Abstract
Aims
A substitution of diacylglycerol (DAG) oil for triacylglycerol (TAG) oil in diet has been reported to reduce body fat and body weight, possibly by increasing postprandial energy expenditure (EE). We have previously studied plasma serotonin, which increases EE and exists in the small intestine, in individuals who ingested TAG and DAG oil, and found that DAG ingestion elevates plasma serotonin levels by about 50% compared with TAG ingestion. We studied the molecular mechanisms for DAG-mediated increase in serotonin and EE.
Methods
We studied effects of 1-monoacylglycerol and 2-monoacylglycerol, distinct digestive products of DAG and TAG, respectively, on serotonin release from the Caco-2 cells (the human intestinal cell line, n = 8). Further, we studied effects of 1- and 2-monoacylglycerol, and serotonin on expression of mRNA associated with β-oxidation, FA metabolism, and thermogenesis, in the Caco-2 cells (n = 5).
Results
1-monoacylglycerol (100 μM 1-monooleyl glycerol [1-MOG]) significantly increased serotonin release from the Caco-2 cells compared with 2-monoacylglycerol (100 μM 2-MOG) by 36.6%. Expression of mRNA of acyl-CoA oxidase (ACO), fatty acid translocase (FAT), and uncoupling protein-2 (UCP-2) were significantly higher in 100 μM 1-MOG-treated Caco-2 cells than 100 μM 2-MOG-treated cells by 12.8%, 23.7%, and 35.1%, respectively. Further, expression of mRNA of ACO, medium-chain acyl-CoA dehydrogenase, FAT, and UCP-2 were significantly elevated in serotonin (400 nM)-treated Caco-2 cells compared with cells incubated without serotonin by 28.7%, 30.1%, and 39.2%, respectively.
Conclusions
Our study demonstrated that 1-monoacylglycerol, a digestive product of DAG, increases serotonin release from the Caco-2 cells, and enhances expression of genes associated with β-oxidation, FA metabolism, and thermogenesis, and that serotonin increases expression of these genes, proposing a novel molecular mechanism for DAG-mediated promotion of negative caloric balance.
Introduction
Diacylglycerol (DAG) is a natural component of various edible oils. DAG has been reported to be more effective for preventing excess adiposity through a variety of potential mechanisms in comparison with triacylglycerol (TAG) in humans.Citation1 In a study that examined the effects of DAG-rich diet on postprandial energy expenditure (EE) and respiratory quotient (RQ), the amount of changed EE after DAG ingestion was higher than that after TAG ingestion, and RQ after DAG ingestion was significantly lower than that after TAG ingestion, suggesting that DAG increases EE by inducing higher lipid oxidation compared with TAG.Citation2 We have previously studied plasma serotonin, which increases EE and body temperature, in subjects who ingested TAG and DAG.Citation3 In that study, we found that DAG oil ingestion elevates plasma serotonin levels by about 50%, compared with TAG oil ingestion, suggesting a possible mechanism for postprandial increase in EE by DAG.
Dietary TAG oil is hydrolyzed by lipase to free fatty acids (FFA) and 2-monoacylglycerol in the intestinal lumen, and these metabolites are absorbed by intestinal cells.Citation1 In the case of DAG oil, the metabolic pathway in the intestinal cells is different from that of TAG oil. 1,3-DAG, the main form of DAG oil, would be hydrolyzed to 1-monoacylglycerol and FFA.Citation1 To understand how DAG ingestion increases plasma serotonin levels as compared with TAG ingestion, we studied effects of 1-monoacylglycerol and 2-monoacylglycerol, hydrolytic products of DAG and TAG, respectively, on serotonin release from the intestinal cells, the Caco-2 cells. Further, we studied effects of serotonin, 1- and 2-monoacylglycerol on expression of mRNA of acyl-CoA oxidase (ACO), medium-chain acyl-CoA dehydrogenase (MCAD), fatty acid translocase (FAT), and uncoupling protein-2 (UCP-2), which are associated with β-oxidation, FA metabolism, and thermogenesis.
Materials and methods
Cells
Caco-2 cells were obtained from ATCC (American Type Culture Collection, Manassas, VA). Caco-2 cells were grown in 75-cm2 plastic flasks at 37 °C in a 5% CO2 95% air environment. The culture medium consisted of Dulbecco’s modified Eagle’s medium (DMEM), 20% fetal bovine serum (FBS), 100 IU/ml penicillin, and 100 μg/ml streptomycin. Cells reached confluence after 4–5 days in culture.
Effects of 1-monooleyl glycerol (1-MOG) and 2-MOG on serotonin release and expression of mRNA of ACO, MCAD, FAT, and UCP-2 in the Caco-2 cells
1-MOG and 2-MOG were purchased from Doosan Serdary Research Laboratories (Toronto, ON, Canada), and were dissolved and dispersed in 10 mmol/l taurocholate in phosphate-buffered saline (PBS), pH 7.4, to obtain the desired concentration, and were further incubated for one hour at 37 °C with shaking at 90 rpm to obtain a homogeneous solution. The stable monomer concentrations for oleic acid and 2-monoacylglycerol have been reported to be <200 μM and <125 μM, respectively.Citation4 The Caco-2 cells were then incubated with medium containing 100 μM and 200 μM 1-MOG or 2-MOG at 37 °C for 24 hours, and serotonin concentrations in the medium were measured by a previously reported method.Citation5 Briefly, serotonin was separated by high-performance liquid chromatography with column-switching system, and was specifically converted into a fluorescent derivative with benzylamine for convenient detection.Citation5
Total RNA was extracted from the Caco-2 cells, and cDNA was synthesized using 1 μg of total RNA by reverse transcription after treatment with DNase. Quantitative polymerase chain reaction (PCR) was done using the 7500 Real-Time PCR system and the TaqMan Gene Expression Assay system (Applied Biosystems, Foster City, CA). Gene expression analyses of ACO, MCAD, FAT, and UCP-2 were performed by relative quantitation (RQ) assay using 18S as an internal control. The Taqman Gene Expression Assays Hs01074241, Hs00936576, Hs01567186, Hs01075227, and Hs99999901_S1 were used for ACO, MCAD, FAT, UCP-2, and 18S, respectively.
Effects of serotonin on expression of mRNA of ACO, MCAD, FAT, and UCP-2
Caco-2 cells were incubated with medium without and with 50 nM and 400 nM serotonin at 37 °C for 24 hours. Total RNA was extracted from the Caco-2 cells, and cDNA was synthesized using 1 μg of total RNA by reverse transcription after treatment with DNase. Quantitative PCR was done using the 7500 Real-Time PCR system and the TaqMan Gene Expression Assay system (Applied Biosystems). Gene expression analyses of ACO, MCAD, FAT, and UCP-2 were performed by Relative RQ assay using 18S as an internal control. The Taqman Gene Expression Assays Hs01074241, Hs00936576, Hs01567186, Hs01075227, and Hs99999901_S1 were used for ACO, MCAD, FAT, UCP-2, and 18S, respectively.
Statistical analysis
Values were expressed as mean ± standard deviation (SD). Comparison of serotonin concentration in the medium of Caco-2 cells treated with 1-MOG and 2-MOG was analyzed using the Mann–Whitney U test. Comparison of mRNA expressions of ACO, MCAD, FAT, and UCP-2 in the Caco-2 cells incubated with 1-MOG and 2-MOG was analyzed using the Mann–Whitney U test. Comparisons in mRNA expressions of ACO, MCAD, FAT, and UCP-2 in the Caco-2 cells incubated with different concentrations of serotonin, were analyzed by one-factor analysis of variance (ANOVA) and Scheffe’s F test.
Results
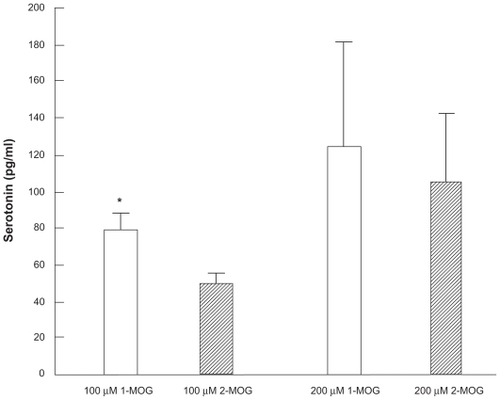
100 μM 1-MOG significantly increased serotonin release from the Caco-2 cells compared with 100 μM 2-MOG by 36.6% (P < 0.001, Mann–Whitney U test) (). Although a statistical significant difference was not obtained, increased serotonin release was observed in the Caco-2 cells treated with 200 μM 1-MOG as compared with cells treated with 200 μM 2-MOG.
Figure 1 Effects of 1-monooleyl glycerol (1-MOG) and 2-monooleyl glycerol (2-MOG) on serotonin release from the Caco-2 cells.
Expression of mRNA of ACO, FAT, and UCP-2 were significantly higher in 100 μM 1-MOG-treated Caco-2 cells than 100 μM 2-MOG-treated cells by 12.8% (P < 0.01), 23.7% (P < 0.01), and 35.1% (P < 0.01), respectively ().
Figure 2 Effects of 100 μM 1-monooleyl glycerol (1-MOG) and 2-monooley glycerol (2-MOG) on expression of mRNA of acyl-CoA oxidase (ACO) A) medium-chain acyl-CoA dehydrogenase (MCAD) B) fatty acid translocase (FAT) C) and uncoupling protein-2 (UCP-2) D) in the Caco-2 cells.
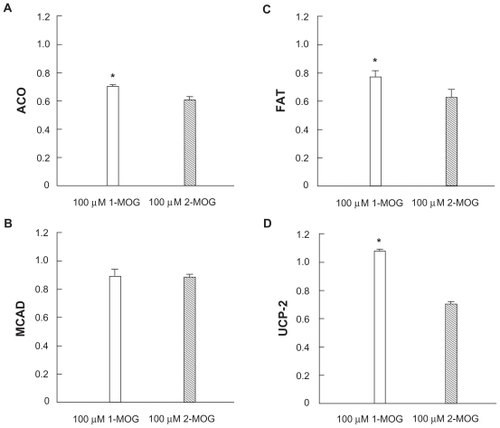
Figure 3 Effects of serotonin on expression of mRNA of acyl-CoA oxidase (ACO) A) medium-chain acyl-CoA dehydrogenase (MCAD) B) fatty acid translocase (FAT) C) and uncoupling protein-2 (UCP-2) D) in the Caco-2 cells.
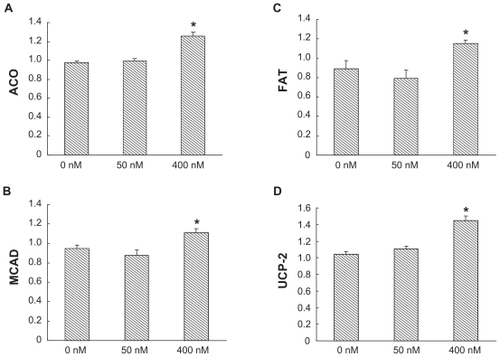
Further, expression of mRNA of ACO, MCAD, FAT, and UCP-2 were significantly elevated in serotonin (400 nM)-treated Caco-2 cells compared with cells incubated without serotonin by 28.7% (P < 0.01), 30.1% (P < 0.01), and 39.2% (P < 0.01), respectively.
Discussion
Upregulated mRNA expression associated with mitochondrial and peroxisomal β-oxidation and thermogenesis in the small intestine may explain in part mechanism for DAG-mediated increase in postprandial EE.Citation6,Citation7 The small intestine is likely to be the key organ to make differences in energy metabolism between DAG and TAG. Serotonin is present in great quantity in the small intestine, and is associated with increase in oxygen consumption and body temperature.Citation8,Citation9 An administration of a serotonin receptor agonist significantly increased body temperature and EE in Zucker rats in a dose-dependent manner, and a selective serotonin receptor antagonist completely inhibited this increase in EE, suggesting that serotonin regulates thermogenesis and EE.Citation10 A single injection of serotonin in rats stimulated resting oxygen consumption; this was reduced by pretreatment with automatic nerve ganglion blocking agent, indicating that serotonin mediates peripheral sympathetic thermogenesis.Citation11 We have previously studied plasma serotonin in subjects who ingested TAG and DAG, and found that DAG oil ingestion elevates plasma serotonin levels by about 50% compared with TAG oil ingestion, suggesting a possible mechanism for postprandial increase in EE by DAG.Citation3
To reveal further molecular mechanisms underlying this effect of DAG, we performed this study. We demonstrated that 1-MOG, a digestive product of DAG, significantly increased serotonin release, and also enhanced expression of mRNA of ACO, FAT, and UCP-2, compared with 2-MOG, a digestive product of TAG, in the Caco-2 cells. Further, our study showed that serotonin upregulates expression of mRNA associated with β-oxidation, FA metabolism, and thermogenesis. These results suggest that a digestive product of DAG increases serotonin release, and serotonin induces expression of genes associated with elevation in energy expenditure, and consequently leading to DAG-mediated promotion of negative caloric balance. To our knowledge, the present study is the first to report serotonin-mediated elevation in expression of genes associated with β-oxidation, FA metabolism, and thermogenesis. However, it remains unclear how much 1- or 2-monoacylglycerol exists in the intestine when we ingested DAG or TAG oil. Therefore, we exactly don’t know whether or not the concentrations used in our studies are similar to those expected in a natural biological system or simply artificial. To make our preliminary results valid and meaningful, we should investigate the concentration of metabolites in the intestine after TAG or DAG oil ingestion, and should also study a dose-dependent and a time-dependent effect of serotonin and 1-, 2-monoacylglycerol for expression of genes associated with β-oxidation, FA metabolism, and thermogenesis.
More than 90% of total body content of serotonin is synthesized, stored and released by intestinal enterochromaffin cells. Activation of the various receptors such as cholinergic, nicotinic, and serotonin 3 receptors, trigger intracellular messenger systems, which regulate the secretion of serotonin.Citation12 Monoacylglycerol has been reported to act as an endogenous ligand for the cannabinoid receptors,Citation13 which seem to be involved in serotonin 3 receptors.Citation14 Although we should perform more studies to elucidate the underlying mechanism in the future, 1-monoacylglycerol may be more strongly associated with stimulatory receptors for serotonin release from the enterochromaffin cells compared with 2-monoacylglycerol.
Serotonin has been reported to be associated with vasoconstriction and platelet function.Citation15 In our previous study,Citation3 we studied plasma von-Willebrand factor, thrombomodulin, and p-selection in individuals who ingested TAG and DAG, and did not observe a significant difference between TAG and DAG ingestion (data not shown). A significance of DAG-mediated increase in plasma serotonin for atherosclerosis remains unclear, which should be elucidated.
In conclusion, our study demonstrated that a hydrolytic product of DAG increases serotonin release from the Caco-2 cells, and enhances expression of genes associated with β-oxidation, thermogenesis, and FA metabolism, which were also upregulated by serotonin, proposing a new molecular biological mechanism for DAG-induced promotion of negative caloric balance.
Disclosures
The authors report no conflicts of interest in this work.
References
- Yanai H Tomono Y Ito K Diacylglycerol oil for the metabolic syndrome Nutr J 2007 6 43 18072966
- Saito S Tomonobu K Hase T Effects of diacylglycerol on postprandial energy expenditure and respiratory quotient in healthy subjects Nutrition 2006 22 30 35 16289979
- Yanai H Yoshida H Tomono Y Effects of diacylglycerol on glucose, lipid metabolism, and plasma serotonin levels in lean Japanese Obesity (Silver Spring) 2008 16 47 51 18223611
- Murota K Storch J Uptake of micellar long-chain fatty acid and sn-2-monoacylglycerol into human intestinal Caco-2 cells exhibits characteristics of protein-mediated transport J Nutr 2005 135 1626 1630 15987840
- Hirowatari Y Hara K Kamihata H High-performance liquid chromatographic method with column-switching and post-column reaction for determination of serotonin levels in platelet-poor plasma Clin Biochem 2004 37 191 197 14972640
- Murase T Nagasawa A Suzuki J Dietary alpha-linolenic acid-rich diacylglycerols reduce body weight gain accompanying the stimulation of intestinal beta-oxidation and related gene expressions in C57BL/KsJ-db/db mice J Nutr 2002 132 3018 3022 12368389
- Murase T Aoki M Wakisaka T Anti-obesity effect of dietary diacylglycerol in C57BL/6J mice: dietary diacylglycerol stimulates intestinal lipid metabolism J Lipid Res 2002 43 1312 1319 12177175
- Resnick RH Gray SJ Distribution of serotonin (5-hydroxytryptamine) in the human gastrointestinal tract Gastroenterology 1961 41 119 121 13740943
- Mazzola-Pomietto P Aulakh CS Wozniak KM Evidence that m-chlorophenylpiperazine-induced hyperthermia in rats is mediated by stimulation of 5-HT2C receptors Psychopharmacology (Berl) 1996 123 333 339 8867872
- Hayashi A Sonoda R Kimura Y Antiobesity effect of YM348, a novel 5-HT2C receptor agonist, in Zucker rats Brain Res 2004 1011 221 227 15157808
- Rothwell NJ Stock MJ Effect of diet and fenfluramine on thermogenesis in the rat: possible involvement of serotonergic mechanisms Int J Obesi 1987 11 319 324
- Hansen MB Witte AB The role of serotonin in intestinal luminal sensing and secretion Acta Physiol (Oxf) 2008 193 311 323 18462271
- Dinh TP Carpenter D Leslie FM Brain monoglyceride lipase participating in endocannabinoid inactivation Proc Natl Acad Sci U S A 2002 99 10819 10824 12136125
- Rácz I Bilkei-Gorzo A Markert A Anandamide effects on 5-HT(3) receptors in vivo Eur J Pharmacol 2008 596 98 101 18775693
- Mohammad-Zadeh LF Moses L Gwaltney-Brant SM Serotonin: a review J Vet Pharmacol Ther 2008 31 187 199 18471139