
Abstract
Background
Diet and nutrition can impact on the biological processes underpinning neuropsychiatric disorders. Amino acid (AA) mixtures lacking a specific neurotransmitter precursor can change the levels of brain serotonin (5-HT) or dopamine (DA) in the central nervous system. The availability of these substances within the brain is determined by the blood–brain barrier (BBB) that restricts the access of peripheral AA into the brain. AA mixtures lacking tryptophan (TRP) compete with endogenous TRP for uptake into the brain across the BBB, which in turn leads to a decrease in central nervous 5-HT synthesis.
Objective
The present study compared the effects of a simplified acute tryptophan depletion (SATD) mixture in mice on blood and brain serotonergic and dopaminergic metabolites to those of a commonly used acute tryptophan depletion mixture (ATD Moja-De) and its TRP-balanced control (BAL).
Design
The SATD formula is composed of only three large neutral AAs: phenylalanine (PHE), leucine (LEU), and isoleucine (ILE). BAL, ATD Moja-De, or SATD formulas were delivered to adult male C57BL/6J mice by gavage. TRP, monoamines, and their metabolites were quantified in blood and brain regions (hippocampus, frontal cortex, amygdala, caudate putamen, and nucleus accumbens).
Results
Both ATD Moja-De and SATD significantly decreased levels of serum and brain TRP, as well as brain 5-HIAA and 5-HT compared with BAL. SATD reduced HVA levels in caudate but did not alter total DA levels or DOPAC. SATD decreased TRP and serotonergic metabolites comparably to ATD Moja-De administration.
Conclusion
A simplified and more palatable combination of AAs can manipulate serotonergic function and might be useful to reveal underlying monoamine-related mechanisms contributing to different neuropsychiatric disorders.
Dietary amino acid (AA) depletion mixtures have been used in neuropsychiatric research (Citation1) to study the underlying role of monoamine neurotransmitters (such as serotonin [5-HT], dopamine [DA], or norepinephrine [NE]) in behavioral and psychiatric disorders (i.e. depression), and also (Citation2) as potential nutritional challenge strategies for these disorders (Citation1, Citation2). The underlying methodological factors that determine the neurochemical response to these particular AA mixtures have been discussed, and two major areas of current interest are the development of simplified mixtures that achieve the same results with greater tolerability as well as the investigation of specific neural mechanisms mediating behavioral changes associated with these treatments. The specific mixture composition is one variable which can influence the consequences of these challenge procedures on neurotransmitter synthesis in the brain (Citation3–Citation7).
Biogenic amines (indoleamines, catecholamines, and histamine) are produced from only three essential AAs: tryptophan (TRP), tyrosine (TYR), and histidine (HIS). From embryonal and fetal development, these neurotransmitters control and regulate various vital functions. Changes in monoamine neurotransmission can modify neural function at multiple levels, such as growth and development at early stages and sexual development during maturity (Citation8, Citation9). Thus, when serum TRP is increased by a higher intake through diet, an immediate boost effect is produced on synthesis of 5-HT in the enterochromaffin cells of the GI tract and in the brain. On the contrary, when TRP intake is lower with diet, a depletion effect on peripheral and central 5-HT will be produced (Citation7). These effects are measurable even during fetal and neonatal development (Citation8).
Lowering the concentration of specific AAs that serve as precursors for neurotransmitters in the brain to transiently lower synthesis and release of the neurotransmitter under investigation can also be used to elucidate behavioral and neurobiological effects of the neurotransmitter in question (Citation10–Citation12). This method has been particularly effective when studying 5-HT, as tryptophan hydroxylase 2 (TPH-2, the rate-limiting enzyme for central nervous synthesis of 5-HT) is not saturated at circulating levels of TRP and therefore manipulating TRP availability significantly influences 5-HT synthesis and release (Citation13). Although less effective, a similar strategy has shown some success in transiently lowering availability of TYR and PHE for catecholamine synthesis (Citation2, Citation14–Citation17).
The basic mechanism underpinning this strategy is that large neutral AAs (LNAAs) compete with each other for transport across the blood–brain barrier (BBB) using a shared transporter (LAT-1), and so changing the ratio of competing AAs (CAAs) can limit the entry of precursor AAs into the brain (Citation18, Citation19). Thus, access of each CAA to the brain depends not only on its serum levels but also on the serum concentration of the other LNAAs competing for transport. Phenylalanine (PHE) and leucine (LEU) have a lower Km for transport across the BBB than TRP (Citation20). These particular AAs should provide optimal competition with TRP for transport into the brain. Alteration of peripheral protein synthesis after AA administration may also contribute to lowering brain TRP availability, since administration of an AA mixture stimulates protein synthesis in the liver, depleting plasma TRP, and thus favoring entry of CAAs with consequent reduction of brain TRP (Citation21). Moreover, branch-chained amino acids (BCAAs) and LEU, in particular, may stimulate skeletal muscle protein synthesis (Citation22).
Much effort had been made to identify the optimal composition of these dietary AA-based depletion formulations. Young et al. (Citation23) adopted a composition based on the human milk profile, containing up to 15 AAs (both essential and non-essential AAs). This formulation was widely accepted and used in many studies in the past. However, this very complex mixture presents some disadvantages, including nauseating taste due to the sulfur-containing AAs and low specificity of the targets due to a high concentration of BCAAs (Citation24). The proportions of the complex mixtures were subsequently modified by other groups (Citation25–Citation30) to optimize depletion. The core features of these dietary mixtures formed by the neurotransmitter precursors (TRP, PHE, and TYR) and BCAAs (valine [VAL], leucine [LEU], and isoleucine [ILE]) have been never altered. However, Badawy et al. (Citation24, Citation31) postulated that decreasing the content of BCAAs might improve target specificity. Many of these investigators have modified the AA profile of their formulations or studied the use of proteins such as gelatin or α-lactalbumin (Citation32–Citation35).
We have previously shown that a modified version of a dietary ATD protocol termed Moja-De that minimizes content of sulfur-containing AAs causes less nausea and discomfort in people but decreases 5-HT synthesis in a mouse model as much as the originally suggested formulation (Citation36). While this new dietary mixture offers an advantage in human studies, it is still quite complex in its formulation () and potentially includes off-target responses due to large neutral AAs. In the present study, we tested the hypothesis that providing a simplified dietary AA formula (SATD, Simplified Acute Tryptophan Depletion) which includes just three AAs to compete at LAT-1 with TRP, will subsequently lower central nervous 5-HT synthesis. We investigated variations in serum and various brain regions of TRP and 5-HT metabolite contents, as well as TYR and DA and their metabolite levels and compared them with a control condition formula (BAL) as well as a positive control (ATD Moja-De).
Table 1 Quantities of amino acids for each mixture
Materials and methods
Animals
Adult male mice (8–9 weeks old) from Jackson Laboratories (Bar Harbor, ME, USA) were housed in groups of eight per cage in an approved animal facility under controlled temperature (21±1°C) and humidity with a 12-h light–dark cycle (lights on at 6 a.m.). The animals were allowed to acclimate for 1 week upon arrival; food (LabDiet®) and water were freely provided until the night prior to the experiment, when food was removed. Mice were handled daily to habituate to handling. On the day of testing, food-deprived mice received an AA mixture (BAL, ATD, or SATD) by gavage. The animals were anesthetized with isoflurane and killed by decapitation 2.5 h after the administration of the corresponding formula. The Animal Care and Use Committee at Duke University Medical Center approved this experimental protocol.
The new AA formula
The present study introduces a simplified dietary acute tryptophan depletion formula (SATD), which is based on the previously reported ATD Moja-De mixture (Citation29, Citation36), but is composed of only three LNAAs: PHE, LEU, and ILE. These AAs compete with TRP for access into the brain and thereby provide a TRP-free mixture of LNAAs that lowers central nervous system 5-HT synthesis. To provide adequate concentrations of CAAs, the total amount of each AA was increased relative to the original formula. SATD administration was compared with a control mixture containing a complex mixture of amino acids (BAL), and to the Moja-De ATD modification of the TRP-free mixture. shows the quantities of the components of the different dietary mixtures. The AA mixtures were prepared by the Pharmacy of the University Hospital of RWTH Aachen University (Germany). They were mixed in deionized water on the morning of the experiment, and polytron and sonication bath were used until the formula was completely suspended. The AA mixtures were administered in two doses spaced 30 min apart by gavage with 2 g/kg BW mixed with 6.66 mL/kg BW deionized water, for a total dose of 4 g/kg.
Blood and tissue preparation
Blood samples were collected by cardiac puncture and spun at 16,000×g and 4°C for 20 min. Aliquots of the supernatant were frozen at −80°C until the day of analysis by high-performance liquid chromatography (HPLC). Samples were thawed on the day of assay and 10 µL of serum was diluted with 990 µL of ice-cold standard buffer (0.5 mM sodium metabisulfite, 0.2 N perchloric acid and 0.5 mM EDTA) and spun at 16,000×g and 4°C for 10 min.
Different brain regions (hippocampus, frontal cortex, amygdala, caudate putamen, and nucleus accumbens) were dissected on ice using a mouse brain block, weighed, placed in 1.5 mL Eppendorf tubes and immediately frozen at −80°C. To quantitate the AAs TRP and TYR, as well as the monoamines and their metabolites, brain samples were homogenized in 250 µL of ice-cold standard buffer, except for the caudate samples that homogenized in 500 µL of standard buffer due to the high DA content. The whole homogenates were disrupted by sonication and centrifuged at 16,000×g and 4°C for 20 min. The supernatants were collected and kept on ice, until analysis.
Quantifying the analytes
To analyze the AAs TRP and TYR, a BAS LC-4B apparatus with an electrochemical detector with dual 3 mm carbon electrode (MF-1000) and reference electrode (MF-2021) was used. TRP separation was conducted on a C18 column (Phenomenex® Kinetex™ 2.6 µm, 100×4.6 mm, 100 Å) using the following mobile phase: 8% acetonitrile (v/v), 0.05 M citric acid, 0.05 M, Na2HPO4·7H2O and 0.1 mM EDTA and a flow rate of 1.0 mL/min. Mobile phase was not corrected for pH. Samples were quantified by electrochemical detection with the detector set to 0.85 V versus Ag/AgCl reference electrode and sensitivity at 200 nA. TYR separation was performed on the same type of column with a mobile phase comprising the following: 12% methanol (v/v), 0.1 M, NaH2PO4·7H2O, 0.1 mM EDTA and 2.7 mM octanesulfonic acid (anhydrous); pH 3.8 and a flow of 1.2 mL/min. For electrochemical detection, the detector was set to 0.80 V, sensitivity at 950 nA.
Dopaminergic (DA, DOPAC, and HVA) and serotonergic (5-HT and 5-HIAA) contents were determined using a Constametric® 4100 apparatus, C18 column (Phenomenex® Kinetex™ 2.6 µm, 100×4.6 mm, 100 Å and an electrochemical detector with dual 3 mm carbon electrode (MF-1000) and reference electrode (MF-2021). The mobile phase used for separating these metabolites consisted of: 18% methanol (v/v), 0.1 M sodium phosphate, 0.8 mM octanesulfonic acid (anhydrous), and 0.1 mM EDTA; pH 3.1. Flow rate at 0.7 mL/min. Detector was set to 0.70 V and sensitivity at 50 nA.
Statistical analysis
Statistical evaluation of the data was carried out using NCSS 2007 software (NCSS LLC, Kaysville, UT, USA). Effects of treatment for every dependent measure (TRP, TYR, 5-HT, 5-HIAA, DA, DOPAC and HVA) were analyzed for all groups (BAL, ATD, SATD) by a repeated measures global ANOVA with ‘treatment’ as a between factor and ‘serum’ or ‘brain region’ as a within factor. Analyses that yielded significant main effects and/or interactions were subjected to a lower-order two-way ANOVA and significant two-way interactions were followed by Fisher's LSD multiple-comparison post-hoc testing with the level of significance set at p<0.05. All data are shown as mean±S.E.M. (). Outliers were determined by a GRUBBS statistical test.
Table 2 Contents of TRP, TYR, serotonergic (5-HT and 5-HIAA), and dopaminergic (DA, DOPAC and HVA) metabolites in the serum and different brain regions of mice
Results
SATD affects TRP content in serum
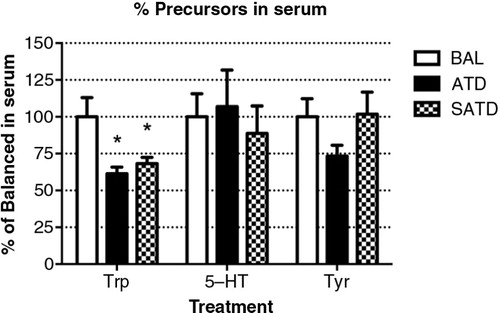
The contents of the AAs TRP and TYR and the neurotransmitter 5-HT were measured in serum of male C57BL/6J mice treated with BAL, ATD, or SATD (). Animals treated with either ATD or SATD showed significantly decreased serum TRP content relative to controls (F [2,23]=6.85, p=0.005). However, no treatment effect of ATD or SATD on serum TYR was detected (F [2,23]=1.86, p=0.181). Circulating 5-HT concentrations were not significantly different between either SATD or ATD treatments versus BAL administration (F [2,23]=0.04, p=0.965).
Fig. 1 Serum levels of tryptophan (TRP), serotonin (5-HT), and tyrosine (TYR) in the mouse after formula administration. Data are represented as mean±S.E.M. Groups of 7–8 mice received either a control condition (BAL), acute tryptophan depletion (ATD), or simplified acute tryptophan depletion (SATD) mixtures. *p<0.05 compared with BAL.
Robust reductions in TRP and serotonergic metabolite levels in brain tissue
A significant decrease of brain TRP levels was shown after SATD and ATD administration (F [2,109]=13.98, p<0.001; F [4,109]=12.97, p<0.0001, for an effect of treatment and region, respectively). A global ANOVA showed a main effect of treatment, region, and treatment×region on brain 5-HT and its metabolite 5-HIAA (Treatment: F [2,108]=41.11, p<0.0001; F [2,108]=50.44, p<0.0001; region: F [4,108]=65.79, p<0.0001; F [4,108]=211.50, p<0.0001; treatment×region: F [8,108]=2.40, p=0.023; F [8,108]=20.23, p<0.0001, respectively). Fisher's post-hoc tests showed the reductions in the brain 5-HT and 5-HIAA contents of mice after dietary SATD or ATD treatment relative to control condition (see – for a summary). While brain 5-HT concentrations were reduced by 20–35% after dietary administration of both SATD and ATD () when compared with BAL, 5-HIAA exhibited a significantly higher decline (around 40–55%, p<0.0001). Globally, dietary administration of the new SATD mixture caused greater 5-HT-related depletion magnitude when compared with the ATD Moja-De formula ().
Fig. 2 Levels of tryptophan in the different brain regions of the mouse after formula administration. Data are represented as mean±S.E.M. Groups of 7–8 mice received either a control condition (BAL), acute tryptophan depletion (ATD), or simplified acute tryptophan depletion (SATD) mixtures. HPC: hippocampus; FC: frontal cortex; Amyg: amygdala; CPu: caudate putamen; NAcc: nucleus accumbens. *p<0.05 compared with BAL.
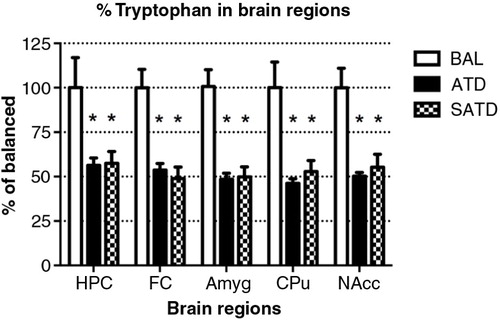
Fig. 3 Levels of serotonin (5-HT) in the different brain regions of the mouse after formula administration. Data are represented as mean±S.E.M. Groups of 7–8 mice received either a control condition (BAL), acute tryptophan depletion (ATD), or simplified acute tryptophan depletion (SATD) mixtures. HPC: hippocampus; FC: frontal cortex; Amyg: amygdala; CPu: caudate putamen; NAcc: nucleus accumbens. *p<0.05 compared with BAL.
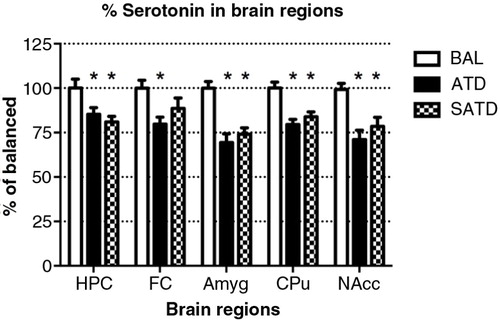
Fig. 4 Levels of 5-hydroxyindoleacetic acid (5-HIAA) in the different brain regions of the mouse after formula administration. Data are represented as mean±S.E.M. Groups of 7–8 mice received either a control condition (BAL), acute tryptophan depletion (ATD), or simplified acute tryptophan depletion (SATD) mixtures. HPC: hippocampus; FC: frontal cortex; Amyg: amygdala; CPu: caudate putamen; NAcc: nucleus accumbens. *p<0.05 compared with BAL.
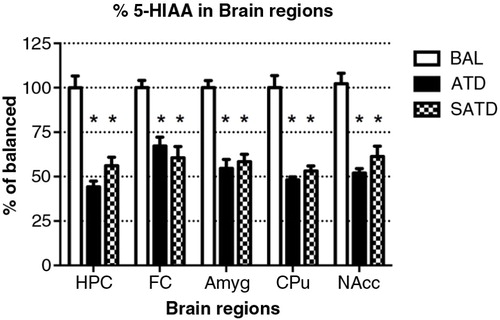
Table 3 This is a summary of the effects of the treatments relative to balanced mixture (BAL)
Brain HVA decreased after administration of ATD or SATD
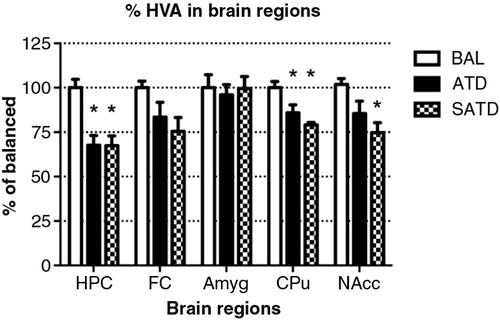
There was no main effect of treatment on brain TYR, DA, or DOPAC (F [2,109]=2.78, p=0.087; F [2,106]=2.06, p=0.154; F [2,106]=0.24, p=0.788, respectively, and ). However, a global ANOVA indicated an effect of region in both analytes (TYR: F [4,109]=31.64, p<0.0001; DA: F [4,108]=518.73, p<0.0001; DOPAC: F [4,108]=302.21, p<0.0001). A lower-order ANOVA and Fisher's post-hoc tests indicated that the dopaminergic metabolite DOPAC in the amygdala of SATD-treated mice was significantly higher relative to BAL-treated mice (F [2,21]=4.91, p=0.020). HVA showed a significant effect of treatment, region and treatment×region (Treatment: F [2,107]=6.29, p=0.008; region: F [4,107]=1349.35, p<0.0001; treatment×region: F [8,107]=7.23, p<0.001). The levels of HVA were affected (25–35% depletion, ) in hippocampus, caudate, and nucleus accumbens when SATD was administered. ATD administration reduced HVA content in hippocampus and caudate ().
Fig. 5 Levels of tyrosine in the different brain regions of the mouse after formula administration. Data are represented as mean±S.E.M. Groups of 7–8 mice received either a control condition (BAL), acute tryptophan depletion (ATD), or simplified acute tryptophan depletion (SATD) mixtures. HPC: hippocampus; FC: frontal cortex; Amyg: amygdala; CPu: caudate putamen; NAcc: nucleus accumbens.
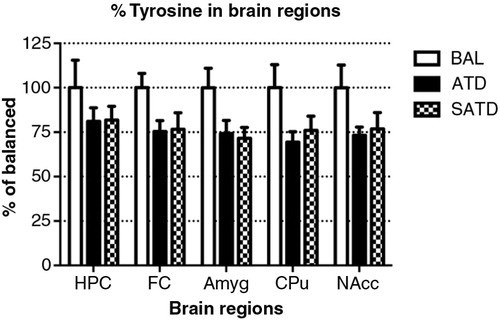
Fig. 6 Levels of dopamine (DA) in the different brain regions of the mouse after formula administration. Data are represented as mean±S.E.M. Groups of 7–8 mice received either a control condition (BAL), acute tryptophan depletion (ATD), or simplified acute tryptophan depletion (SATD) mixtures. HPC: hippocampus; FC: frontal cortex; Amyg: amygdala; CPu: caudate putamen; NAcc: nucleus accumbens.
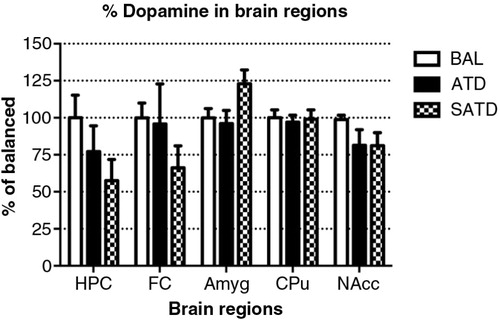
Fig. 7 Levels of homovanillic acid (HVA) in the different brain regions of the mouse after formula administration. Data are represented as mean±S.E.M. Groups of 7–8 mice received either a control condition (BAL), acute tryptophan depletion (ATD), or simplified acute tryptophan depletion (SATD) mixtures. HPC: hippocampus; FC: frontal cortex; Amyg: amygdala; CPu: caudate putamen; NAcc: nucleus accumbens. *p<0.05 compared with BAL.
Effect on 5-HT and DA turnovers
5-HT turnover (5-HIAA/5-HT ratio) presented a significant effect of treatment (F [2,108]=11.17, p<0.001), region (F [4,108]=62.51, p<0.0001), and a treatment×region interaction (F [8,108]=9.87, p<0.0001, ). These values showed a reduced ratio after dietary SATD and ATD administration when compared with the BAL condition in the hippocampus, caudate, and nucleus accumbens. Moreover, DA turnover (HVA/DA ratio) did not exhibit a main effect of treatment but of region (F [2,106]=1.45, p=0.260; F [4,106]=10.78, p<0.001, respectively). An increase in this ratio was achieved with SATD and ATD treatments in the hippocampus and frontal cortex compared with other regions, as indicated by a lower-order ANOVA and Fisher's post-hoc tests.
Discussion
Previous studies from our group have demonstrated that the ATD formula Moja-De or combined monoamine depletion mixture (CMD) successfully reduce TRP and serotonergic metabolite levels in blood and brain tissues (Citation29, Citation36). The present study demonstrates the efficacy of a simplified dietary formulation in affecting serum and different brain AA and neurotransmitter metabolite contents in a mouse model. These findings suggest that SATD, which is composed of only three essential AAs (PHE, LEU, and ILE), can compete with TRP at the BBB sufficiently to significantly impair its transport. SATD induced a robust decrease in serum and brain TRP levels of male C57BL/6J mice compared with control condition (BAL) and these results are in line with those obtained with ATD. Moreover, significant reductions in brain 5-HT and its metabolite 5-HIAA caused after SATD administration were equivalent to those induced by ATD Moja-De relative to BAL () and greater than those induced by CMD in our previous study (Citation29). SATD reduced the final product of this pathway (HVA); however, no changes were observed either on the AA precursor TYR nor the subsequent molecules DA or DOPAC. One explanation for this finding could be that DA turnover was affected indirectly by altered serotonergic turnover. These results support the potential utility of this simplified combination of AAs to examine the neurochemical underpinnings of different neuropsychiatric disorders.
The AA content of the new dietary SATD mixture is dramatically simplified from the original Moja-De ATD (Citation28). The AA composition was reduced from 7 to 3 essential AAs (compensated by an increased amount of each). PHE, LEU and ILE were selected for this formulation based on their reported affinity for the transporter LAT-1 and concentration in the plasma to provide optimal competition for TRP transport into the central nervous system (Citation37, Citation38). It is unlikely that peripheral effects on protein synthesis are the critical determinants for successful TRP depletion in the brain and successful transient inhibition of serotonergic function. While it will be important to investigate the neurochemical outcome of this mixture further, the present findings suggest that this simpler formulation may present two advantages for effecting TRP depletion in studies focusing on serotonergic effects on behavior and disease, in particular because a simpler mixture is available that may be better tolerated by humans, and a mixture with fewer off-target effects due to the lack of LNAAs.
A drawback of this new mixture might be the higher total dose of AAs used in this particular experiment, which may lead to a reduced tolerability in humans. High doses of AAs have been used with conventional ATD mixtures, some applying up to 100 g of AAs in one dose (Citation23). However, since these mixtures contained a different composition of AAs, and the observed clinical effects as well as side effects cannot be directly compared. To optimize the dosing regimen, a comparison of different doses should be conducted to find an optimal dose–response relationship before testing this mixture in humans and to allow for the smallest effective dose possible and thereby minimize possible side and adverse effects.
Another possible pitfall of this novel formulation may be its main attribute, namely the reduced number of AAs used. PHE, which is the essential AA precursor of tyrosine and comprises a relative larger ratio of SATD as compared with conventional ATD mixtures, might increase DA synthesis. It has been used as a challenge test in neurological research in a diagnostic setting (Citation39, Citation40). Patients with dopa-responsive dystonia (DRD), who have a significantly different PHE/TYR ratio as compared with other dystonic patients, showed an improvement of symptoms after a PHE-challenge with 100 mg PHE/kg bodyweight (Citation39, Citation40). Taking into account that the amount of PHE applied with the SATD is relatively larger than the conventional ATD mixtures, and the amount of AA to compete with PHE's uptake across the BBB is relatively small, one possible result could be an increase in dopaminergic synthesis in some brain areas. This aspect of SATD needs to be further investigated, ideally in brain areas typically densely populated with dopaminergic neurons.
Another consideration is that although this mixture is simplified, it contains BCAA, which contributes to off-target effects. BCAA may contribute to the change in DA and possibly glutamate after TRP depletion, and behavioral consequences of these changes (Citation24, Citation31) (Citation41, Citation42). In addition, BCAA have both central and peripheral effects on glucose and protein metabolism that can indirectly change brain function (Citation43).
Dietary TRP depletion affects mood, behavior and cognition in many species (Citation26, Citation28) (Citation29, Citation36) (Citation44). Its great value is that changes are most notable in specific vulnerable patient populations (Citation12, Citation27) (Citation45). Although TRP depletion does not lower mood in most healthy individuals, volunteer participants – in particular women – with a past history of depression or bipolar illness or family history of depression can exhibit decreased mood after dietary TRP depletion (Citation12, Citation46–Citation48). More recent research has shown that changes in emotional and cognitive processing occur when even small changes in central nervous system TRP levels occur in vulnerable populations (Citation30, Citation49). However, many formulations are complex to prepare, and nauseating to subjects. The present results have identified a dietary formulation (SATD) with a very limited number of AAs (only three of them) that achieved a level of depletion previously obtained with a standard ATD protocol. Such combinations may provide a useful tool for future studies of neurochemical mechanisms related to the neurobiological underpinnings of neuropsychiatric disorders in terms of a neurodietary approach.
The use of AA-based depletion paradigms is also of clinical relevance. One possible target could be seen in the use of ATD protocols with regard to manic symptoms, which are at least in part thought to be related to a hyper-serotonergic state (Citation50–Citation52). For instance, one double-blind, placebo-controlled pilot study using ATD in acutely manic patients showed that patients who ATD in combination with sodium valproate showed greater improvement in mania ratings (Citation53). However, a high intolerance rate limited the use of the used depletion paradigm. However, with regard to recent research using more refined and body weight–adapted depletion paradigms such as the Moja-De ATD protocol tolerability was at acceptable rates (Citation54, Citation55), and neurodietary depletion strategies such as the ATD technique can provide important information on brain function and the neurochemical modulation of neurocircuitries in patients with psychiatric disorders and healthy people (Citation53–Citation57).
In conclusion, the present study has successfully demonstrated the action of a new simplified dietary AA formulation as regards impacting brain 5-HT content in the same way as the previously used ATD Moja-De depletion protocol (with a more complex AA-related content) does.
Authors’ contributions
CLS, FDZ, and CMK designed the experiments, interpreted the data, and wrote the paper. CLS performed experiments, collected, and analyzed the data. CLS, AEDV, and AEA gathered blood and brain data. CSB wrote complementary information.
Conflict of interest and funding
CLS, AEDV, AEA, CSB, and CMK have no conflict of interest.
FDZ was the recipient of an unrestricted award donated by the American Psychiatric Association (APA), the American Psychiatric Institute for Research and Education (APIRE) and AstraZeneca (Young Minds in Psychiatry Award). He has also received research support from the German Federal Ministry for Economics and Technology, the European Union (EU), the German Society for Social Pediatrics and Adolescent Medicine, the Paul and Ursula Klein Foundation, the Dr. August Scheidel Foundation, the IZKF fund of the University Hospital of RWTH Aachen University, and a travel stipend donated by the GlaxoSmithKline Foundation. He is the recipient of an unrestricted educational grant, travel support and speaker honoraria by Shire Pharmaceuticals, Germany. In addition, he has received support from the Raine Foundation for Medical Research (Raine Visiting Professorship), and editorial fees from Co-Action Publishing (Sweden).
Acknowledgements
This work received funding from the Bundesministerium für Wirtschaft und Technologie (BMWi).
Related Research Data
References
- Bell CJ, Hood SD, Nutt DJ. Acute tryptophan depletion. Part II: clinical effects and implications. Aust N Z J Psychiatry. 2005; 39: 565–74. [PubMed Abstract].
- Harmer CJ, McTavish SFB, Clark L, Goodwin GM, Cowen PJ. Tyrosine depletion attenuates dopamine function in healthy volunteers. Psychopharmacology. 2001; 154: 105–11. [PubMed Abstract].
- Badawy A. Novel nutritional treatment for manic and psychotic disorders: a review of tryptophan and tyrosine depletion studies and the potential of protein-based formulations using glycomacropeptide. Psychopharmacology. 2013; 228: 347–58. [PubMed Abstract].
- Young SN. Acute tryptophan depletion in humans: a review of theoretical, practical and ethical aspects. J Psychiatry Neurosci. 2013; 38: 294–305. [PubMed Abstract] [PubMed CentralFull Text].
- Young SN. Appropriate amino acid mixtures for tryptophan depletion and tyrosine/phenylalanine depletion and the safety of long-term amino acid depletion in humans. Psychopharmacology. 2013; 229: 377–8. [PubMed Abstract].
- Young SN. Methodology and interpretation of acute tryptophan depletion studies. Acta Psychiatr Scand. 2014; 129: 156. [PubMed Abstract].
- Zepf FD, Sánchez CL, Biskup CS, Kötting WF, Bubenzer S, Helmbold K, etal. Acute tryptophan depletion – converging evidence for decreasing central nervous serotonin synthesis in rodents and humans. Acta Psychiatr Scand. 2014; 129: 157–9. [PubMed Abstract].
- Castrogiovanni P, Musumeci G, Trovato FM, Avola R, Magro G, Imbesi R. Effects of high-tryptophan diet on pre- and postnatal development in rats: a morphological study. Eur J Nutr. 2014; 53: 297–308.
- Musumeci G, Trovato FM, Avola R, Imbesi R, Castrogiovanni P. Serotonin/growth hormone/insulin-like growth factors axis on pre- and post-natal development: a contemporary review. OA Anatomy. 2013; 1: 12.
- Wurtman RJ, Fernstrom JD. Control of brain neurotransmitter synthesis by precursor availability and nutritional state. Biochem Pharmacol. 1976; 25: 1691–6.
- Fernstrom JD. Effects on the diet on brain neurotransmitters. Metabolism. 1977; 26: 207–23.
- Booij L, Van der Does AJ, Riedel WJ. Monoamine depletion in psychiatric and healthy populations: review. Mol Psychiatry. 2003; 8: 951–73.
- Fadda F, Cocco S, Stancampiano R. A physiological method to selectively decrease brain serotonin release. Brain Res Protoc. 2000; 5: 219–22.
- Bongiovanni R, Kyser AN, Jaskiw GE. Tyrosine depletion lowers in vivo DOPA synthesis in ventral hippocampus. Eur J Nutr. 2012; 696: 70–6.
- Palmour RM, Ervin FR, Baker GB, Young SN. An amino acid mixture deficient in phenylalanine and tyrosine reduces cerebrospinal fluid catecholamine metabolites and alcohol consumption in vervet monkeys. Psychopharmacology. 1998; 136: 1–7.
- McTavish SF, Cowen PJ, Sharp T. Effect of a tyrosine-free amino acid mixture on regional brain catecholamine synthesis and release. Psychopharmacology. 1999; 141: 182–8.
- Sheehan BD, Tharyan P, McTavish SF, Campling GM, Cowen PJ. Use of a dietary manipulation to deplete plasma tyrosine and phenylalanine in healthy subjects. J Psychopharmacol. 1996; 10: 231–4.
- Pardridge WM. Blood-brain barrier carrier-mediated transport and brain metabolism of amino acids. Neurochem Res. 1998; 23: 635–44.
- Jakeman PM. Amino acid metabolism, branched-chain amino acid feeding and brain monoamine function. Proc Nutr Soc. 1998; 57: 35–41.
- Pardridge WM. Kinetics of competitive inhibition of neutral amino acid transport across the blood-brain barrier. J Neurochem. 1977; 28: 103–8.
- Hood SD, Bell CJ, Nutt DJ. Acute tryptophan depletion. Part I: rationale and methodology. Aust N Z J Psychiatry. 2005; 39: 558–64.
- Anthony TG, McDaniel BJ, Knoll P, Bunpo P, Paul GL, McNurlan MA. Feeding meals containing soy or whey protein after exercise stimulates protein synthesis and translation initiation in the skeletal muscle of male rats. J Nutr. 2007; 137: 357–62.
- Young SN, Smith SE, Pihp RO, Ervin FR. Psychopharmacology Tryptophan depletion causes a rapid lowering of mood in normal males. Psychopharmacology. 1985; 87: 173–7.
- Badawy AA, Dougherty DM, Richard DM. Specificity of the acute tryptophan and tyrosine plus phenylalanine depletion and loading tests part II: normalisation of the tryptophan and the tyrosine plus phenylalanine to competing amino acid ratios in a new control formulation. Int J Tryptophan Res. 2010; 3: 35–47. [PubMed Abstract] [PubMed CentralFull Text].
- Biggio G, Fadda F, Fanni P, Tagliamonte A, Gessa GL. Rapid depletion of serum tryptophan, brain tryptophan, serotonin and 5-hyfroxyindoleacetic acid by a tryptophan-free diet. Life Sci. 1974; 14: 1321–9.
- Delgado P, Price L, Miller H, Salomon R, Aghajanian G, Heninger G, etal. Serotonin and the neurobiology of depression. Effects of tryptophan depletion in drug-free depressed patients. Arch Gen Psychiatry. 1994; 51: 865–74.
- Hayward G, Goodwin GM, Cowen PJ, Harmer CJ. Low-dose tryptophan depletion in recovered depressed patients induces changes in cognitive processing without depressive symptoms. Biol Psychiatry. 2005; 57: 517–24. [PubMed Abstract].
- Moja EA, Stoff DM, Gessa GL, Castoldi D, Assereto R, Tofanetti O. Decrease in plasma tryptophan after tryptophan-free amino acid mixtures in man. Life Sci. 1988; 42: 1551–6.
- Sánchez CL, Van Swearingen AED, Arrant AE, Kuhn CM, Zepf FD. Dietary manipulation of serotonergic and dopaminergic function in C57BL/6J mice with amino acid depletion mixtures. J Neural Transm. 2014; 121: 153–62.
- Dingerkus VLS, Gaber TJ, Helmbold K, Bubenzer S, Eisert A, Sánchez CL, etal. Acute tryptophan depletion in accordance with body weight: influx of amino acids across the blood–brain barrier. J Neural Transm. 2012; 119: 1037–45. [PubMed Abstract] [PubMed CentralFull Text].
- Badawy AA, Dougherty DM, Richard DM. Specificity of the acute tryptophan and tyrosine plus phenylalanine depletion and loading tests I. Review of biochemical aspects and poor specificity of current amino Acid formulations. Int J Tryptophan Res. 2010; 2010: 23–34. [PubMed Abstract].
- Jans LAW, Lieben CKJ, Smits LT, Blokland A. Pharmacokinetics of acute tryptophan depletion using a gelatin-based protein in male and female Wistar rats. Amino Acids. 2009; 37: 349–57. [PubMed Abstract] [PubMed CentralFull Text].
- Choi S, DiSilvio B, Fernstrom MH, Fernstrom JD. The chronic ingestion of diets containing different proteins produces marked variations in brain tryptophan levels and serotonin synthesis in the rat. Neurochem Res. 2011; 36: 559–65. [PubMed Abstract].
- Choi S, Disilvio B, Fernstrom MH, Fernstrom JD. Meal ingestion, amino acids and brain neurotransmitters: effects of dietary protein source on serotonin and catecholamine synthesis rates. Physiol Behav. 2009; 98: 156–62. [PubMed Abstract].
- Lieben C. Acute tryptophan and serotonin depletion using an optimized tryptophan-free protein–carbohydrate mixture in the adult rat. Neurochem Int. 2004; 44: 9–16. [PubMed Abstract].
- Biskup CS, Sánchez CL, Arrant A, Van Swearingen AED, Kuhn C, Zepf FD. Effects of acute tryptophan depletion on brain serotonin function and concentrations of dopamine and norepinephrine in C57BL/6J and BALB/cJ mice. PLoS One. 2012; 7: 35916.
- Gijsman HJ, Scarnà A, Harmer CJ, McTavish SB, Odontiadis J, Cowen PJ, etal. A dose-finding study on the effects of branch chain amino acids on surrogate markers of brain dopamine function. Psychopharmacology. 2002; 160: 192–7. [PubMed Abstract].
- Scarnà A, Gijsman HJ, Harmer CJ, Goodwin GM, Cowen PJ. Effect of branch chain amino acids supplemented with tryptophan on tyrosine availability and plasma prolactin. Psychopharmacology. 2002; 159: 222–3.
- Opladen T, Hoffmann GF, Kühn AA, Blau N. Pitfalls in phenylalanine loading test in the diagnosis of dopa-responsive dystonia. Mol Genet Metab. 2013; 108: 195–7. [PubMed Abstract].
- Bandmann O, Goertz M, Zschocke J, Deuschl G, Jost W, Hefter H, etal. The phenylalanine loading test in the differential diagnosis of dystonia. Neurology. 2003; 60: 700–2. [PubMed Abstract].
- Fernstrom JD. Large neutral amino acids: dietary effects on brain neurochemistry and function. Amino Acids. 2013. 45: 419–30.
- Le Masurier M, Oldenzeil W, Lehman C, Cowen P, Sharp T. Effect of acute tyrosine depletion in using a branched chain amino-acid mixture on dopamine neurotransmission in the rat brain. Neuropsychopharmacology. 2006; 31: 310–17.
- Murín R, Hamprecht B. Metabolic and regulatory roles of leucine in neural cells. Neurochem Res. 2008; 33: 279–84.
- Young SN, Ervin FR, Pihl RO, Finn P. Biochemical aspects of tryptophan depletion in primates. Psychopharmacology. 1989; 98: 508–11. [PubMed Abstract].
- Bell C. Tryptophan depletion and its implications for psychiatry. Br J Psychiatry. 2001; 178: 399–405. [PubMed Abstract].
- Ellenbogen MA, Young SN, Dean P, Palmour RM, Benkelfat C. Mood response to acute tryptophan depletion in healthy volunteers: sex differences and temporal stability. Neuropsychopharmacology. 1996; 15: 465–74.
- Klaassen T, Riedel WJ, van Someren A, Deutz NE, Honig A, van Praag HM. Mood effects of 24-hour tryptophan depletion in healthy first-degree relatives of patients with affective disorders. Biol Psychiatry. 1999; 46: 489–97.
- Feder A, Skipper J, Blair JR, Buchholz K, Mathew SJ, Schwarz M, etal. Tryptophan depletion and emotional processing in healthy volunteers at high risk for depression. Biol Psychiatry. 2011; 69: 804–7. [PubMed Abstract] [PubMed CentralFull Text].
- Van der Veen FM, Evers EAT, Deutz NEP, Schmitt JAJ. Effects of acute tryptophan depletion on mood and facial emotion perception related brain activation and performance in healthy women with and without a family history of depression. Neuropsychopharmacology. 2007; 32: 216–24.
- Applebaum J, Bersudsky Y, Klein E. Rapid tryptophan depletion as a treatment for acute mania: a double-blind, pilot-controlled study. Bipolar Disord. 2007; 9: 884–7.
- Zepf FD, Wöckel L, Herpertz-Dahlmann B, Poustka F, Freitag CM. Tryptophan depletion in bipolar mania: reduction of metabolic imbalance by administration according to body weight?. Bipolar Disord. 2009; 11: 557–8. [PubMed Abstract].
- Zepf FD, Wöckel L, Poustka F, Holtmann M. Dietary tryptophan depletion according to body weight – A new treatment option in acute mania?. Med Hypotheses. 2009; 72: 47–8. [PubMed Abstract].
- Helmbold K, Zvyagintsev M, Dahmen B, Bubenzer-Busch S, Gaber TJ, Crockett MJ, etal. Effects of serotonin depletion on punishment processing in the orbitofrontal and anterior cingulate cortices of healthy women. Eur Neuropsychopharmacol. 2015; 25: 846–56. [PubMed Abstract].
- Hildebrand P, Konigschulte W, Gaber TJ, Bubenzer-busch S, Helmbold K, Biskup CS, etal. Effects of dietary tryptophan and phenylalanine–tyrosine depletion on phasic alertness in healthy adults – A pilot study. Food Nutr Res. 2015; 59: 26407.
- Von Polier GG, Biskup CS, Kötting WF, Bubenzer S, Helmbold K, Eisert A, etal. Change in electrodermal activity after acute tryptophan depletion associated with aggression in young people with attention deficit hyperactivity disorder (ADHD). J Neural Transm. 2014; 121: 451–5. [PubMed Abstract].
- Sánchez CL, Biskup CS, Herpertz-Dahlmann B, Gaber TJ, Kuhn CM, Hood SD, etal. The role of serotonin (5-HT) in behavioral control: findings from animal research and clinical implications. Int J Neuropsychopharmacol. 2015; 1–13. doi: http://dx.doi.org/10.1093/ijnp/pyv050 [Epub ahead of print]..
- Zepf FD, Dingerkus VLS, Helmbold K, Bubenzer-Busch S, Biskup CS, Herpertz-Dahlmann B, etal. Effects of a short-term reduction in brain serotonin synthesis on the availability of the soluble leptin receptor in healthy women. J Neural Transm. 2015; 122: 343–8. [PubMed Abstract].