Abstract
The purpose of this study was to determine whether reflex sweating during isometric handgrip exercise (IHG) in the heat was due to a priming effect in the sweat glands or an increase in skin temperature. Ten male subjects completed four trials where they performed IHG for three minutes at 40% of their maximal voluntary contraction (MVC). The four trials included: (1) a control trial in thermoneutral conditions (23 ± 1oC), (2) after sitting in hyperthermic conditions (35 ± 1oC) for 30 min, (3) a local heating trial after having their non-exercising arm wrapped in a heat pad that maintained forearm skin temperature at ∼35oC for 30 min, 4) and after pilocarpine iontophoresis to a 5 cm2 area of the forearm. The sweating rate (SR), as measured by resistance hygrometry, was not significantly different (P > 0.05) from baseline during IHG in either the control or local heating trial, but was significantly increased (P < 0.05) from baseline during the hyperthermic and pilocarpine trials. Baseline SR values of the hyperthermic and pilocarpine trials (∼0.25 mg/cm2/min) were significantly greater than the control and local heating trials (∼0.05 mg/cm2/min). These results suggest that reflex sweating in the heat during IHG is primarily due to a priming effect in the sweat glands and not because of an increase in skin temperature.
Introduction
The evaporation of sweat is the primary method that humans have developed to moderate their body temperature in hyperthermic environments and when exercising. The primary stimuli for sweat production are increases in core and skin temperature Citation1, Citation2. Isometric handgrip exercise (IHG) however, has been shown to increase sweat rate (SR), with the increase seemingly unrelated to thermal factors as neither skin nor core temperature changes significantly during acute IHG exercise Citation3–8.
Most studies have suggested that a reflex mechanism is responsible for the increase in SR seen with IHG as sweating occurs in the arm opposite the one performing the exercise Citation4, Citation6–8. It has been shown that various nonthermal factors, such as an increase in central command and metabo- and mechanoreceptor stimulation are capable of modulating SR and are responsible for the reflex sweating seen with IHG Citation5, Citation7, Citation9–11.
Interestingly, reflex sweating during IHG is not observable when performed in thermoneutral environments but rather seems to occur only under hyperthermic conditions. For example, Shibasaki et al. Citation7 had nine healthy subjects perform two minutes of IHG during hyperthermic and normothermic conditions. SR increased significantly by the second minute of IHG in the hyperthermic trial, but did not increase during the same exercise under thermoneutral conditions. It seems unlikely that alterations in known thermal and nonthermal stimuli could explain such results. Alternately, two potential mechanisms could be hypothesized to explain the findings. First, during the hyperthermic condition the subjects were already engaged in sweating prior to the initiation of IHG. In other words, the sweat glands were already primed (i.e. already sweating) and thus could more rapidly respond to the IHG in the heat. Second, the skin temperature was elevated during the hyperthermic condition, thus potentially improving sweat gland sensitivity by causing an increase in the amount of neurotransmitter released at the sweat gland, increasing receptor sensitivity, or lowering the sweating threshold Citation5, Citation6, Citation12, Citation13. In light of the above, the purpose of the current study was to determine whether reflex sweating during IHG in the heat was due to a priming effect in the sweat glands or an increase in skin temperature. To do this the current study, unlike past studies, used four separate SR trials in which local skin temperature(via local heating) and sweat gland priming (via pilocarpine iontophoresis) were independent controlled prior to the initiation of IHG exercise in the heat. It was hypothesized that both local skin temperature and sweat gland priming are both involved with reflex sweating during IHG exercise in the heat.
Methods
Subjects
The experiments were performed on 10 healthy males aged 24–54 y. They had a mean ± SD height and weight of 1.75 ± 0.04 m, and 79.85 ± 18.43 kg, respectively. Signed informed consent was obtained for all subjects prior to the start of data collection and approval for the study was given by the IRB of San Diego State University.
Procedures
Subjects completed four trials, with a minimum of 48 hours between each. Subjects performed all four trials at approximately the same time of day to minimize circadian variation Citation3, Citation4. Subjects were advised to drink at least 0.5 L of water one hour before the start of each trial and to refrain from exercise for at least three hours. To ensure subjects were adequately hydrated, they self-collected a urine sample and specific gravity was measured. Subjects were only allowed to perform the trial if their urinary specific gravity was <1.020. Each subject performed three maximal contractions on a Smedley handgrip dynamometer before their first trial, with the highest result being used to calculate 40% of their maximal voluntary contraction (MVC). During IHG, a marker was placed on the hand dynamometer to indicate 40% of the subject's MVC, and the investigator carefully watched to ensure that the appropriate force was maintained during each trial. Before each trial, core body temperature was recorded using an electronic oral thermometer and forearm skin temperature was recorded using a skin thermister (YSI series 400). Sweat rate was measured using the ventilated capsule method according to the procedure outlined by Bullard Citation14. The sweat capsule and skin thermister were attached on the flexor surface of the non-exercising forearm, half way between the elbow and wrist. During each trial baseline values were recorded for two minutes prior to the start of exercise. The subjects then began three minutes of IHG performed at 40% of MVC. The 40% MVC intensity is consistent with past studies Citation7, Citation8. Two minutes of post-exercise data were also recorded. SR data before, during and after IHG were recorded every 20 seconds. Immediately following exercise, skin and core temperature were again recorded.
The control trial was conducted in thermoneutral conditions (23 ± 1°C, 50 ± 3% relative humidity). The hyperthermic trial was conducted in an environmental chamber set at 35 ± 1°C and 40 ± 3% relative humidity. Subjects sat in the chamber for 30 minutes prior to performing IHG. The local heating trial was conducted in thermoneutral conditions with the subject's non-exercising forearm wrapped in an electric heating pad, which increased the skin temperature to 35.9 ± 0.7oC for thirty minutes prior to IHG. The purpose of the local heating trial was to raise the forearm skin temperature to the level seen during the hyperthermic trial (35.0 ± 0.6oC). It has previously been shown that localized skin heating to below 38oC does not induce sweating directly from the eccrine glands Citation13. The final trial involved stimulating a 5 cm2 area of skin on the subject's forearm to sweat using pilocarpine iontophoresis. A 1% solution of pilocarpine nitrate diluted in deionized water was iontophoresed (1.5 mA) for one minute, at the site where the ventilated capsule was to be placed. Baseline SR was allowed to stabilize (∼10–20 min) before the subject performed IHG. The purpose of this trial was to pharmacologically prime the sweat glands and initiate sweating prior to IHG in a thermoneutral setting. The order of the four trials was randomized for all subjects.
One-way repeated measures ANOVAs were performed to determine whether there were significant differences in the SR during each trial and between similar time points across trials. Post-hoc comparisons were performed using Tukey HSD analysis. The alpha level was set at P < 0.05.
Results
As can be seen in , IHG did not significantly increase the SR from baseline values during either the control trial or local heating trial. Such data suggests that elevated local skin temperature is not the cause for the previously reported increase in SR seen during IHG exercise in the heat. Conversely, during both the hyperthermic and pilocarpine trials IHG significantly increased SR compared to the baseline values. However, no significant differences were found at any time point between the hyperthermic and pilocarpine trials at baseline or during the 3 minutes of IHG. In other words, priming the sweat gland via either heat exposure (i.e., hyperthermia) or pharmacologically using pilocarpine ionthohoresis both caused significant increases in SR during IHG exercise.
Figure 1. Mean ± SE sweat rate before, during, and after three minutes of isometric handgrip exercise during the four trials. (*indicates significantly different (P < 0.05) from baseline, ł indicates that the hyperthermic and pilocarpine trials are significantly different (P < 0.05) from the control and local heating trial at baseline, NS indicates that there were no significant differences (P > 0.05) between trials at any point). On the X axis, the pre-IHG exercise period was between −120 and 0 seconds. IHG exercise occurred from 0 to 180 seconds. The post-IHG exercise period was beween 180 and 300 seconds.
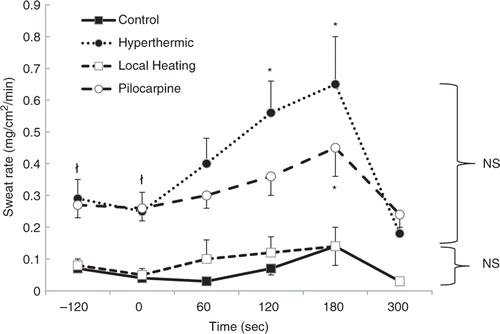
There was no significant difference in the baseline SR between the control and local heating trials. However, baseline SR during the hyperthermic and pilocarpine trials, which were essentially identical (∼0.25 mg/cm2/min), were significantly increased five-fold from the baseline SR seen in the control and local heating trials (∼0.05 mg/cm2/min). Thus, the pilocarpine iontophoresis procedure used in the current study was successful in matching the baseline SR with that seen during the hyperthermic trial. Therefore, an increase in SR during IHG during the pilocarpine trial could be attributed to sweat gland priming, rather than an increase in local skin temperature.
As expected, since the IHG exercise only lasted 3 min, there were no significant differences in core temperature across all four trials, both before and after IHG. Mean core temperatures ranged from 36.1 to 36.9oC during all recorded time points.
Discussion
The purpose of the current study was to determine whether reflex sweating during IHG in the heat was due to a priming effect in the sweat glands or an increase in skin temperature. The results agree with numerous previous studies which have reported that reflex sweating occurs when IHG exercise is performed in hyperthermic conditions, but is not observable in normothermic conditions Citation3–8. The most important new finding of the current study was that reflex sweating in the heat during IHG is primarily due to a priming effect in the sweat glands and not because of an increase in skin temperature. This conclusion is based on the fact that SR was not significantly different (P > 0.05) from baseline during IHG in either the control or local heating trial, but was significantly increased (P < 0.05) from baseline during the hyperthermic and pilocarpine trials. Furthermore, baseline SR values of the hyperthermic and pilocarpine trials were significantly greater than the control and local heating trials. These results indicate that when the subjects were already engaged in sweating prior to the initiation of IHG, as seen during the hyperthermic and pilocarpine trials, reflex sweat was observable. However, when skin temperature was increased during the local heating trial to the level seen during the hyperthermic trial, IHG exercise did not result in a reflex sweating response.
The magnitude of the increase in SR seen in the current study during hyperthermic IHG exercise is comparable to that reported by others Citation4, Citation7 in the past. Shibasaki et al. Citation7 found no increase in SR during IHG performed at 40% MVC during their control trial (22–23oC). However, SR increased by approximately 50% from baseline (from ∼0.40 to ∼0.60 mg/cm2/min) after two minutes of IHG during whole body heating. Similarly, Aoki et al. Citation4 had subjects perform one minute of IHG at 50% MVC during which time SR tripled from baseline values (∼0.1 to 0.3 mg/cm2/min). Such results agree with the findings of the current study which found that after three minutes of IHG exercise performed at 40% MVC in the heat, SR had more than doubled from baseline values (∼0.3 to ∼0.65 mg/cm2/min). Conversely, Crandall et al. Citation15 only found a 17% increase in SR during IHG exercise in the heat (∼0.6 to ∼0.7 mg.cm2/min). However their subjects performed IHG at 30% MVC for three minutes, achieving much smaller increases in SR from baseline to three minutes of IHG). It seems reasonable to hypothesize that the differences in the magnitude are most likely due to differences in either the duration or intensity (i.e. % of MVC) of the IHG protocols, and the subsequent differing activation of the muscle metaboreflex Citation4, Citation5, Citation7.
Our finding that the priming of the sweat gland is more important than an increase in local skin temperature for causing an increase in SR during IHG exercise in the heat is consistent with the results of Shibasaki et al. Citation7. They measured forearm SR at a control site and at a site treated with neostigmine, an acetylcholinesterase inhibitor, during IHG in both normothermic and hyperthermic conditions. They did not observe an increase in SR at the untreated site during the normothermic trial; a finding that is consistent with the current study and past research Citation7. However, SR was significantly increased at the neostigmine treated site during IHG in normothermia. Interestingly, they reported that spontaneous sweating was observed at the neostigmine treated site prior to the initiation of IHG in many subjects. In other words, increases in SR were only observed in the sweat glands that were already engaged (i.e. primed) prior to the start of IHG exercise; namely the neostigmine treated site.
In the current study, priming, or engaging the sweat glands prior to the start of IHG exercise was accomplished via pilocarpine iontophoresis whereas Shibasaki et al. Citation7 used neostigmine infusion. Regardless of the method used to prime the sweat gland, taken together the results of the current study and those of previous studies strongly suggest that priming of the sweat gland is more important for increasing SR during IHG in the heat than an increase in local temperature.
In addition, Shibasaki et al. Citation7 reported that the magnitude of the increase in SR during IHG was similar in the control and neostigmine treated sites during whole body heating. Presumably there was an increase in the periglandular acetylcholine concentration at the treated site, a situation analogous to what had been suggested to occur during local heating. Such findings strongly suggest that once the sweat glands are already sweating, the addition of more acetylcholine, whether from neostigmine infusion or local heating, does not affect the increase in SR seen during IHG.
In conclusion, the results of the current study indicate that when the subjects were already engaged in sweating prior to the initiation of IHG, as seen during the hyperthermic and pilocarpine trials, reflex sweat was observable. However, when skin temperature was increased during the local heating trial to the level seen during the hyperthermic trial, IHG exercise did not result in a reflex sweating response. The results of the current study add to the literature by showing that reflex sweating in the heat during IHG is primarily due to a priming effect in the sweat glands and not because of an increase in local skin temperature.
Declaration of interest: The authors report no conflicts of interest. The authors alone are responsible for the content and writing of the paper.
References
- Donaldson GC, Keatinge WR, Saunders RD. Cardiovascular responses to heat stress and their adverse consequences in healthy and vulnerable human populations. Int J Hyperthermia 2003; 19: 225–235
- Shibasaki M, Wilson TE, Crandall CG. Neural control and mechanisms of eccrine sweating during heat stress and exercise. J Appl Physiol 2006; 100: 1692–1701
- Aoki K, Kondo N, Shimomura Y, Iwanaga K, Harada H, Katsuura T. Time-of-Day effect on nonthermal control of sweating response to maintained static exercise in humans. Eur J Appl Physiol 2002; 86: 388–393
- Aoki K, Kondo N, Shimomura Y, Yokoi M, Iwanaga K, Harada H, Katsuura T. Sweating responses during activation of the muscle metaboreflex in humans is altered by time of day. Acta Physiol Scand 2004; 180: 63–70
- Kondo N, Tominaga H, Shibasaki M, Aoki K, Koga S, Nishiyasu T. Modulation of the thermoregulatory sweating response to mild hyperthermia during activation of the muscle metaboreflex in humans. J Physiol 1999; 515: 591–598
- Kondo N, Tominaga H, Shibasaki M, Aoki K, Okada S, Nishiyasu T. Effects of exercise intensity on the sweating response to a sustained static exercise. J Appl Physiol 2000; 88: 1590–1596
- Shibasaki M, Kondo N, Crandall CG. Evidence for metaboreceptor stimulation of sweating in normothermic and heat stressed humans. J Physiol 2001; 534: 605–611
- Yanagimoto S, Aoki K, Horikawa M, Shibasaki M, Inoue I, Nishiyasu T, Kondo N. Sweating response in physically trained men to handgrip exercise in mildly hyperthermic conditions. Acta Physiol Scand 2002; 174: 31–39
- Crandall CG, Stephens DP, Johnson JM. Muscle metaboreceptor modulation of cutaneous active vasodilation. Med Sci Sports Exerc 1998; 30: 490–496
- Shibasaki M, Secher NH, Selmer C, Kondo N, Crandall CG. Central command is capable of modulating sweating from non-glabrous human skin. J Physiol 2003; 553: 999–1004
- Vissing SF, Hjortso EM. Central motor command activates sympathetic outflow to the cutaneous circulation in humans. J Physiol 1996; 492: 931–939
- MacIntyre BA, Bullard MR, Banerjee MR, Elizondo R. Mechanism of enhancement of eccrine sweating by localized heating. J Appl Physiol 1968; 25: 255–260
- Ogawa T. Local effect of skin temperature on threshold concentration of sudorific agents. J Appl Physiol 1970; 28: 18–22
- Bullard RW. Continuous recording of sweating rate by resistance hygrometry. J Appl Physiol 1962; 17: 735–737
- Crandall CG, Musick J, Hatch JP, Kellogg DL, Johnson JM. Cutaneous vascular and sudomotor responses to isometric exercise in humans. J Appl Physiol 1995; 79: 1946–1950