Abstract
Based primarily on studies that employ Pavlovian fear conditioning, extinction of conditioned fear has been found to be mediated by N-methyl-D-aspartate (NMDA) receptors in the amygdala and medial prefrontal cortex. This led to the discovery that an NMDA partial agonist, D-cycloserine, could facilitate fear extinction when given systemically or locally into the amygdala. Because many forms of cognitive behavioral therapy depend on fear extinction, this led to the successful use of D-cycloserine as an adjunct to psychotherapy in patients with so-called simple phobias (fear of heights), social phobia, obsessive-compulsive behavior, and panic disorder. Data in support of these conclusions are reviewed, along with some of the possible limitations of D-cycloserine as an adjunct to psychotherapy.
Principaimente mediante esiudios que empiean el miedo condicionado pavloviano se ha encontrado que la extinción del miedo condicionado es mediada por receptores de N-metil-D-aspartato (NMDA) locaiizados en ia amígdala y en la corteza prefrontal medial. Esto llevó al descubrimiento que la D-cicloserina, un agonista parcial del NMDA, podría facilitar la extinción del miedo al administrarse en forma sistémica o localmente en la amígdala. Ya que muchas formas de terapia cognitivo conductual dependen de la extinción del miedo, se ha empleado con éxito la cicloserina como tratamiento conjunto con la psicoterapia en pacientes con fobias simples (temor a la altura), fobia social, conductas obsesivo-compulsivas y trastorno de pánico. Además se revisa la información que sustenta estas conclusiones y algunas de las posibles limitaciones de la D-cicloserina como tratamienio conjunto con la psicoterapia.
Selon des études qui emploient le conditionnement pavlovien à la peur, les récepteurs NMDA (N-méthyl-D-aspartate) de l'amygdale et du cortex préfrontal médian assurent la médiation de l'extinction de la peur conditionnée ; ces résultats ont conduit à la découverte qu'un agoniste partiel au NMDA, la D-cyclosérine, pouvait faciliter l'extinction de la peur après administration par voie générale ou locale dans l'amygdale. Puisque de nombreuses formes de thérapie cognitivo-comportementale dépendent de l'extinction de la peur, la D-cyclosérine a été utilisée avec succès comme aide à la psychothérapie chez des patients atteints de phobies dites simples (vertige), de phobie sociale, de comportement obsessionnel compulsif et de trouble panique. Des données en faveur de ces conclusions sont proposées ainsi que certaines limites éventuelles de la D-cyclosérine comme traitement d'appoint à la psychothérapie.
Introduction
This section will discuss extinction of conditioned fear and how it is mediated by a protein called the N-methyl-D-aspartate (NMDA) receptor in the amygdala and medial prefrontal cortex. This will be followed by a review of the literature showing that a compound called D-cycloserine, which facilitates the NMDA receptor, speeding up extinction in animals and psychotherapy in people. Much of this progress can be attributed to the use of Paviovian fear conditioning as a model system. In this paradigm, an initially innocuous stimulus, the to-be conditioned stimulus (eg, a light, tone, or distinctive place) is paired with an innately aversive unconditioned stimulus (eg, a footshock in rats, a blast of air to the throat in humans) and the subject comes to exhibit a conditioned fear response to the conditioned stimulus. In rodents, fear is defined operationally as a cessation of all bodily movements except those required for respiration (freezing), an increase in the amplitude of an acoustically elicited startle response (fear-potentiated startle), an increase in blood pressure, changes in respiration, emission of ultrasonic distress calls, avoidance of the place where shock occurred, or several other possible measures, in the presence of the conditioned stimulus. In humans fear is typically measured as a change in skin conductance and increased startle when elicited in the presence of the conditioned stimulus. Unlike Pavlov's dog, which salivated when it heard the metronome, just as it did when it swallowed the dry food powder, the fear response may or may not mimic the unconditioned response to the aversive stimulus. For example, rats jump around when they are shocked, yet the conditioned fear response is just the opposite; they freeze and hold very still. Hence, fear is really a hypothetical construct that is used to describe the constellation of behaviors that are seen following fear conditioning, and these may or may not mimic what happens in the presence of the unconditioned stimulus.
Fear is a highly adaptive form of learning that prevents us from returning to a place where we were harmed (the alley where you were raped) or distraught (the airplane where we had a very bumpy flight) or contacting something that was harmful (a hot burner on a stove). Fear conditioning can be produced by a single training trial, and fear memories can last a lifetime. Normally, fear memories are suppressed by the process called fear extinction or habituation when the situation signals that these cues are no longer dangerous (eg, a soldier returning from combat) or when they are experienced over and over again in the absence of any negative consequence (eg, multiple smooth airplane flights). However, fear can become pathological if a person continues to be afraid in situations where they no longer should be afraid. For example, a soldier who is still afraid of a helicopter or the sound of a car backfiring long after he returned from service is no longer adaptive; he has a deficit in extinction or the ability to respond appropriately to safety signals (eg, as seen in post-traumatic stress disorder).
Extinction—behavioral characteristics
As mentioned above, extinction of fear refers to the reduction in the measured level of fear to a cue previously paired with an aversive event when that cue is presented repeatedly in the absence of the aversive event. Actually, the term extinction is used in several different ways in the literature. Extinction may refer to: (i) the experimental procedure used to produce a decrement in the fear response; (ii) the decremental effect of this procedure on the fear response, which can be measured both at the time the cue is presented in the absence of the aversive event and at a later time; or (iii) the hypothesized associative or cellular process responsible for that effect. As suggested elsewhere,Citation1 we will define the experimental procedure as extinction training, the decrement in the fear response measured during extinction training as within-session extinction, and the decrement measured at some interval after extinction training as extinction retention. The term extinction will be reserved for the process underlying the loss of the fear response.
Extinction is not the same as forgetting
Although some forgetting of the original conditioned fear association may occur in extinction (see outstanding review),Citation2 numerous studies show that extinction cannot fully be explained by forgetting because it requires exposure to the conditioned stimulus in the absence of the aversive event as opposed to the simple passage of time.
Extinction is generally cue-specific
Most studies show that fear extinction is cue-specific. For example, if a tone is paired with a shock and a light is paired with a shock, and then extinction training is only given to the tone, fear of the light will be undiminished. Generalization gradients of extinction are typically seen where the magnitude of extinction is greatest to the cue given during extinction training, and less so to cues along some continuum, such as a series of different auditory frequencies that received no extinction training.Citation3 Generalization of extinction is negligible across cues drawn from different sensory modalities, or drawn from a single modality but differing substantially in their physical characteristics.
Extinction generally is not permanent
The decrement in conditioned fear responses during and shortly after extinction training generally is not permanent, as there are several instances in which extinguished fear responses are observed to reappear.
Reinstatement
This refers to the reappearance of a fear response following exposure to unsignaled presentations of the unconditioned stimulus after the completion of extinction training.Citation4 Unsignaled unconditioned stimulus presentations must occur within the context in which animals ultimately are tested if a return of fear is to be observed.Citation5,Citation6 Thus, reinstatement seems to depend on context conditioning and is likely to involve summation of two fear-inducing tendencies, each behaviorally subthreshold when considered independently, but suprathreshold when combined: weak conditioning to context and residual conditioned fear to the extinguished stimulus (see ref 7).
Renewal
Renewal refers to a reappearance of extinguished fear when animals are tested in a context different from the one in which extinction training took place. For example, when animals are first trained to fear a light in context A, then receive extinction training to the light in context B, and finally are tested for fear to the light in either context A or context B, different outcomes are obtained: animals tested in context B (the same context where extinction training took place) exhibit little fear to the light, whereas animals tested in context A exhibit robust fear to the light.Citation6,Citation8 A similar postextinction return of fear is observed when animals are tested in a third, novel context C following acquisition in context A and extinction in context B.Citation8,Citation9 Thus, rather than learning that “now the cue is no longer paired with the shock,” the animal learns that “now, in this place, the cue is no longer paired with the shock.”
Spontaneous recovery
Spontaneous recovery refers to a reappearance of fear with the passage of time following extinction training in the absence of any further explicit training.Citation10
So, extinction is not a full erasure of the original fear memory but instead an active form of learning that acts to suppress or inhibit the original fear memory. This second learning process is referred to as “inhibitory” learning, as opposed to the original “excitatory” learning that occurred during pairings between the conditioned and the unconditioned stimulus. These two types of learning work at cross-purposes in terms of their tendency to stimulate or oppose, respectively, fear output, eg, refs 11-13. In other words, the conditioned stimulus emerges from extinction training with two meanings: following acquisition, the conditioned stimulus signals that an aversive event is coming, and following extinction, the conditioned stimulus signals that an aversive event will be withheld.Citation11
To account for recovery of fear following extinction, the inhibitory learning that accrues in extinction may not be expressed either because it is particularly “fragile” and subject to disruption or because it is gated by context, where “context” is defined broadly to include temporal and interoceptive cues, as well as spatial ones.Citation11 That is, following extinction, appropriate behavior (no fear) is expressed within the temporal and spatial context of extinction training, whereas acquisition-appropriate behavior (fear) is expressed at other times and in other places.
Extinction may be “erased” under certain circumstances
Recently, however, new data have emerged in support of a mechanism more consistent with an “unlearning” account of extinction, in which plasticity underlying fear memory is reversed through a process known as synaptic depotentiation. Depotentiation refers to a reversal of long-term potentiation (ie, a return of potentiated synapses to baseline synaptic efficacy) when low-frequency stimulation is applied to afferent pathways shortly following induction of long-term potentiation. Evidence indicates that the biochemical and molecular mechanisms of depotentiation are opposite to those of long-term potentiation. For example, long-term potentiation is associated with membrane insertion of nonNMDA receptors.Citation14 Depotentiation, by contrast, is associated with internalization of the same type of receptors (see ref 15).
Po-Wu Gean and colleagues demonstrated that depotentiation occurs in the amygdala.Citation16,Citation17 For example, depotentiation-inducing low-frequency stimulation of the amygdala in vivo 10 min after fear acquisition blocked the expression of conditioned fear 24 h later, an effect that could be interpreted as a mimicking of extinction.Citation16 These findings are intriguing, but puzzling, because they would seem to offer no explanation of recovery of fear following extinction through reinstatement, renewal, or spontaneous recovery. Although “new learning” and “unlearning” mechanisms of extinction are often presented as mutually exclusive possibilities, it has been acknowledged that both may occur to some extent, eg, ref 2. Interestingly, depotentiation is inducible more readily at short intervals following induction of longterm potentiation and does not seem to be inducible at all at intervals greater than about 1 h (see ref 18). In rodents, extinction studies typically do not use intervals between acquisition and extinction training of less than 24 h, although biochemical processes of extinction were reported to be different when extinction training was conducted immediately following acquisition compared with 1 h or 3 h after extinction training.Citation19
To test the hypothesis that extinction training given shortly after conditioning might “erase” the original fear memory, rats were fear conditioned and then given extinction training either 10 min, 1 h, 24 h, or 3 days later.Citation18 Consistent with an inhibitory learning mechanism of extinction, rats extinguished 24 or 72 h following acquisition exhibited moderate to strong reinstatement, renewal, and spontaneous recovery. By contrast, and consistent with an erasure mechanism, rats extinguished 10 min to 1 h after acquisition exhibited little or no reinstatement, renewal, or spontaneous recovery. These data support a model in which different neural mechanisms are recruited depending on the temporal delay of fear extinction. Based on these results, Dr Barbara Rolhbaum's group at Emory has been testing whether a full therapeutic dose of exposure therapy in the emergency room will lead to stronger fear extinction in traumatized individuals compared with delayed extinction, although the results are not yet fully in.
Extinction training after memory recall may also “erase” fear memories
Very similar results have been found when extinction training was carried out 10 min to 1 h after fear memory recall.Citation20 Rats were trained to associate a tone with a footshock and then divided into five groups. Four groups were given a single retrieval trial presentation of the tone in the absence of a footshock. Extinction training then began either 10 min, 1 h, 6 h, or 24 h later. The fifth group was exposed to the context but did not receive memory retrieval. Twenty-four hours later, all groups were tested to see if they would show between-session extinction and then they were tested once again, 1 month later. Twenty-four hours after extinction all groups had low levels of freezing. However, 1 month later, the groups given extinction training 10 min or 1 h after recall showed no spontaneous recovery, whereas the groups extinguished 6 or 24 h later did. Very similar results were seen when relapse of extinction was measured with renewal or reinstatement. Hence, just like extinction given shortly after fear conditioning seems to block consolidation extinction given shortly after recall seems to block reconsolidation. Importantly, this work was extended in humans and extinction given shortly after recall blocked spontaneous recovery 1 year later!Citation21 These are exciting results and clearly indicate that the timing of extinction either after original learning or after memory recall can have pro found effects on the durability of extinction. It remains unclear, however, as to why a 10-min interval between the first extinction trial (ie, a memory retrieval trial is identical to the first trial of extinction training) produces such a different effect than the usual intertrial interval during normal extinction training.
Role of NMDA receptors in extinction of conditioned fear in rodents
Like fear acquisition,Citation22 fear extinction depends on NMDA receptors within the basolateral amygdala. Thus, intra-amygdala infusions of a compound that blocks NMDA receptors prior to extinction training dose-dependently blocked retention of extinction of fear-potentiated startle measured 1 day after extinction training.Citation23 This impairment could not be attributed to an effect on NMDA receptors outside the amygdala, to damage or destruction of the amygdala, or to an impairment of sensory transmission during extinction training. Later studies showed that systemic administration of NMDA receptor antagonists prior to fear extinction training lead to dose-dependent impairments of both within-session extinction and extinction retention.Citation24-Citation28 Systemic NMDA receptor antagonists also impair extinction retention or reinstatement when administered immediately after extinction training,Citation29-Citation32 indicating that NMDA receptors are involved in consolidation as well as encoding of extinction memory. A similar blockade of extinction of contextual fear conditioning, and inhibitory avoidance conditioning has been reported with both systemic and localized administration of various NMDA receptor antagonists,Citation33,Citation34 and additional studies have confirmed that these effects cannot be explained by state dependency.Citation24,Citation35
Different effects of NMDA blockade in the amygdala and medial prefrontal cortex on extinction
The medial prefrontal cortex (mPFC) sends dense projections to the amygdala that terminate, in part, on inhibitory interneurons.Citation36-Citation40 Hence, mPFC is in a position to inhibit the amygdala, a possible extinction mechanism,Citation41 at least under some circumstances.Citation42,Citation43 Electrolytic lesionsCitation44 or localized inactivationCitation45 of the infralimbic region of mPFC impair extinction retention while having little to no effect on acquisition or within-session extinction, suggesting a role for this region specifically in consolidation and/or expression of extinction memory (see also ref 46). Single units within infralimbic cortex fire selectively to presentations of a previously fear-conditioned cue during an extinction retention test 24 h after extinction training but not during the extinction training session itself.Citation47 When infralimbic cortex microstimulation was paired with presentations of a previously fear conditioned cue in nonextinguished animals, freezing to those cues was attenuated, and this effect was also seen the next day when no stimulation was given.Citation47,Citation48 Collectively, these findings indicate that mPFC plays a significant role in many cases in extinction memory consolidation and expression, likely via its interactions with the amygdala. NMDA receptors within amygdala seem to be involved in the initiation of extinction, whereas in infralimbic cortex, they seem to be involved in consolidation of extinction. Microinfusions of NMDA receptor antagonists into basolateral nucleus of the amygdala prior to fear extinction training impair both within-session extinction and extinction retention.Citation16,Citation23,Citation30,Citation31,Citation33 However, local infusions of NMDA 2A, 2B antagonists into basolateral amygdala block the expression of several fear-related conditioned responses, including freezing, suggesting these drugs could artifactually block extinction retention by interfering with synaptic transmission. However, infusion of ifenprodil, a drug that blocks a subtype of the NMDA receptor but does not block expression of fear conditioned responses, still blocked extinction retention.Citation28,Citation30,Citation31 Immediate post-extinction training infusions into the amygdala of ifenprodil have no effect on subsequent extinction retention when extinction of fear is measured.Citation27,Citation30 This suggests that NMDA receptor-dependent synaptic plasticity within amygdala is involved in encoding extinction of fear, but that the subtype of the NMDA receptor where ifenprodil acts in the amygdala is not required for consolidation of extinction, at least for conditioned fear.
In contrast, pre-extinction training infusions of NMDA receptor antagonists into mPFC have no effect on within-session extinction but generally impair later extinction retentionCitation29,Citation31,Citation49; (but see ref 27). Immediate postextinction infusions of NMDA antagonists into the infralimbic cortex do block extinction retention consistently,Citation27,Citation29-Citation31 providing strong evidence that NMDA receptor-dependent synaptic plasticity within this cortical area is involved primarily in consolidation of extinction memory.
NMDA receptors do not seem to be involved the second time extinction is given
Perhaps surprisingly, when the experimental protocol involves fear acquisition and extinction followed by reacquisition and re-extinction of the same cue, systemic administration of the NMDA receptor antagonist MK801 prior to re-extinction training does not impair subsequent extinction retention. However, it does block reextinction when the extinction retention test occurs in a context different from that of initial acquisition and initial extinction,Citation50 suggesting that NMDA receptor activation is required when extinction events are relatively novel but not when they are relatively familiar.Citation50 On the other hand, novelty does not seem to matter for fear conditioning itself because disruption of the NMDA receptor blocks fear acquisition in both a novel and a familiar context.Citation33,Citation49
Effects of localized infusions of NMDA receptor antagonists prior to second extinction are more complex and are reviewed elsewhere (see ref 51).
Role of D-cycloserine in fear extinction
Because blockade of the NMDA receptor impairs extinction, we wondered whether enhancing the functioning of that receptor would enhance extinction. To test this we administered a compound called D-cycloserine (DCS) either systemically or directly into the rats' amygdala prior to extinction training and then tested retention of extinction the next day.Citation52 DCS does not bind to the NMDA receptor itself, but to another receptor on the NMDA protein called the glycine regulatory site. Activation of this site improves the ability of the NMDA receptor protein to flux calcium which initiates a variety of intracellular events that are critical for extinction. As predicted, when DCS was given in combination with repeated exposure to the feared stimulus without a shock, extinction retention was enhanced, when testing took place after DCS had worn off. This did not occur in control rats that received the drug alone, without extinction training. Based on these results, we concluded that the positive effects of the DCS were specifically connected with extinction and did not result from a general dampening of fear expression.
These effects have now been replicated in a large number of studies.
Systemic administration of DCS either beforeCitation52-Citation61 or afterCitation54 extinction training facilitates extinction. Local infusion of DCS into the basolateral nucleus of the amygdala prior toCitation52,Citation62 or afterCitation54 fear extinction training mimics the effects of systemic administration. Chang and MarenCitation63 recently showed that although DCS infusions directly into infralimbic cortex did not facilitate extinction, these infusions did facilitate the subsequent reextinction of fear when animals were trained in a drug-free state.Citation63
DCS may normalize impaired extinction
There is a growing body of evidence to suggest that DCS reverses fear extinction deficits caused by a variety of factors including stress,Citation64-Citation66 alcohol withdrawal,Citation67 REM sleep deprivation,Citation68 genetically modified mice that have a polymorphism in their BDNF gene,Citation69 or even adolescent rats in which the medial prefrontal cortex has not yet developed fully.Citation70 Perhaps consistent with these stress-related effects, DCS interacts with stress hormones: DCS blocks the extinction-impairing effect of the corticosteroid synthesis inhibitor metyrapone and enhances the extinction-facilitating effects of the synthetic glucocorticoid dexamethasone.Citation71 Judo et alCitation72 showed that changes in prefrontal synaptic efficacy during extinction training did not occur in adult rats exposed to early postnatal stress, and these synaptic changes and resulting deficits in extinction were restored by DCS. These observations may also be consistent with findings in clinical studies described below, that DCS facilitates exposure therapy in clinical, but not subclinical, populations, and that different types of mechanisms could be involved in the two groups of subjects.Citation73
DCS facilitates psychotherapy
Many forms of psychotherapy depend in part on extinction of fear. Patients with fear of snakes avoid snakes and do not allow themselves to extinguish this fear. However, repeated exposure to pictures of snakes, a snake in a jar, or even a live snake are extremely effective in eliminating such simple phobias and are widely used. Panic patients afraid they will have a panic attack driving over a high bridge are taken back to the bridge to show them that they will not always have a panic attack there. Patients with a contamination phobia who are forced to touch the bottom of a toilet seat, but not allowed to wash their hands, learn not only they do not die but they don't even get sick, identical to an extinction trial. Exposure to scenes of combat in people with post-traumatic stress disorder (PTSD) in the presence of a supportive therapist often leads to substantial improvement, and cognitive behavioral therapy has been found to be helpful in many PTSD patients. In all these cases of exposure-based psychotherapy extinction is the fundamental mechanism that is operating.
The finding that DCS can facilitate fear extinction in animalsCitation52 and that fear extinction was so central to many types of psychotherapy suggested that DCS might also be effective in facilitating exposure therapy for fear and anxiety disorders in people. DCS had been FDA-approved for some time as an antibiotic treatment for tuberculosis at high doses. Although this effect had nothing to do with its ability to facilitate extinction it allowed us to test whether it would facilitate exposure-based psychotherapy right away.Citation74
In this study, the ability of DCS to enhance exposure therapy for acrophobia, or fear of heights, using virtual reality exposure therapy, was examined. Previous work had shown improvements on acrophobia outcome measures after seven or eight weekly virtual reality therapy sessions.Citation75 Participants in the DCS study underwent a suboptimal amount of virtual reality therapy for acrophobia (only two virtual reality sessions) and were instructed to take a single dose of study medication before each session. So DCS was only taken twice: prior to each of the two sessions that were separated by average of 12 days. Similar to the rats in the preclinical work, participants receiving DCS exhibited significantly more improvement than did participants receiving placebo, measured either 1 week or 3 months later, long after the drug was out of the body ().Citation74 At the 1-week follow-up, DCS-treated patients exhibited less subjective fear and fewer skin conductance fluctuations in the virtual reality environment.
Reproduced from ref 74: Ressler KJ, Rothbaum BO, Tannenbaum L, et al. Cognitive enhancers as adjuncts to psychotherapy: use of D-cycloserine in phobic individuals to facilitate extinction of fear Arch Gen Psychiatry. 2004:61:1136-1144. Copyright © American Medical Association 2004: ref 77: Hofmann SG, Meuret AE, Smits JA, et al. Augmentation of exposure therapy with D-cycloserine for social anxiety disorder. Arch Gen Psychiatry. 2006:63:298-304. Copyright © American Medical Association 2006: ref 79: Wilhelm S, Buhlmann U, Tolin DF, et al. Augmentation of behavior therapy with D-cycloserine for obsessive-compulsive disorder. Am J Psychiatry. 2008:165:335-341: quiz 409. Copyright © Hanover 2008: ref 80: Otto MW, Tolin DF, Simon NM.etal. Efficacy of dcycloserine for enhancing response to cognitive-behavior therapy for panic disorder. Biol Psychiatry. 2010:67:365-370. Copyright © Elsevier 2010
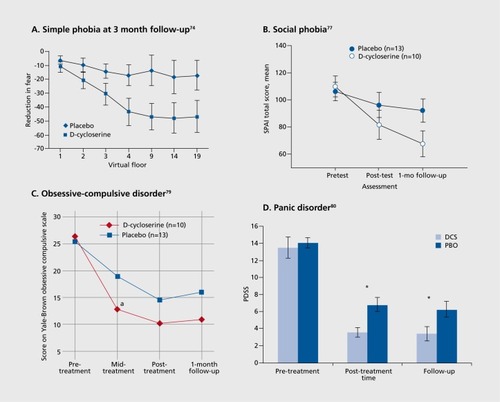
Most importantly, outside of the virtual reality environment patients reported a decrease in overall acrophobia symptoms, increased self-reports of exposure to heights in the “real world,” and higher self-ratings of improvement. These later results are very important because they indicate that extinction of fear is not always context-specific, as seen so often in animal studies. The reason for this appears to be that humans begin to feel safe in situations they previously avoided, once they have some successful psychotherapy and avoid these situations less often. People with fear of elevators do not want to continue to walk up 20 flights of stairs once they learn the elevator will not harm them. In contrast, rats have no opportunity to continue to extinguish because they are put back in their home cage with no further exposure to the fearful conditioned stimulus. So, the several measures of relapse from extinction may be overestimated in rodent studies.
Other groups found that DCS enhanced exposure therapy for social anxiety disorder— ,Citation76,Citation77 obsessivecompulsive disorder— ,Citation78,Citation79 and panic disorder— ,Citation80 indicating that the DCS effect is a relatively general one. The failure of another study to see on effect in OCT)Citation81 may have resulted from giving DCS 4 hours prior to exposure therapy, which may have been too early. There has been one report of a failure of DCS to facilitate exposure therapy for subclinical spider phobia,Citation82 and failures of DCS to facilitate extinction of Pavlovian conditioned fear in a laboratory situation in humans.Citation73,Citation82 However, these negative effects may indicate that DCS is useful only in people with clinically significant, maladaptive fear—consistent, perhaps, with the preclinical data suggesting that DCS is particularly effective in stressed animals (described above).
A note of caution
DCS may not work on re-extinction
It should be recalled that NMDA antagonists block extinction the first time extinction training is carried out but not when rats are retrained and then extinguished again.Citation50 The same is true for D-cycloserine.Citation83
DCS and serotonin reuptake inhibitors
DCS also failed to facilitate extinction in rats with prior administration of a serotonin reuptake inhibitor (imipramine).Citation84 However, because it is likely that many patients in the positive trials of DCS in anxiety disorders were taking serotonin reuptake inhibitors, it is hard to know how important this variable is because the database is just not large enough to allow an adequate evaluation of this variable. In our own study of fear of heights we could find no relationship.
DCS shows tolerance
DCS also failed to facilitate extinction in rats given prior daily injections of DCS,Citation85 consistent with several preclinical studies showing tolerance with repeated DCS treatment.Citation85 Hence, we suggest spacing DCS treatments by at least a week.
DCS should not be given too far in advance of psychotherapy
As mentioned above, DCS is known to facilitate consolidation of fear extinction so it is important not to give it too early prior to psychotherapy. In fact, post-extinction training is used routinely in rodent studies and this maybe especially effective clinically. For example, if a patient had a bad session of psychotherapy it might not be useful to use DCS. But, if they have a good session then the therapist could give DCS right after therapy, which would more likely cover the consolidation period and improve compliance.Citation86 In fact, we have preliminary data in rats that giving DCS prior to sleep, when many types of memory consolidate, may be especially effective (Davis, Bowser, McDevitt, and Walker, in revision).
DCS is very unstable in humid conditions and in solution
It is very important to keep it dry during compounding into lower doses from Seromycin as well as in storage.Citation87
Will DCS make patients worse?
A question that is often asked is why, if DCS is a cognitive enhancer, does it not stamp in the bad memories brought up during psychotherapy and make patients worse? DCS has been shown to facilitate retention of inhibitory avoidance and spatial learning in rats,Citation88 stimulus attributes in inhibitory avoidance in rats,Citation89 inhibitory avoidance in chicksCitation90 or mice,Citation91,Citation92 thirst-motivated maze learning in mice,Citation93 object location in mice,Citation94 taste aversion in rats,Citation95,Citation96 delayed nonmatching- to-sample in rhesus monkeys,Citation97 and acquisition of eyeblink conditioning in rabbits when trace conditioning was used.Citation98 It also improves memory due to aging in mice,Citation91 spatial memory in rats,Citation99 and eyeblink conditioning in rabbits.Citation100 In rats, DCS reverses scopolamine-induced deficits in: the T-maze and water maze,Citation101 working memory,Citation102 or inhibitory avoidance,Citation103 or reduces deficits following brain injuryCitation104 or hippocampal lesions,Citation105 or deficits in inhibitory avoidance in mice caused by β 25-35-amyloid peptidesCitation106 or convulsant drugs.Citation107
Because most, if not all, these tasks depend on the hippocampus, one might expect that DCS would facilitate hippocampally dependent declarative memory in humans. However, the literature is very inconsistent in this area. Otto et alCitation108 found no effect of DCS in improving verbal or nonverbal learning given at weekly sessions nor did D'Souza et alCitation109 on several tests of verbal learning and it had no effect on a procedural task (finger tapping). On the other hand, improvement of procedural learning (sequential finger tapping) but not of declarative (word-pair) learning by DCS was found.Citation110 DCS accelerated rate of learning on item-category associations, but had no beneficial effect in the object-location association task, both declarative memory tasks.Citation111 There was improvement on one cognitive task (delayed thematic recall on the logical memory test) in schizophrenic patients.Citation112 There was one report showing enhanced fear conditioning with DCS in humans,Citation86 but the stydy desing was so complex that it is hard to know what to conclude from this study, especially because there appear to be no positive studies of DCS on classical fear conditioning in humans. Finally, no reports were found of patients getting worse on or after DCS in the six positive studies that have been published with cognitive behavioral therapy. Hence, despite the ability of DCS to facilitate learning in animal studies, for reasons that are not clear, this has not been found universally in humans, even though DCS generally has facilitated fear extinction in clinical populations. Possible reasons for this are discussed elsewhere.Citation113
Conclusion
Because excessive fear and anxiety occur in so many psychiatric disorders, research continues to investigate how the brain normally Inhibits or suppresses these emotions. Exposure-based cognitive behavioral therapy (CBT), in which patients are repeatedly exposed to anxiogenic situations in the absence of any aversive consequences, has been quite successful in treating these disorders. CBT is procedurally similar to fear extinction in animals, in which a fearful stimulus also Is exposed repeatedly without aversive events. Extinction does not erase the original fear memory but instead actively inhibits that memory. It is dependent on a protein called the NMDA receptor in brain areas such as the amygdala and medial prefrontal cortex. A medication called D-cycloserine allows the NMDA receptor to work even better and It also facilitates fear extinction, especially when extinction is compromised following stress. However, it does not work when given alone, but only in combination with extinction training. Six independent clinical trials have shown that D-cycloserine facilitates CBT in patients with phobia, obsessive-compulsive and panic disorder, and several trials are underway to tests its effects in PTSD. Continued analysis of normal and abnormal fear extinction in animals will almost surely lead to other medications to facilitate CBT.
REFERENCES
- MyersKM.DavisM.Behavioral and neural analysis of extinction: a review.Neuron.20023656758412441048
- DelamaterAR.Experimental extinction in Paviovian conditioning: behavioural and neuroscience perspectives.QJ Exp Psychol B.20045797132
- DubinWJ.LevisDJ.Generalization of extinction gradients: a systematic analysis.J Exp Psychol.1974100403412
- RescorlaRA.HethCD.Reinstatement of fear to an extinguished conditioned stimulus.J Exp Psychol Anim Behav Process.1975188961151290
- BoutonME.BollesRC.Role of conditioned contextual stimuli in reinstatement of extinguished fear.J Exp Psychol Anim Behav Process.19795368378528893
- BoutonME.KingDA.Contextual control of conditioned fear: tests for the associative value of the context.J Exp Psychol Anim Behav Process.198392482566886630
- HendryJS.Summation of undetected excitation following exinction of the CER.Animal Learn Behav.198210476482
- BoutonME.BollesRC.Contextual control of the extinction of conditioned fear.Learn Motivation.197910455466
- HarrisJA.JonesML.BaileyGK.WestbrookRF.Contextual control over conditioned responding in an extinction paradigm.J Exp Psychol Anim Behav Process.20002617418510782432
- QuirkGJ.Memory for extinction of conditioned fear is long-lasting and persists following spontaneous recovery.Learn Mem.2002940240712464700
- BoutonME.Context, time, and memory retrieval in the interference paradigms of Paviovian learning.Psychol Bull.199311480998346330
- KonorskiJ.Conditioned Reflexes and Neuronal Organization. London, UK: Cambridge University Press; 1948.
- WagnerAR.SOP: a model of automatic memory processing in animal behavior. In: Spear NE, Miller RR, eds.Information Processing in Animals: Memory Mechanisms. Hillsdale, NJ: Lawrence Erlbaum Associates;1981547
- MalinowR.MalenkaRC.AMPA receptor trafficking and synaptic plasticity.Annu Rev Neurosci.20022510312612052905
- ZhouQ.PooMM.Reversal and consolidation of activity-induced synaptic modifications.Trends Neurosci.20042737838315219736
- LinCH.YehSH.LuHY.GeanPW.The similarities and diversities of signal pathways leading to consolidation of conditioning and consolidation of extinction of fear memory.J Neurosci.2003238310831712967993
- LinCH.LeeCC.HuangYC.WangSJ.GeanPW.Activation of group II metabotropic glutamate receptors induces depotentiation in amygdala slices and reduces fear-potentiated startle in rats.Learn Mem.20051213013715774944
- MyersKM.ResslerKJ.DavisM.Different mechanisms of fear extinction dependent on length of time since fear acquisition.Learn Mem.20061321622316585797
- CainCK.GodsilBP.JamiS.BaradM.The L-type calcium channel blocker nifedipine impairs extinction, but not reduced contingency effects, in mice.Learn Mem.20051227728415930506
- MonfilsMH.CowansageKK.KlannE.LeDouxJE.Extinction-reconsolidation boundaries: key to persistent attenuation of fear memories.Science.200932495195519342552
- SchillerD.MonfilsMH.RaioCM.JohnsonDC.LedouxJE.PhelpsEA.Preventing the return of fear in humans using reconsolidation update mechanisms.Nature.2010463495320010606
- MiserendinoMJ.SananesCB.MeliaKR.DavisM.Blocking of acquisition but not expression of conditioned fear-potentiated startle by NMDA antagonists in the amygdala.Nature.19903457167181972778
- FallsWA.MiserendinoMJ.DavisM.Extinction of fear-potentiated startle: blockade by infusion of an NMDA antagonist into the amygdala.J Neurosci.1992128548631347562
- BakerJD.AzorlosaJL.The IMMDA antagonist MK-801 blocks the extinction of Paviovian fear conditioning.Behav Neurosci.19961106186208889007
- DaltonGL.WangYT.FlorescoSB.PhillipsAG.Disruption of AMPA receptor endocytosis impairs the extinction, but not acquisition of learned fear.Neuropsychopharmacology.2008332416242618046303
- SantiniE.MullerRU.QuirkGJ.Consolidation of extinction learning involves transfer from NMDA- independent to NMDA-dependent memory.J Neurosci.2001219009901711698611
- Sotres-BayonF.Diaz-MataixL.BushDE.LeDouxJE.Dissociable rôles for the ventromedial prefrontal cortex and amygdala in fear extinction: NR2B contribution.Cereb Cortex.20091947448218562331
- Sotres-BayonF.BushDE.LeDouxJE.Acquisition of fear extinction requires activation of NR2B-containing NMDA receptors in the lateral amygdala.Neuropsychopharmacology.2007321929194017213844
- Burgos-RoblesA.Vidal-GonzalezI.SantiniE.QuirkGJ.Consolidation of fear extinction requires NMDA receptor-dependent bursting in the ventromedial prefrontal cortex.Neuron.20075387188017359921
- LaurentV.MarchandAR.WestbrookRF.The basolateral amygdala is necessary for learning but not relearning extinction of context conditioned fear.Learn Mem.20081530431418463174
- LaurentV.WestbrookRF.Distinct contributions of the basolateral amygdala and the medial prefrontal cortex to learning and relearning extinction of context conditioned fear.Learn Mem.20081565766618772253
- SantiniE.GeH.RenK.Pena de OrtizS.QuirkGJ.Consolidation of fear extinction requires protein synthesis in the medial prefrontal cortex.J Neurosci.2004245704571015215292
- LeeH.KimJJ.Amygdalar NMDA receptors are critical for new fear learning in previously fear-conditioned rats.J Neurosci.199818844484549763487
- SzapiroG.ViannaMR.McGaughJL.MedinaJH.IzquierdoI.The role of NMDA glutamate receptors, PKA, MAPK, and CAMKII in the hippocampus in extinction of conditioned fear.Hippocampus.200313535812625457
- CoxJ.WestbrookRF.The NMDA receptor antagonist MK-801 blocks acquisition and extinction of conditioned hypoalgesia responses in the rat.Q J Exp Psychol.1994476187210
- BerrettaS.PantazopoulosH.CalderaM.PantazopoulosP.PareD.Infralimbic cortex activation increases c-Fos expression in intercalated neurons of the amygdala.Neuroscience.200513294395315857700
- RosenkranzJA.GraceAA.Dopamine attenuates prefrontal cortical suppression of sensory inputs to the basolateral amygdala of rats.J Neurosci.2001214090410311356897
- RosenkranzJA.GraceAA.Cellular mechanisms of infralimbic and prelimbic prefrontal cortical inhibition and dopaminergic modulation of basolateral amygdala neurons in vivo.J Neurosci.20022232433711756516
- RosenkranzJA.MooreH.GraceAA.The prefrontal cortex regulates lateral amygdala neuronal plasticity and responses to previously conditioned stimuli.J Neurosci.200323110541106414657162
- QuirkGJ.LikhtikE.PelletierJG.PareD.Stimulation of medial prefrontal cortex decreases the responsiveness of central amygdala output neurons.J Neurosci.2003238800880714507980
- QuirkGJ.MuellerD.Neural mechanisms of extinction learning and retrieval.Neuropsychopharmacology.200833567217882236
- GarciaR.ChangCH.MarenS.Electrolytic lesions of the medial prefrontal cortex do not interfere with long-term memory of extinction of conditioned fear.Learn Mem.200613141716418435
- GewirtzJC.FallsWA.DavisM.Normal conditioned inhibition and extinction of freezing and fear potentiated startle following electrolytic lesions of medial prefrontal cortex.Behav Neurosci.19971117127269267649
- QuirkGJ.RussoGK.BarronJL.LebronK.The role of ventromedial prefrontal cortex in the recovery of extinguished fear.J Neurosci.2000206225623110934272
- Sierra-MercadoD.Padilla-CoreanoN.QuirkGJ.Dissociable roles of prelimbic and infralimbic cortices, ventral hippocampus, and basolateral amygdala in the expression and extinction of conditioned fear.Neuropsychopharmacology.20113652953820962768
- HsuE.PackardMG.Medial prefrontal cortex infusions of bupivacaine or AP-5 block extinction of amphetamine conditioned place preference.Neurobiol Learn Mem.20088950451217905604
- MiladMR.QuirkGJ.Neurons in medial prefrontal cortex signal memory for fear extinction.Nature.2002420707412422216
- MiladMR.Vidal-GonzalezI.QuirkGJ.Electrical stimulation of medial prefrontal cortex reduces conditioned fear in a temporally specific manner.Behav Neurosci.200411838939415113265
- LaurentV.WestbrookRF.Inactivation of the infralimbic but not the prelimbic cortex impairs consolidation and retrieval of fear extinction.Learn Mem.20091652052919706835
- ChanWY.McNallyGP.Conditioned stimulus familiarity determines effects of MK-801 on fear extinction.Behav Neurosci.200912330331419331454
- MyersKM.CarlezonWA Jr.DavisM.Glutamate receptors in extinction and extinction-based therapies for psychiatric illness.Neuropsychopharmacology.201136910
- WalkerDL.ResslerKJ.LuK-T.DavisM.Facilitation of conditioned fear extinction by systemic administration or intra-amygdala infusions of Dcycloserine as assessed with fear-potentiated startle in rats.J Neurosci.2002222343235111896173
- BoutonME.VurbicD.WoodsAM.D-cycloserine facilitates context-specific fear extinction learning.Neurobiol Learn Mem.20089050451018672079
- LedgerwoodL.RichardsonR.CranneyJ.Effects of D-cycloserine on extinction of conditioned freezing.Behav Neurosci.200311734134912708530
- LedgerwoodL.RichardsonR.CranneyJ.D-cycloserine facilitates extinction of learned fear: effects on reacquisition and generalized extinction.Biol Psychiatry.20055784184715820704
- LeeJL.MiltonAL.EverittBJ.Reconsolidation and extinction of conditioned fear: inhibition and potentiation.J Neurosci.200626100511005617005868
- MaoSC.LinHC.GeanPW.Augmentation of fear extinction by Dcycloserine is blocked by proteasome inhibitors.Neuropsychopharmacology.2008333085309518368037
- WalkerDL.DavisM.Light enhanced startle: further pharmacological and behavioral evaluation.Psychopharmacology.200215930431011862363
- WeberM.HartJ.RichardsonR.Effects of D-cycloserine on extinction of learned fear to an olfactory cue.Neurobiol Learn Mem.200787476 48217275356
- WoodsA.BoutonM.D-cycloserine facilitates extinction but does not eliminate renewal of the conditioned emotional response.Behav Neurosci.20061201159116217014266
- YangYL.LuKT.Facilitation of conditioned fear extinction by d-cycloserine is mediated by mitogen-activated protein kinase and phosphatidylinositol 3-kinase cascades and requires de novo protein synthesis in basolateral nucleus of amygdala.Neuroscience.200513424726015951121
- MaoSC.HsiaoYH.GeanPW.Extinction training in conjunction with a partial agonist of the glycine site on the NMDA receptor erases memory trace.J Neurosci.2006268892889916943544
- ChangCH.MarenS.Medial prefrontal cortex activation facilitates reextinction of fear in rats.Learn Mem.20111822122521430044
- MatsumotoM.TogashiH.KonnoK.et al.Early postnatal stress alters the extinction of context-dependent conditioned fear in adult rats.Pharmacol Biochem Behav.2008 8924725218262260
- Nic DhonnchadhaBA.SzalayJJ.Achat-MendesC.et al.D-cycloserine deters reacquisition of cocaine self-administration by augmenting extinction learning.Neuropsychopharmacology.20103535736719741593
- YamamotoS.MorinobuS.FuchikamiM.KurataA.KozuruT.YamawakiS.Effects of single prolonged stress and D-cycloserine on contextual fear extinction and hippocampal NMDA receptor expression in a rat model of PTSD.Neuropsychopharmacology.2008332108211617957211
- BertottoME.BustosSG.MolinaVA.MartijenaID.Influence of ethanol withdrawal on fear memory: effect of d-cycloserine.Neuroscience.200614297999016934411
- SilvestriAJ.RootDH.Effects of REM deprivation and an NMDA agonist on the extinction of conditioned fear.Physiol Behav.20089327428117920644
- YuH.WangY.PattwellS.et al.Variant BDNF Val66Met polymorphism affects extinction of conditioned aversive memory.J Neurosci.2009294056406419339601
- McCallumJ.KimJH.RichardsonR.Impaired extinction retention in adolescent rats: effects of D-cycloserine.Neuropsychopharmacology.2010352134214220592716
- YangYL.ChaoPK.RoLS.WoYY.LuKT.Glutamate NMDA receptors within the amygdala participate in the modulatory effect of glucocorticoids on extinction of conditioned fear in rats.Neuropsychopharmacology.2007321042105117047672
- JudoC.MatsumotoM.YamazakiD.et al.Early stress exposure impairs synaptic potentiation in the rat medial prefrontal cortex underlying contextual fear extinction.Neuroscience.20101691705171420600655
- GrillonC.D-cycloserine facilitation of fear extinction and exposurebased therapy might rely on lower-level, automatic mechanisms.Biol Psychiatry.20096663664119520359
- ResslerKJ.RothbaumBO.TannenbaumL.et al.Cognitive enhancers as adjuncts to psychotherapy: use of D-cycloserine in phobic individuals to facilitate extinction of fear.Arch Gen Psychiatry.2004611136114415520361
- RothbaumBO.HodgesLF.KooperR.OpdykeD.WillifordJS.NorthM.Effectiveness of computer-generated (virtual reality) graded exposure in the treatment of acrophobia.Am J Psychiatry.19951526266287694917
- GuastellaAJ.RichardsonR.LovibondPF.et al.A randomized controlled trial of D-cycloserine enhancement of exposure therapy for social anxiety disorder.Biol Psychiatry.20086354454918179785
- HofmannSG.MeuretAE.SmitsJA.et al.Augmentation of exposure therapy with D-cycloserine for social anxiety disorder.Arch Gen Psychiatry.20066329830416520435
- KushnerMG.KimSW.DonahueC.et al.D-cycloserine augmented exposure therapy for obsessive-compulsive disorder.Biol Psychiatry.20076283583817588545
- WilhelmS.BuhlmannU.TolinDF.et al.Augmentation of behavior therapy with D-cycloserine for obsessive-compulsive disorder.Am J Psychiatry. quiz 409.2008165335341
- OttoMW.TolinDF.SimonNM.et al.Efficacy of d-cycloserine for enhancing response to cognitive-behavior therapy for panic disorder.Biol Psychiatry.20106736537019811776
- StorchEA.MerloLJ.BengtsonM.et al.D-cycloserine does not enhance exposure-response prevention therapy in obsessive-compulsive disorder.Int Clin Psychopharmacol.20072223023717519647
- GuastellaAJ.DaddsMR.LovibondPF.MitchellP.RichardsonR.A randomized controlled trial of the effect of D-cycloserine on exposure therapy for spider fear.J Psychiatr Res.20074146647116828803
- LangtonJM.RichardsonR.D-cycloserine facilitates extinction the first time but not the second time: an examination of the role of NMDA across the course of repeated extinction sessions.Neuropsychopharmacology.2008333096310218354389
- Werner-SeidlerA.RichardsonR.Effects of D-cycloserine on extinction: consequences of prior exposure to imipramine.Biol Psychiatry.2007101195119717555721
- ParnasAS.WeberM.RichardsonR.Effects of multiple exposures to dcycloserine on extinction of conditioned fear in rats.Neurobiol Learn Mem.20058322423115820858
- KalischR.HoltB.PetrovicP.et al.The NMDA agonist D-cycloserine facilitates fear memory consolidation in humans.Cereb Cortex.20091918719618477687
- NageswaraRao K.EidusL.EvansC.et al.Deterioration of cycloserine in the tropics.World Health Organization.196839781789
- MonahanJB.HandelmannGE.HoodWF.CordiAA.D-cycloserine, a positive modulator of the N-methyl-D-aspartate receptor, enhances performance of learning tasks in rats.Pharmacol Biochem Behav.1989346496532560209
- LandC.RiccioDC.d-Cycloserine: effects on long-term retention of a conditioned response and on memory for contextual attributes.Neurobiol Learn Mem.19997215816810536095
- SteeleRJ.DermonCR.StewartMG.D-cycloserine causes transient enhancement of memory for a weak aversive stimulus in day-old chicks (Gallus domesticus).Neurobiol Learn Mem.1996662362408946416
- FloodJF.MorleyJE.LanthornTH.Effect on memory processing by Dcycloserine, an agonist of the NMDA/glycine receptor.Eur J Pharmacol.19922212492541330624
- DubrovinaNI.ZinovyevDR.ZinovyevaDV.N-methyl-D-aspartate receptors and amnesia in mice with depression-like state.Bull Exp Biol Med.200714464764918683485
- QuartermainD.MowerJ.RaffertyMF.HertingRL.LanthornTH.Acute but not chronic activation of the NMDA-coupled glycine receptor with Dcycloserine facilitates learning and retention.Eur J Pharmacol.19942577128082709
- AssiniFL.DuzzioniM.TakahashiRN.Object location memory in mice: pharmacological validation and further evidence of hippocampal CA1 participation.Behav Brain Res.200920420621119523494
- LandCL.RiccioDC.D-Cycloserine, a positive modulator of the NMDA receptor enhances acquisition of a conditioned taste aversion.Psychobiology.199725210216
- NunninkM.DavenportRA.OrtegaB.HouptTA.D-Cycloserine enhances conditioned taste aversion learning in rats.Pharmacol Biochem Behav.20078732133017561237
- MatsuokaN.AignerTG.D-cycloserine, a partial agonist at the glycine site coupled to N-methyl-D-aspartate receptors, improves visual recognition memory in rhesus monkeys.J Pharmacol Exp Ther.19962788918978768744
- ThompsonLT.MoskalJr.DisterhofttJF.Hippocampus-dependent learning facilitated by a monoclonal antibody or D-cycloserine.Nature.19923596386411406995
- BaxterMG.LanthornTH.FrickKM.GolskiS.WanRQ.OltonDS.Dcycloserine, a novel cognitive enhancer, improves spatial memory in aged rats.Neurobiol Aging.1994152072137838293
- ThompsonLT.DisterhoftJF.Age- and dose-dependent facilitation of associative eyeblink conditioning by D-cycloserine in rabbits.Behav Neurosci.1997111130313129438799
- FishkinRJ.InceES.CarlezonWA Jr.DunnRW.D-cycloserine attenuates scopolamine-induced learning and memory deficits in rats.Behav Neural Biol.1993591501578476382
- OhnoM.WatanabeS.D-cycloserine, a glycine site agonist, reverses working memory failure by hippocampal muscarinic receptor blockade in rats.Eur J Pharmacol.19963182672719016914
- ZajaczkowskiW.DanyszW.Effects of D-cycloserine and aniracetam on spatial learning in rats with entorhinal cortex lesions.Pharmacol Biochem Behav.19975621298981605
- TempleMD.HammRJ.Chronic, post-injury administration of Dcycloserine, an NMDA partial agonist, enhances cognitive performance following experimental brain injury.Brain Res.19967412462519001729
- SchusterGM.SchmidtWJ.D-cycloserine reverses the working memory impairment of hippocampal-lesioned rats in a spatial learning task.Eur J Pharmacol.199222497981451747
- MauriceT.LockhartBP.SuTP.PrivatA.Reversion of beta 25-35-amyloid peptide-induced amnesia by NMDA receptor-associated glycine site agonists.Brain Res.19967312492538883881
- WlazP.RolinskiZ.CzuczwarSJ.Influence of D-cycloserine on the anticonvulsant activity of phenytoin and carbamazepine against electroconvulsions in mice.Epilepsia.1996376106178681892
- OttoMW.BasdenSL.McHughRK.et al.Effects of D-cycloserine administration on weekly nonemotional memory tasks in healthy participants.Psychother Psychosom.200978495419018157
- D'SouzaDC.GilR.CasselloK.et al.IV glycine and oral D-cycloserine effects on plasma and CSF amino acids in healthy humans.Biol Psychiatry.20004745046210704956
- KuriyamaK.HonmaM.KoyamaS.KimY.D-cycloserine facilitates procedural learning but not declarative learning in healthy humans: a randomized controlled trial of the effect of d-cycloserine and valproic acid on overnight properties in the performance of non-emotional memory tasks.Neurobiol Learn Mem.20119550550921402164
- OnurOA.SchlaepferTE.KukoljaJ.et al.The N-methyl-D-aspartate receptor co-agonist D-cycloserine facilitates declarative learning and hippocampal activity in humans.Biol Psychiatry.2010671205121120303474
- GoffDC.GatherC.GottliebJD.et al.Once-weekly D-cycloserine effects on negative symptoms and cognition in schizophrenia: an exploratory study.Schizophr Res.200810632032718799288
- DavisM.ResslerK.RothbaumBO.RichardsonR.Effects of D-cycloserine on extinction: translation from preclinical to clinical work.Biol Psychiatry.20066036937516919524