
Abstract
Subcellular distribution of the apoptosis inhibitor survivin and its ability to relocalize as a result of cell cycle phase or therapeutic insult has led to the hypothesis that these subcellular pools may coincide with different survivin functions. The PIK kinases (ATM, ATR and DNA-PK) phosphorylate a variety of effector substrates that propagate DNA damage signals, resulting in various biological outputs. Here we demonstrate that subcellular repartitioning of survivin in MCF-7 cells as a result of UV light-mediated DNA damage is dependent upon DNA damage-sensing proteins as treatment with the pan PIK kinase inhibitor wortmannin repartitioned survivin in the mitochondria and diminished it from the cytosol and nucleus. Mitochondrial redistribution of survivin, such as was recorded after wortmannin treatment, occurred in cells lacking any one of the three DNA damage sensing protein kinases: DNA-PK, ATM or ATR. However, failed survivin redistribution from the mitochondria in response to low-dose UV occurred only in the cells lacking ATM, implying that ATM may be the primary kinase involved in this process. Taken together, this data implicates survivian’s subcellular distribution is a dynamic physiological process that appears responsive to UV light-initiated DNA damage and that its distribution may be responsible for its multifunctionality.
Keywords:
Supplement
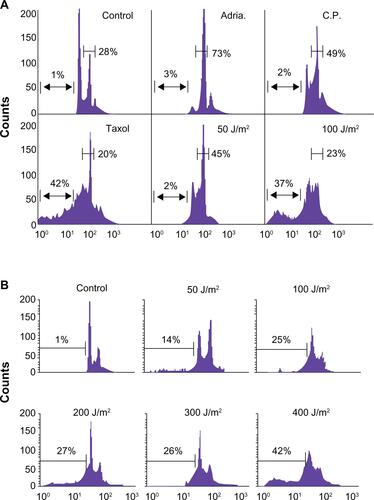
Supplemental Figure 1 (A) Low dose genotoxic stressors such as Adriamycin (Adria., 100 nM), Cisplatin (C.P., 3 mM) and UV (50 J/m2) arrest MCF-7 cells in the G2/M phase of the cell cycle while high-dose UV (doses ≥ 100 J/m2) or the tubulin poison Taxol (2 mM) induce cellular death. (B) Breast carcinoma MCF-7 cells were irradiated with 50–400 J/m2 UV under minimal 1X PBS. PBS was immediately removed and complete culture medium was added. After 24 hours, cells were harvested and analyzed for DNA content by propidium iodide staining and flow cytometry. Percentages of apoptotic cells with hypodiploid (sub-G1) DNA content are indicated per each condition tested. Data are representative of one of two independent experiments with comparable results.
Acknowledgements
The author’s would like to thank especially, Dr. William A. Cliby (Mayo Clinic, Rochester, MN USA) for the kind gift of the GM 847kd fibroblast cells. Grant Support: NCMHD Project EXPORT Program 5P20MD001632/Project 3 (N.R. Wall). Funding was also obtained as part of a start-up package from Loma Linda University’s Center for Molecular Biology and Gene Therapy, now the Center for Health Disparities Research and Molecular Medicine (NRW) and a National Merit Test Bed (NMTB) award sponsored by the Department of the Army under Cooperative Agreement Number DAMD17–97–2–7016 (NRW).
Disclosure
The authors declare that they have no competing interests.