
Abstract
Background. The forkhead box M1 (FOXM1) transcription factor plays an important role in the metastases of many cancers. Down-regulation of FOXM1 by its inhibitor, thiostrepton, can inhibit the metastatic potential of some cancers; however, there are few studies regarding the functional significance of FOXM1 and thiostrepton in the metastases of nasopharyngeal carcinoma (NPC) and the underlying mechanism.
Methods. Expression of FOXM1 in NPC, normal nasopharyngeal tissues, a NPC cell line (C666-1), and a nasopharyngeal epithelial cell line (NP69) was investigated by immunohistochemical staining, qRT-PCR, and Western blot. The correlation between FOXM1 expression and the clinical characteristics of patients was analyzed. Moreover, the effects of thiostrepton on expression of FOXM1 in C666-1 and NP69 cells, and the invasion and migration ability of C666-1 cells were examined. The expressions of MMP-2, MMP-9, fascin-1, ezrin, and paxillin were determined after treatment with thiostrepton.
Results. FOXM1 was overexpressed in NPC and C666-1 cells compared with normal nasopharyngeal tissues and NP69 cells. Overexpression of FOXM1 was associated with lymph node metastasis and advanced tumor stage. Moreover, thiostrepton inhibited expression of FOXM1 in C666-1 cells in a dose-dependent manner, but had a minimal effect on NP69 cells. Thiostrepton inhibited the migration and invasion ability of C666-1 cells by down-regulating the expression of MMP-2, MMP-9, fascin-1, and paxillin.
Conclusions. Overexpression of FOXM1 is associated with metastases of NPC patients. Thiostrepton inhibits the metastatic ability of NPC cells by down-regulating the expression of FOXM1, MMP-2, MMP-9, fascin-1, and paxillin.
Introduction
Nasopharyngeal carcinoma (NPC), an Epstein–Barr virus-associated malignancy, is the most common head and neck cancer in China, with an incidence of 30–80/100,000 population/year in southern China (Citation1). NPC has high invasive and metastatic features, and ∼90% of patients show cervical lymph node metastasis at the time of initial diagnosis (Citation2). Radiotherapy with or without chemotherapy is the mainstream treatment approach for NPC patients; however, there are limitations in these therapeutic applications due to undesirable side effects, local recurrence, and metastasis (Citation3). Thus, challenges in NPC treatment still exist, and novel therapeutic targets and new approaches for NPC treatment are urgently needed (Citation4).
The forkhead box M1 (FOXM1), a member of the Fox transcription factor family, regulates a number of cell cycle key factors involved in the transition from the G1 to S phase, G2 to M phase progression, and transition to mitosis (Citation5-7). It has been shown that FOXM1 represents a promising therapeutic target based on up-regulation in numerous human malignancies (Citation8-13), while FOXM1 is suppressed in non-dividing cells (Citation14). It has been recently reported that FOXM1 is overexpressed in NPC and knockdown of FOXM1 expression diminishes NPC cell proliferation (Citation15). Indeed, the functional significance of FOXM1 in NPC metastases has not been defined.
The FOXM1 inhibitor, thiostrepton, is a natural product with antibiotic properties isolated from Streptomyces azureus and is known to interact directly with FOXM1 and inhibit the binding of FOXM1 to genomic target sites (Citation16). In addition, it has been shown that thiostrepton inhibits cell growth and induces apoptosis in a variety of human cancer cell lines by inhibiting FOXM1 expression (Citation17-19). There are few studies that have focused on the effects of FOXM1 and thiostrepton on metastases of NPC, and the correlation between FOXM1 and thiostrepton, and MMP-2, MMP-9, fascin-1, ezrin, and paxillin.
In the current study we determined the expression of FOXM1 in NPC, normal nasopharyngeal tissues, a NPC cell line, and a nasopharyngeal epithelial cell line. The relationship between the expression of FOXM1 and clinicopathologic factors, such as TNM and clinical stage, was analyzed.
Materials and methods
Patients and tissue samples
A total of 65 histologically confirmed NPC patients were consecutively recruited between 2005 and 2008 from The First Affiliated Hospital of Chongqing Medical University in China. Twenty non-cancerous nasopharyngeal samples were collected from patients with clinical symptoms suggestive of NPC, but ruled out by biopsy. None of the patients received radiotherapy or chemotherapy prior to biopsy. Prior patient consent and approval from the Institute Research Ethics Committee were obtained. Among 65 patients with NPC, 45 were males and 20 were females (age range 33–75 years; median age 39 years). The clinical stage and histopathologic classification were based on the World Health Organization (WHO) classification, as follows: T1, 9 (13.8%); T2, 22 (33.8%); T3, 23 (35.4%); T4, 11 (16.9%); N–, 7 (10.8%); N+, 58 (89.2%); stage I, 7 (10.8%); stage II, 9 (13.8%); stage III, 30 (46.2%), and stage IV, 19 (29.2%). Furthermore, 10 NPC and 10 non-cancerous nasopharyngeal tissue samples were included in the cohort for immunohistochemistry to evaluate the expression of FOXM1 RNA and protein.
Immunohistochemical staining
Immunohistochemical staining was performed on 4-µm paraffin tissue sections mounted on poly-lysine-coated slides and dried at 37°C overnight. After the slides were deparaffinized in xylene and rehydrated conventionally, the endogenous peroxidase was blocked with 3% hydrogen peroxide in methanol for 20 min. Each slide was incubated with normal goat serum for 20 min at room temperature. The sections were incubated with rabbit anti-FOXM1 polyclonal antibody (1:100 dilution, sc-502; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) overnight at 4°C. After washing with PBS, the sections were incubated for 30 min with a horseradish peroxidase-labeled goat anti-rabbit secondary antibody (Boster Biological Technology, Ltd, Wuhan, China). Chromogen 3,3-diaminobenzidine (Boster Biological Technology, Ltd) was used for 15 min to visualize immunolabeling, resulting in a brown precipitate. After washing, the sections were counterstained with hematoxylin. Positive and negative immunohistochemistry controls were routinely performed.
The immunostaining results were evaluated and scored independently by two pathologists without knowledge of the clinicopathologic outcomes of the patients. Immunostaining for FOXM1 was limited to the cytoplasm. Specimens with staining in >10% of the cells were considered positive. Immunohistochemical reactivity was graded on a scale of 0–3, according to the intensity of staining and percentage of immunopositive cells as follows: 0, no staining or <10% positive cells; 1, weak staining in >10% of tumor cells or moderate staining in 10%–40% of tumor cells; 2, moderate staining in >40% of tumor cells or strong staining in 10%–40% of tumor cells; and 3, strong staining in >40% of tumor cells (Citation20).
Cell lines and chemical compounds
The NPC cell line, C666-1 (carries EBV), was cultured in RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA) with 10% fetal bovine serum (Gibco, Grand Island, NY, USA), 100 µg/mL of streptomycin, and 100 U/mL of penicillin (Hyclone, Logan, UT, USA). The immortalized normal human nasopharyngeal epithelial cell line, NP69, was grown in defined-KSFM medium supplemented with epidermal growth factor (EGF) (Invitrogen). Cells were cultured under a 5.0% CO2 atmosphere. Thiostrepton (Sigma, St. Louis, MO, USA) was dissolved in dimethylsulfoxide (DMSO).
Quantitative real-time RT-PCR
Total RNA was extracted using Trizol reagent (Takara, Dalian, China) according to the manufacturer’s instructions. Contaminating genomic DNA was removed by incubation with DNase I (Takara). RNA purity and concentration were determined by spectrophotometry. PCR was carried out according to the standard protocol on a real-time PCR system with SYBR Green detection. After an initial incubation of the 10-µL reaction mixture for 1 min at 95°C, 39 cycles (95°C for 20 s and 58°C for 25 s) were performed for amplification. The specificity of amplification was confirmed by melting curve analysis. Each sample was tested in triplicate, and the results were normalized to the level of the GAPDH gene. The sequences for each set of primers were as follows: 5′-TGCAGCTAGGGATGTGAATCTTC-3′ (sense) and 5′-GGAGCCCAG TCCATCAGAACT-3′ (antisense) for FOXM1; 5′-CTTCCCTTTGGTCTTT CTGT-3′ (sense) and 5′-GGTTGTCTGAAGTCAC TGCACAGT-3′ (antisense) for MMP-2; 5′-CTCG GTAGGGA CATGCTAAGTAGAG-3′ (sense) and 5′-CCTCT GGAGGTTCGACGTGA-3′ (antisense) for MMP-9; 5′-TCATGAGGAAGAGCTCTGAG T-3′ (sense) and 5′-TCATGAGGAAGAGCTCTGA GT-3′ (antisense) for fascin-1; 5′-TGCCGAAACCAATCA ATG TC-3′ (sense) and 5′-GCTTCAGCCAGGTAGGAA ATC-3′ (antisense) for ezrin; 5′-CC AACAGCAG ACACGCATCT-3′ (sense) and 5′-GGTGAGCTG CTCCCTGTCTT-3′ (antisense) for paxillin; and 5′-CAGCGACACCCACTCCTC-3′ (sense) and 5′-TGAGGTCCACCACCCTGT-3′ (antisense) for GAPDH. The 2-ΔΔCT method was used to calculate the relative expressions.
Western blot
Protein samples were separated on precast 10% SDS polyacrylamide gels (SDS-PAGE). After electrophoresis, the proteins were transferred to PVDF membrane filters (Millipore Biotechnology, Billerica, MA, USA). The membranes were incubated overnight at 4°C with primary mouse polyclonal FOXM1 antibody (Abgent, Inc., San Diego, CA, USA), primary rabbit polyclonal fascin-1 (Boster Biological Technology, Ltd), or primary rabbit polyclonal ezrin (Cell Signaling Technology, Inc., Danvers, MA, USA), or primary rabbit polyclonal paxillin antibody (Immunoway Biotechnology Company, Newark, DE, USA). After washing three times in TBS-T, horseradish peroxidase (HRP)-conjugated secondary antibodies were used at a dilution of 1:5000 in TBS-T for 2 h at room temperature. After three additional washes with TBS-T, the immunoreactive bands were visualized with a chemiluminescence reagent (ECL; Millipore Biotechnology) and quantified using a Bio-Rad imaging system (Bio-Rad Laboratories, Inc., Hemel Hempstead, Hertfordshire, UK).
Cell invasion and migration assays
A Transwell chamber was used (8 mm, 24-well format; Corning, Lowell, MA, USA) to perform cell invasion and migration assays. For the migration assay, 5 × 104 cells in serum-free media were added to the upper chamber after treatment with 2, 4, or 6 µM thiostrepton for 48 h. For the invasion assay, 1 × 105 cells in serum-free medium were added to the upper chamber coated with diluted Matrigel (Sigma-Aldrich, St. Louis, USA) after treatment with 2, 4, or 6 µM thiostrepton for 48 h. Media containing 10% FBS were placed in the lower chambers. After 24 h of incubation at 37°C with 5% CO2, the medium was removed from the upper chamber. The non-invaded cells on the upper side of the chamber were gently scraped off with a cotton swab, whereas the cells that had migrated or invaded through the membrane were stained with methanol and 0.1% crystal violet, imaged, and counted using an inverted microscope (Olympus, Tokyo, Japan) and quantified from visualizing five random fields at a magnification of ×200. Experiments were independently repeated three times.
MMP-2 and MMP-9 assay
Cells were seeded to 50% confluence before treatment with thiostrepton or DMSO. The supernatants were then harvested after 48 h. MMP-2 and MMP-9 activity assays were performed using MMP-2 and MMP-9 ELISA kits (Colorfulgene Biological Technology, Ltd, Wuhan, China).
Statistical analysis
For statistical evaluation, SPSS software (version 14.0; SPSS, Inc., Chicago, IL, USA) was used. The Mann–Whitney U test, Kruskall–Wallis test, Student’s t test, and two-way ANOVA were used for comparing groups. The Spearman correlation test was used to analyze the relationship between FOXM1 expression and clinicopathologic characteristics. A p < 0.05 was considered statistically significant.
Results
FOXM1 expression is up-regulated in NPC
First, 65 NPC specimens at different grades of malignancy and 20 non-cancerous nasopharyngeal tissue specimens were tested for FOXM1 expression by immunohistochemistry. FOXM1 was expressed predominantly in the cytoplasm of tumor cells. As is known, FOXM1 utilization in adults is restricted to organs with proliferating cells involved in replenishing differentiated cell populations (Citation21). Therefore, low expression of FOXM1 was detected in normal nasopharyngeal epithelial cells, whereas there was nearly no expression of FOXM1 in submucous tissues (). Positive staining of FOXM1 was observed in 66.2% (43/65) of NPC samples and 40.0% (8/20) of normal tissues (p < 0.01) (). To confirm the above results, the levels of FOXM1 mRNA and protein expression were determined by qRT-PCR and Western blot, respectively, in NPC and normal nasopharyngeal tissues. The expression of FOXM1 mRNA and protein was significantly up-regulated in NPC compared with normal nasopharyngeal tissues (both p < 0.05) ().
Figure 1. Expression of FOXM1 in NPC and normal nasopharyngeal tissues (NT). A: Overexpression of FOXM1 in NPC and low expression in NT as evidenced by immunohistochemical staining. B: Relative FOXM1 mRNA levels in NPC and NT were determined by qRT-PCR. C: Overexpression of FOXM1 in NPC and low expression in NT were determined by Western blot. Experiments were independently repeated three times. *p < 0.05.
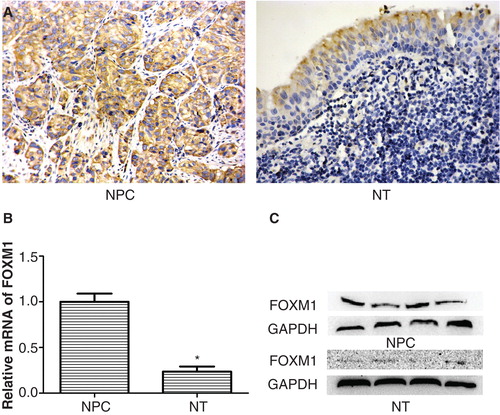
Table I. Expression of FOXM1 in NPC and normal tissue specimens.
Higher level of FOXM1 expression in NPC patients is significantly correlated with metastases
Next, the relationship between FOXM1 expression and the pathologic features of NPC patients was determined. The positive expression of FOXM1 was not related to age, gender, histologic classification, T stage, or M classification. Of note, an increased expression of FOXM1 was significantly correlated with lymph node metastasis (p < 0.05). Specifically, 41 of 58 tumors with lymph node metastasis (+) had high expression of FOXM1, whereas only two of seven tumors without lymph node metastasis – had low expression of FOXM1. Moreover, 73.5% of high-stage tumors (stages III and IV) expressed FOXM1, whereas this was the case in 43.8% of lower-stage tumors (stages I and II) (). Spearman correlation analysis also showed that the expression ratio of FOXM1 was positively correlated with lymph node metastasis (rs = 0.273, p = 0.028) and clinical stage (rs = 0.249, p = 0.046).
Table II. Correlation between the clinicopathologic features and expression of FOXM1.
Thiostrepton inhibits the expression of FOXM1 mRNA and protein in C666-1, but has less of an effect on NP69
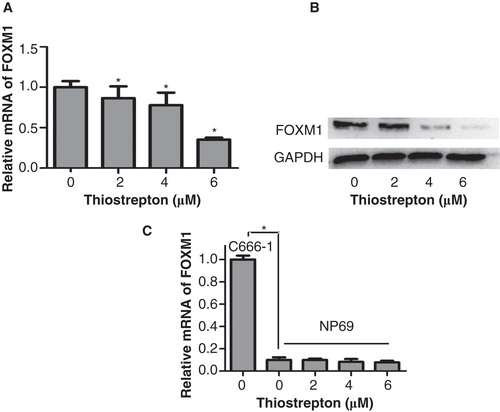
More than 90% of NPC patients in China have undifferentiated tumors; EBV is consistently present in undifferentiated NPC (Citation22). Therefore, the EBV-positive cell line, C666-1, was more suitable for use in the current study than other EBV-negative NPC cell lines. Thiostrepton has been previously reported to inhibit the expression of FOXM1 in several cancer cell lines (Citation18,23); however, the effects of thiostrepton on NPC and normal nasopharyngeal epithelial cell lines remain unknown. The NPC cell line, C666-1, and the transformed human nasopharyngeal epithelial cell line, NP69, were treated with 2, 4, or 6 µM thiostrepton for 48 h, and FOXM1 expression was detected by qRT-PCR and Western blot. The expression of FOXM1 mRNA in C666-1 cells was decreased in a dose-dependent manner after treatment with thiostrepton compared with the control group (). Similarly, the expression of FOXM1 protein in C666-1 cells was also decreased in a dose-dependent manner; notably, there was virtually no expression of FOXM1 protein after treatment with 6 µM thiostrepton (). However, qRT-PCR analysis suggested that FOXM1 is expressed at a lower level in NP69 cells compared with C666-1 cells, and was minimally inhibited after thiostrepton treatment ().
Figure 2. Thiostrepton significantly reduces the expression of FOXM1 mRNA and protein in a NPC cell line other than a nasopharyngeal epithelial cell line. A: The expression of FOXM1 mRNA in C666-1 cells was inhibited after thiostrepton treatment at 2, 4, or 6 µM for 48 h. B: Thiostrepton down-regulated the expression of FOXM1 protein in C666-1 cells. C: The expression of FOXM1 mRNA in NP69 cells was lower than in C666-1 cells and was less affected by thiostrepton. Experiments were independently repeated three times. *p < 0.05.
Down-regulation of FOXM1 by thiostrepton inhibits invasion and migration of NPC cells
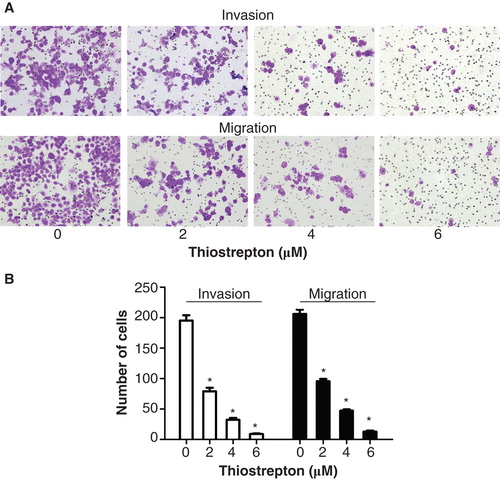
Because higher expression of FOXM1 in NPC patients was significantly related to metastases, we hypothesized that down-regulation of FOXM1 could inhibit the migration and invasion capacity of NPC cells. C666-1 cells were treated with 2, 4, or 6 µM thiostrepton in advance. An invasion chamber assay was used to verify the biological function of thiostrepton in NPC cell invasion. After 24 h, the invading cells were fixed, stained, and observed microscopically. As the representative micrographs clearly demonstrate, down-regulation of FOXM1 by thiostrepton can indeed inhibit NPC cell invasion in vitro in a dose-dependent manner (). Likewise, thiostrepton repressed the migration ability of NPC cells in a dose-dependent fashion compared with the control group ().
Figure 3. Thiostrepton reduces the invasion and migration activities of NPC cells. A: Invasion and migration assays were used to detect the invasion and migration ability of C666-1 cells following treatment with 2, 4, or 6 µM thiostrepton for 48 h. B: Manual counts of invasion and migration experiments. Experiments were independently repeated three times. *p < 0.05.
Thiostrepton decreased the expression of MMP-2, MMP-9, fascin-1, and paxillin in NPC cells
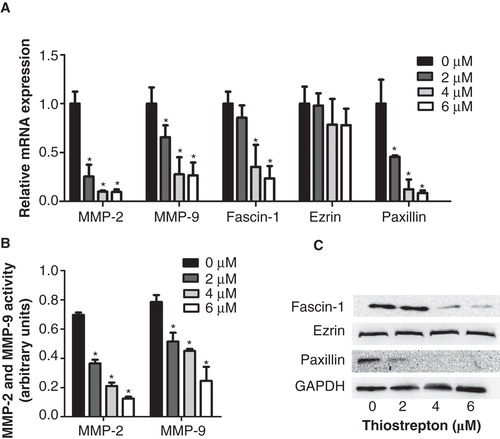
To determine the mechanism by which thiostrepton inhibited the invasion and migration of NPC cells other than direct repression of FOXM1, we first determined the expression of MMP-2 and MMP-9 mRNA and protein by qRT-PCR and ELISA in C666-1 cells. The expression of MMP-2 and MMP-9 mRNA and protein decreased with increasing concentrations of thiostrepton (). Furthermore, the expression of fascin-1, ezrin, and paxillin, essential components of cancer motility, was determined in C666-1 cells. The expression of fascin-1 and paxillin mRNA and protein decreased in a dose-dependent manner, and a significant reduction was observed at concentrations of 2 or 4 µM (); however, the expression of ezrin mRNA and protein was not significantly down-regulated ().
Figure 4. Thiostrepton reduces mRNA and protein expressions of metastatic associated genes in NPC cells. A: Relative MMP-2, MMP-9, fascin-1, ezrin, and paxillin mRNA levels were determined by qRT-PCR. B: An ELISA assay was used to detect protein levels of MMP-2 and MMP-9. C: Fascin-1, ezrin, and paxillin protein levels were determined by Western blot. Experiments were independently repeated three times. *p < 0.05.
Discussion
Regional and systemic metastases seriously affect the cure and survival rates of NPC patients. Recent studies have shown that FOXM1 and its inhibitor, thiostrepton, represented an attractive therapeutic target and novel anticancer drug in the fight against cancer (Citation24-27). The current study showed, for the first time, that overexpression of FOXM1 is associated with NPC metastases. Thiostrepton inhibited the expression of FOXM1 in a NPC cell line, but had minimal effects on a normal nasopharyngeal epithelial cell line. Thiostrepton inhibited the migration and invasion of NPC cells through down-regulation of MMP-2, MMP-9, fascin-1, ezrin, and paxillin.
FOXM1 is frequently up-regulated in various human cancers and is able to enhance cell proliferation, migration, and invasion (Citation28). Overexpression of FOXM1 has been reported in cervical intraepithelial neoplasia (CIN) and carcinoma compared with normal epithelium (Citation12). Overexpression of FOXM1b promotes the growth and metastasis of gastric cancer cells and gastric tumorigenesis by transactivation of vascular endothelial growth factor expression and elevation of angiogenesis (Citation13). In the current study we showed that FOXM1 is prominently expressed in 66.2% (43/65) of NPC tissue specimens, 69.8% (30/43) of which were > grade 2; however, only 40.0% (8/20) of normal nasopharyngeal mucous tissues expressed FOXM1. Taken together, these results suggest that FOXM1 is closely linked to nasopharyngeal carcinogenesis.
A study involving pulmonary squamous cell carcinoma (SCC) revealed that FOXM1 expression is correlated with poor histologic differentiation, lymph node metastasis, and advanced tumor stage (Citation9). In gliomas, the level of FOXM1 expression is significantly higher in glioblastoma multiforme than anaplastic astrocytomas and low-grade astrocytomas (Citation29); however, FOXM1 expression is not correlated with clinicopathologic parameters in non-small cell lung carcinomas (Citation8). These findings suggest that the relationship between FOXM1 and clinicopathologic parameters might be tissue-specific. In the current study we showed that FOXM1 expression was more frequently detected in NPC with lymph node metastasis and advanced tumor stage. The results showed that 41 of 58 lymph node metastasis (+) NPC had high expression of FOXM1, whereas only two of seven lymph node metastasis – NPC had low expression of FOXM1 and 73.5% of higher-stage (III and IV) NPC had high expression of FOXM1 compared to 43.8% of lower-stage (I and II) NPC. Moreover, during recent years it has been shown that overexpression of FOXM1 increases the invasion and migration potential of pancreatic and ovarian cancer cells in vitro (Citation30,31). Therefore, we conclude that higher expression of FOXM1 is also associated with metastases of NPC patients.
In established animal models, FOXM1 is critical for the development and metastasis of chemically induced hepatocellular carcinoma and prostate, lung, and colorectal cancers (Citation32-35). Down-regulation of FOXM1 by thiostrepton inhibits the metastatic potential of ovarian cancer cells (Citation31). Moreover, thiostrepton specifically inhibits FOXM1 expression in breast cancer cells, but not in non-cancer cells (Citation23). In the current study we also showed that thiostrepton inhibits the expression of FOXM1 in NPC cells in a dose-dependent manner, but has a minimal inhibitory effect on normal nasopharyngeal epithelial cells. In addition, we also showed that thiostrepton depresses the invasion and migration of NPC cells in a dose-dependent manner.
The expression of matrix metalloproteinases, particularly MMP-2 and MMP-9, has been strongly implicated in invasion (Citation36,37). Recent studies have reported that FOXM1 up-regulates the expression of MMP-2 and MMP-9 in osteosarcoma cells and transfection of FOXM1 into glioma cells directly activates the MMP-2 promoter, whereas inhibition of FOXM1 expression by FOXM1-siRNA suppresses activation (Citation38,39). In addition, it is known that FOXM1 can be directly inhibited by thiostrepton, and thiostrepton can also inhibit the binding of FOXM1 to genomic target sites. In the current study it was shown that the expression of MMP-2 and MMP-9 was significantly down-regulated by thiostrepton. As mentioned above, FOXM1 directly or indirectly targets MMP-2 and MMP-9 in gliomas and osteosarcoma cells. Thus, we hypothesized that inhibition of FOXM1 by thiostrepton may also directly inhibit MMP-2 expression and indirectly inhibit MMP-9 expression in NPC cells.
The incremental motility of malignant cells is a critical step in metastasis, which is regulated by reorganization of actin cytoskeleton and regulation of focal adhesion (Citation40-42). Fascin-1, ezrin, and paxillin are essential components of these cellular structures. Overexpression of fascin-1, ezrin, and paxillin has been reported in numerous human malignancies and is associated with metastasis in many cancers (Citation43-46); however, there are few studies to date regarding the effects of thiostrepton on the expression of fascin-1, ezrin, and paxillin. In the current study we showed that thiostrepton significantly decreased the expression of fascin-1 and paxillin in NPC cells. In addition, we also observed that ezrin was not down-regulated after treatment with thiostrepton. Because fascin-1 and paxillin have not been reported as the direct or indirect target gene of FOXM1, the mechanisms responsible for these results were not established through the thiostrepton-FOXM1 pathway. Therefore, further studies are needed to determine whether or not FOXM1 directly targets fascin-1 and paxillin or indirectly through another endogenous gene.
In summary, we have characterized the expression of FOXM1 in NPC tissue specimens, its relationship with clinicopathologic factors, the effects of the FOXM1 inhibitor, thiostrepton, on invasion and migration ability of NPC cells in vitro, and the molecular mechanism involved. Our results suggest that overexpression of FOXM1 is involved in NPC development and metastases. Moreover, inhibition of FOXM1, MMP-2, MMP-9, fascin-1, and paxillin expression by thiostrepton suppresses the metastases of NPC cells. Based on these findings, additional studies are needed to elucidate the exact relationship between thiostrepton and FOXM1 and the above metastatic-related gene. In addition, it is possible that thiostrepton may synergize with chemotherapeutic drugs in NPC combination treatment to improve the efficacy of currently available treatments.
Acknowledgements
This work was supported by the Natural Science Foundation of China (grant number: 81272980) and the National Key Clinical Specialties Construction Program of China ([2012]649).
Declaration of interest: The authors report no conflicts of interest. The authors alone are responsible for the content and writing of the paper.
References
- Tang F, Zou F, Peng Z, Huang D, Wu Y, Chen Y, et al. N,N’-dinitrosopiperazine-mediated ezrin protein phosphorylation via activation of Rho kinase and protein kinase C is involved in metastasis of nasopharyngeal carcinoma 6-10B cells. J Biol Chem. 2011;286:36956–67.
- Leung TW, Tung SY, Sze WK, Wong FC, Yuen KK, Lui CM, et al. Treatment results of 1070 patients with nasopharyngeal carcinoma: an analysis of survival and failure patterns. Head Neck. 2005;27:555–65.
- Chua DT, Ma J, Sham JS, Mai HQ, Choy DT, Hong MH, et al. Long-term survival after cisplatin-based induction chemotherapy and radiotherapy for nasopharyngeal carcinoma: a pooled data analysis of two phase III trials. J Clin Oncol. 2005;23:1118–24.
- Razak AR, Siu LL, Liu FF, Ito E, O’Sullivan B, Chan K. Nasopharyngeal carcinoma: the next challenges. Eur J Cancer. 2010;46:1967–78.
- Leung TW, Lin SS, Tsang AC, Tong CS, Ching JC, Leung WY, et al. Over-expression of FoxM1 stimulates cyclin B1 expression. FEBS Lett. 2001;507:59–66.
- Wang X, Kiyokawa H, Dennewitz MB, Costa RH. The Forkhead Box m1b transcription factor is essential for hepatocyte DNA replication and mitosis during mouse liver regeneration. Proc Natl Acad Sci USA. 2002;99:16881–6.
- Wang IC, Chen YJ, Hughes D, Petrovic V, Major ML, Park HJ, et al. Forkhead box M1 regulates the transcriptional network of genes essential for mitotic progression and genes encoding the SCF (Skp2-Cks1) ubiquitin ligase. Mol Cell Biol. 2005;25:10875–94.
- Gialmanidis IP, Bravou V, Amanetopoulou SG, Varakis J, Kourea H, Papadaki H. Overexpression of hedgehog pathway molecules and FOXM1 in non-small cell lung carcinomas. Lung Cancer. 2009;66:64–74.
- Yang DK, Son CH, Lee SK, Choi PJ, Lee KE, Roh MS. Forkhead box M1 expression in pulmonary squamous cell carcinoma: correlation with clinicopathologic features and its prognostic significance. Hum Pathol. 2009;40:464–70.
- Li Q, Zhang N, Jia Z, Le X, Dai B, Wei D, et al. Critical role and regulation of transcription factor FoxM1 in human gastric cancer angiogenesis and progression. Cancer Res. 2009;69:3501–9.
- Zeng J, Wang L, Li Q, Li W, Björkholm M, Jia J, et al. FoxM1 is up-regulated in gastric cancer and its inhibition leads to cellular senescence, partially dependent on p27 kip1. J Pathol. 2009;218:419–27.
- Chan DW, Yu SY, Chiu PM, Yao KM, Liu VW, Cheung AN, et al. Over-expression of FOXM1 transcription factor is associated with cervical cancer progression and pathogenesis. J Pathol. 2008;215:245–52.
- Bektas N, Haaf At, Veeck J, Wild PJ, Lüscher-Firzlaff J, Hartmann A, et al. Tight correlation between expression of the Forkhead transcription factor FOXM1 and HER2 in human breast cancer. BMC Cancer. 2008;8:42.
- Laoukili J, Stahl M, Medema RH. FoxM1: at the crossroads of ageing and cancer. Biochim Biophys Acta. 2007;1775:92–102.
- Chen H, Yang C, Yu L, Xie L, Hu J, Zeng L, et al. Adenovirus-mediated RNA interference targeting FOXM1 transcription factor suppresses cell proliferation and tumor growth of nasopharyngeal carcinoma. J Gene Med. 2012;14:231–40.
- Hegde NS, Sanders DA, Rodriguez R, Balasubramanian S. The transcription factor FOXM1 is a cellular target of the natural product thiostrepton. Nat Chem. 2011;3:725–31.
- Bhat UG, Zipfel PA, Tyler DS, Gartel AL. Novel anticancer compounds induce apoptosis in melanoma cells. Cell Cycle. 2008;7:1851–5.
- Bhat UG, Halasi M, Gartel AL. Thiazole antibiotics target FoxM1 and induce apoptosis in human cancer cells. PLoS One. 2009;4:e5592.
- Bhat UG, Halasi M, Gartel AL. FoxM1 is a general target for proteasome inhibitors. PLoS One. 2009;4:e6593.
- Taylor CR. Quantifiable internal reference standards for immunohistochemistry: the measurement of quantity by weight. Appl Immunohistochem Mol Morphol. 2006;14:253–9.
- Ye H, Kelly TF, Samadani U, Lim L, Rubio S, Overdier DG, et al. Hepatocyte nuclear factor 3/fork head homolog 11 is expressed in proliferating epithelial and mesenchymal cells of embryonic and adult tissues. Mol Cell Biol. 1997;17:1626–41.
- Cheung ST, Huang DP, Hui AB, Lo KW, Ko CW, Tsang YS, et al. Nasopharyngeal carcinoma cell line (C666-1) consistently harbouring Epstein-Barr virus. Int J Cancer. 1999;83:121–6.
- Kwok JM, Myatt SS, Marson CM, Coombes RC, Constantinidou D, Lam EW. Thiostrepton selectively targets breast cancer cells through inhibition of forkhead box M1 expression. Mol Cancer Ther. 2008;7:2022–32.
- Radhakrishnan SK, Bhat UG, Hughes DE, Wang IC, Costa RH, Gartel AL. Identification of a chemical inhibitor of the oncogenic transcription factor forkhead box M1. Cancer Res. 2006;66:9731–5.
- Gartel AL. FoxM1 inhibitors as potential anticancer drugs. Expert Opin Ther Targets. 2008;12:663–5.
- Radhakrishnan SK, Gartel AL. FOXM1: the Achilles’ heel of cancer? Nat Rev Cancer. 2008;8:c1; author reply c2.
- Adami GR, Ye H. Future roles for FoxM1 inhibitors in cancer treatments. Future Oncol. 2007;3:1–3.
- Halasi M, Gartel AL. FOX(M1) News–it is cancer. Cancer Ther. 2013;12:245–54.
- Liu M, Dai B, Kang SH, Ban K, Huang FJ, Lang FF, et al. FoxM1B is overexpressed in human glioblastomas and critically regulates the tumorigenicity of glioma cells. Cancer Res. 2006;66:3593–602.
- Kong X, Li L, Li Z, Le X, Huang C, Jia Z, et al. Dysregulated expression of FOXM1 isoforms drives progression of pancreatic cancer. Cancer Res. 2013;73:3987–96.
- Chan DW, Hui WW, Cai PC, Liu MX, Yung MM, Mak CS, et al. Targeting GRB7/ERK/FOXM1 signaling pathway impairs aggressiveness of ovarian cancer cells. PLoS One. 2012;7:e52578.
- Kalinichenko VV, Major ML, Wang X, Petrovic V, Kuechle J, Yoder HM, et al. Foxm1b transcription factor is essential for development of hepatocellular carcinomas and is negatively regulated by the p19ARF tumor suppressor. Genes Dev. 2004;18:830–50.
- Kim IM, Ackerson T, Ramakrishna S, Tretiakova M, Wang IC, Kalin TV, et al. The Forkhead Box m1 transcription factor stimulates the proliferation of tumor cells during development of lung cancer. Cancer Res. 2006;66:2153–61.
- Kalin TV, Wang IC, Ackerson TJ, Major ML, Detrisac CJ, Kalinichenko VV, et al. Increased levels of the FoxM1 transcription factor accelerate development and progression of prostate carcinomas in both TRAMP and LADY transgenic mice. Cancer Res. 2006;66:1712–20.
- Yoshida Y, Wang IC, Yoder HM, Davidson NO, Costa RH. The forkhead box M1 transcription factor contributes to the development and growth of mouse colorectal cancer. Gastroenterology. 2007;132:1420–31.
- Egeblad M, Werb Z. New functions for the matrix metalloproteinases in cancer progression. Nat Rev Cancer. 2002;2:161–74.
- Deryugina EI, Quigley JP. Matrix metalloproteinases and tumor metastasis. Cancer Metastasis Rev. 2006;25:9–34.
- Wang IC, Chen YJ, Hughes DE, Ackerson T, Major ML, Kalinichenko VV, et al. FoxM1 regulates transcription of JNK1 to promote the G1/S transition and tumor cell invasiveness. J Biol Chem. 2008;283:20770–8.
- Dai B, Kang SH, Gong W, Liu M, Aldape KD, Sawaya R, et al. Aberrant FoxM1B expression increases matrix metalloproteinase-2 transcription and enhances the invasion of glioma cells. Oncogene. 2007;26:6212–19.
- Yamaguchi H, Condeelis J. Regulation of the actin cytoskeleton in cancer cell migration and invasion. Biochim Biophys Acta. 2007;1773:642–52.
- Gardel ML, Schneider IC, Aratyn-Schaus Y, Waterman CM. Mechanical integration of actin and adhesion dynamics in cell migration. Annu Rev Cell Dev Biol. 2010;26:315–33.
- Benitez-King G, Soto-Vega E, Ramirez-Rodriguez G. Melatonin modulates microfilament phenotypes in epithelial cells: implications for adhesion and inhibition of cancer cell migration. Histol Histopathol. 2009;24:789–99.
- Jawhari AU, Buda A, Jenkins M, Shehzad K, Sarraf C, Noda M, et al. Fascin, an actin-bundling protein, modulates colonic epithelial cell invasiveness and differentiation in vitro. Am J Pathol. 2003;162:69–80.
- Wu DW, Cheng YW, Wang J, Chen CY, Lee H. Paxillin predicts survival and relapse in non-small cell lung cancer by microRNA-218 targeting. Cancer Res. 2010;70:10392–401.
- Akisawa N, Nishimori I, Iwamura T, Onishi S, Hollingsworth MA. High levels of ezrin expressed by human pancreatic adenocarcinoma cell lines with high metastatic potential. Biochem Biophys Res Commun. 1999;258:395–400.
- Gao W, Zhang C, Feng Y, Chen G, Wen S, Huangfu H, et al. Fascin-1, ezrin and paxillin contribute to the malignant progression and are predictors of clinical prognosis in laryngeal squamous cell carcinoma. PLoS One. 2012;7:e50710.