenase, may be a substrate for the mETC under stress conditions.Citation43 Finally, glycerol production by the glycerol-3-phosphate permitted the reoxidation of NADH and the activity of the mETCCitation47 (). In a microaerophilic environment, the activation of glycolysis is concordant with the role of glycolytic metabolism in the reoxidation of NADH,Citation48 an essential cofactor of the alternative respiratory pathway in T. bruceiCitation26 and C. parvum.Citation44 Indeed, T. brucei brucei survives using an anaerobic pathway while T. brucei brucei AOX is inhibited.Citation26,Citation41 Collectively, these results showed that SHAM, via its action on glycolysis metabolism, appears to play a role in NAD/NADH balance. The mechanism of regulation of AOX activity involved several compounds, including those in redox states (NAD/NADH) and allosteric effectors (pyruvate).Citation49 Thus, the existence of an AOX function in P. falciparum could be beneficial for the survival of the parasite in hyperoxic environments as in the salivary glands of mosquitoes or perhaps in the human lungs.
Figure 3 A schematic representation of the Plasmodium falciparum metabolic pathways perturbed under salicylhydroxamic acid treatment. Glycolysis and mitochondrial respiratory chain pathways based on the Ginsburg website are represented.Citation50
SHAM treatment leads to an inhibition of antioxidant proteins. Among the proteins identified, some originated from the human erythrocyte host and correspond to host proteins imported by the parasite or proteins arising from host–parasite interaction. Consequently, cytosolic catalase and carbonic anhydrase I were down-represented under SHAM pressure. While P. falciparum lacks the antioxidant enzyme catalase, host catalase was imported into the food vacuoleCitation39,Citation50 and accomplished hydrogen peroxide detoxification.Citation51 Thus, the down-importation of catalase could be attributed to a SHAM effect against oxidative damage by diminishing hydrogen peroxide production. Carbonic anhydrase has been identified as a major intracellular peroxidation target in erythrocytes and could be attributed to oxidative stress induced by SHAM in the parasitized erythrocytes.Citation52
In our study, a number of identified proteins (enolase, catalase, glyceraldehyde-3-phosphate dehydrogenase, carbonic anhydrase) have been previously found as potential glutaredoxin target proteins.Citation53 In higher plants, the reduction of AOX in its active form involved a glutathione/thioredoxin coupling system, suggesting a SHAM effect on feedback regulation of AOX activity as described in higher plants.Citation24,Citation46
The other deregulated proteins were particularly involved in protein processing (chaperonin-containing T-complex 1), as well as translation (elongation factor 2), consequences of a general stress state of the parasite. Chaperone-assisted protein folding was described to contribute to protein folding, assembly, and translocation, inducing expression of proteins in these pathways under several types of cellular stress.Citation54
Conclusion
Although the AOX gene was not characterized yet in the P. falciparum, SHAM, an AOX specific inhibitor, disturbed the in vitro growth of P. falciparum in hyperoxic conditions. Additionally, recent studies sustained the existence of the alternative respiratory pathway in P. falciparum, illustrated by the identification of alternative NADH dehydrogenase, mostly described in some detail for plants.Citation13,Citation55 Despite the fact that the P. falciparum AOX gene was not yet identified, the metabolic pathways involved in SHAM response illustrates the existence of a similar AOX function. The failure of a comparative sequence analysis is mainly due to the extreme adenine-thymine bias (80%) of the parasite genome or the high gene variability between other species and P. falciparum.Citation56 Nevertheless, an AOX-like function could play a role in oxidative stress defense and could be a major benefit to P. falciparum, which appears to be sensitive to hyperoxic conditions. Indeed, the respiratory metabolism was the predominant source of ROS on P. falciparum, and partial inhibition of this metabolism led to a decrease of ROS. This hypothesis is in agreement with a publication reporting the AOX protective role in preventing ROS production.Citation23
Several studies on the use of mETC inhibitors underlined the crucial role of respiratory metabolism to parasite survival.Citation9 However, much work remains to scrutinize the specific biochemical pathways of the P. falciparum mitochondria.Citation8 This unique particularity of the alternative respiratory pathway could be a promising target for the development of a new group of antimalarial drugs.
Acknowledgments
The authors acknowledge the financial support of Délégation Générale pour l’Armement and the Direction Centrale du Service de Santé des Armées (grant no. 06co008). This paper has been reviewed and corrected by a journal expert.
Disclosure
The authors report no conflicts of interest in this work.
References
- FryMBeesleyJEMitochondria of mammalian Plasmodium sppParasitology1991102 Pt 117262038500
- Mi-IchiFTakeoSTakashimaEUnique properties of respiratory chain in Plasmodium falciparum mitochondriaAdv Exp Med Biol200353111713312916785
- RalphSAStrange organelles–Plasmodium mitochondria lack a pyruvate dehydrogenase complexMol Microbiol20055511415612911
- Van DoorenGGStimmlerLMMcFaddenGIMetabolic maps and functions of the Plasmodium mitochondrionFEMS Microbiol Rev200630459663016774588
- FryMWebbEPudneyMEffect of mitochondrial inhibitors on adenosinetriphosphate levels in Plasmodium falciparumComp Biochem Physiol B19909647757822171868
- PainterHJMorriseyJMMatherMWVaidyaABSpecific role of mitochondrial electron transport in blood-stage Plasmodium falciparumNature20074467131889117330044
- MogiTKitaKIdentification of mitochondrial complex II subunits SDH3 and SDH4 and ATP synthase subunits a and b in Plasmodium sppMitochondrion20099644345319682605
- Torrentino-MadametMDesplansJTravailléCJammesYParzyDMicroaerophilic respiratory metabolism of Plasmodium falciparum mitochondrion as a drug targetCurr Mol Med2010101294620205678
- VaidyaABMitochondrial and plastid functions as antimalarial drug targetsCurr Drug Targets Infect Disord200441112315032631
- MatherMWHenryKWVaidyaABMitochondrial drug targets in apicomplexan parasitesCurr Drug Targets200781496017266530
- Torrentino-MadametMAlmérasLDesplansJGlobal response of Plasmodium falciparum to hyperoxia: a combined transcriptomic and proteomic approachMalar J201110421223545
- UyemuraSALuoSVieiraMMorenoSNDocampoROxidative phosphorylation and rotenone-insensitive malate- and NADH-quinone oxidoreductases in Plasmodium yoelii yoelii mitochondria in situJ Biol Chem2004279138539314561763
- BiaginiGAViriyavejakulPO’NeillPMBrayPGWardSAFunctional characterization and target validation of alternative complex I of Plasmodium falciparum mitochondriaAntimicrob Agents Chemother20065051841185116641458
- DongCKPatelVYangJCType II NADH dehydrogenase of the respiratory chain of Plasmodium falciparum and its inhibitorsBioorg Med Chem Lett200919397297519097788
- MartinWMullerMThe hydrogen hypothesis for the first eukaryoteNature1998392667137419510246
- Lopez-GarciaPMoreiraDMetabolic symbiosis at the origin of eukaryotesTrends Biochem Sci1999243889310203753
- Van der GiezenMTovarJDegenerate mitochondriaEMBO Rep20056652553015940286
- SmilksteinMJForquerIKanazawaAA drug-selected Plasmodium falciparum lacking the need for conventional electron transportMol Biochem Parasitol20081591646818308406
- SrivastavaIKRottenbergHVaidyaABAtovaquone, a broad spectrum antiparasitic drug, collapses mitochondrial membrane potential in a malarial parasiteJ Biol Chem19972727396139669020100
- LegrandEDemarMVolneyBFirst case of emergence of atovaquone resistance in Plasmodium falciparum during second-line atovaquoneproguanil treatment in South AmericaAntimicrob Agents Chemother20075162280228117438062
- MurphyADLang-UnnaschNAlternative oxidase inhibitors potentiate the activity of atovaquone against Plasmodium falciparumAntimicrob Agents Chemother199943365165410049282
- SchonbaumGRBonnerWDJrStoreyBTBahrJTSpecific inhibition of the cyanide-insensitive respiratory pathway in plant mitochondria by hydroxamic acidsPlant Physiol19714711241285543780
- JuszczukIMRychterAMAlternative oxidase in higher plantsActa Biochim Pol20035041257127114740012
- VanlerbergheGCMcIntoshLALTERNATIVE OXIDASE: from gene to functionAnnu Rev Plant Physiol Plant Mol Biol19974870373415012279
- DivoAAGearyTGJensenJBGinsburgHThe mitochondrion of Plasmodium falciparum visualized by rhodamine 123 fluorescenceJ Protozool19853234424463900366
- ChaudhuriMOttRDHillGCTrypanosome alternative oxidase: from molecule to functionTrends Parasitol2006221048449116920028
- Bar NunNPlakhineDJoelDMMayerAMChanges in the activity of the alternative oxidase in Orobanche seeds during conditioning and their possible physiological functionPhytochemistry200364123524112946422
- RobertsCWRobertsFHenriquezFLEvidence for mitochondrial-derived alternative oxidase in the apicomplexan parasite Cryptosporidium parvum: a potential anti-microbial agent targetInt J Parasitol200434329730815003491
- SuzukiTHashimotoTYabuYAlternative oxidase (AOX) genes of African trypanosomes: phylogeny and evolution of AOX and plastid terminal oxidase familiesJ Eukaryot Microbiol200552437438116014016
- SuzukiTNiheiCYabuYMolecular cloning and characterization of Trypanosoma vivax alternative oxidase (AOX) gene, a target of the trypanocide ascofuranoneParasitol Int200453323524515468531
- BriolantSAlmerasLBelghaziMPlasmodium falciparum proteome changes in response to doxycycline treatmentMalar J2010914120500856
- DesjardinsRECanfieldCJHaynesJDChulayJDQuantitative assessment of antimalarial activity in vitro by a semiautomated microdilution techniqueAntimicrob Agents Chemother1979166710718394674
- TragerWJensenJBHuman malaria parasites in continuous culture. 1976J Parasitol200591348448616108535
- LambrosCVanderbergJPSynchronization of Plasmodium falciparum erythrocytic stages in cultureJ Parasitol1979653418420383936
- UhlemannACStaalsoeTKlinkertMQHviidLAnalysis of Plasmodium falciparum-infected red blood cellsMACS & more20004278
- PastorinoBBoucomont-ChapeaublancEPeyrefitteCNIdentification of cellular proteome modifications in response to West Nile virus infectionMol Cell Proteomics2009871623163719395707
- ShevchenkoAWilmMVormOMannMMass spectrometric sequencing of proteins silver-stained polyacrylamide gelsAnal Chem19966858508588779443
- MurphyADDoellerJEHearnBLang-UnnaschNPlasmodium falciparum: cyanide-resistant oxygen consumptionExp Parasitol19978721121209326886
- KoncarevicSRohrbachPDeponteMThe malarial parasite Plasmodium falciparum imports the human protein peroxiredoxin 2 for peroxide detoxificationProc Natl Acad Sci U S A200910632133231332819666612
- KrungkraiJBuratDKudanSKrungkraiSPrapunwattanaPMitochondrial oxygen consumption in asexual and sexual blood stages of the human malarial parasite, Plasmodium falciparumSoutheast Asian J Trop Med Public Health199930463664210928353
- HelfertSEstevezAMBakkerBMichelsPClaytonCRoles of triosephosphate isomerase and aerobic metabolism in Trypanosoma bruceiBiochem J2001357Pt 111712511415442
- OliveiraPLOliveiraMFVampires, Pasteur and reactive oxygen species. Is the switch from aerobic to anaerobic metabolism a preventive antioxidant defence in blood-feeding parasites?FEBS Lett20025251–33612163151
- RasmussonAGGeislerDAMollerIMThe multiplicity of dehydrogenases in the electron transport chain of plant mitochondriaMitochondrion200881476018033742
- HenriquezFLRichardsTARobertsFMcLeodRRobertsCWThe unusual mitochondrial compartment of Cryptosporidium parvumTrends Parasitol2005212687415664529
- RhoadsDMUmbachALSweetCRLennonAMRauchGSSiedowJNRegulation of the cyanide-resistant alternative oxidase of plant mitochondria. Identification of the cysteine residue involved in alpha-keto acid stimulation and intersubunit disulfide bond formationJ Biol Chem19982734630750307569804851
- Arnholdt-SchmittBCostaJHde MeloDFAOX – a functional marker for efficient cell reprogramming under stress?Trends Plant Sci200611628128716713324
- LianLYAl-HelalMRoslainiAMGlycerol: an unexpected major metabolite of energy metabolism by the human malaria parasiteMalar J200983819267910
- VaidyaABMatherMWMitochondrial evolution and functions in malaria parasitesAnnu Rev Microbiol20096324926719575561
- SluseFEJarmuszkiewiczWAlternative oxidase in the branched mitochondrial respiratory network: an overview on structure, function, regulation, and roleBraz J Med Biol Res19983167337479698817
- Ginsburg,HagaiMalaria parasite metabolic pathways1112000 Available from: http://sites.huji.ac.il/malariaAccessed July 28, 2011
- ClareboutGSlomiannyCDelcourtPStatus of Plasmodium falciparum towards catalaseBr J Haematol1998103152599792289
- WiwanitkitVPlasmodium and host carbonic anhydrase: molecular function and biological processGene Ther Mol Biol2006102251254
- SturmNJortzikEMailuBMIdentification of proteins targeted by the thioredoxin superfamily in Plasmodium falciparumPLoS Pathog200954e100038319360125
- AcharyaPKumarRTatuUChaperoning a cellular upheaval in malaria: heat shock proteins in Plasmodium falciparumMol Biochem Parasitol20071532859417307260
- FisherNBrayPGWardSABiaginiGAThe malaria parasite type II NADH: quinone oxidoreductase: an alternative enzyme for an alternative lifestyleTrends Parasitol200723730531017499024
- PailaUKondamRRanjanAGenome bias influences amino acid choices: analysis of amino acid substitution and re-compilation of substitution matrices exclusive to an AT-biased genomeNucleic Acids Res200836216664667518948281