
Abstract
Tryptophan is an essential amino acid with an indole nucleus. Humans cannot produce this amino acid themselves, but must obtain it through their diet. Much attention is currently paid to the wide physiological and clinical implications of the tryptophan-derived substances, serotonin and kynurenines, generated by human enzymes following the intestinal absorption of tryptophan. However, even before being absorbed, several microbial metabolites of tryptophan are formed, mainly from ‘malabsorbed’ (incompletely digested) proteins within the colon. The normal smell of human faeces is largely due to indole, one of the major metabolites. Recent studies indicate that this foul-smelling substance is also of utmost importance for our health.
The gastrointestinal tract is rich in microbe-generated signal substances that affect several biological functions in the body (‘inter-kingdom communication’) and regulate the growth and functioning of the complex microbial ecosystem (‘intra-kingdom communication’, ‘quorum sensing’). The chemical ‘language’ used in the communication is extremely old in a phylogenetic context, although well preserved and developed in evolutionary terms. Its constituents were described already in the early 1980s (Citation1), although the concept of ‘microbial endocrinology’ is of a more recent origin (Citation2). Today, it is considered possible that alterations of the microbiome (‘dysbiosis’) may be a major cause not only of irritable bowel syndrome, fibromyalgia, and chronic fatigue syndrome but also a number of other common diseases such as obesity, diabetes mellitus, atherosclerosis, and allergic and autoimmune diseases (Citation3, Citation4). Indole is an example of a microbe-generated signal substance that has positive effects on its host as well as the microbiome, and normal-smelling faeces may be an underestimated health indicator.
Indole: a tryptophan metabolite
In the intestine, tryptophan can be metabolised by bacteria to form a number of metabolically active substances. Under experimental conditions, the amount of microbe-generated indole is related to the amount of exogenously supplied tryptophan (Citation5). In humans, however, free tryptophan is nearly completely absorbed within the small intestine, and it is uncertain how much of it is transformed to indole (by the intestinal bacteria) when tryptophan is provided in its pure form (Citation6). Single doses of 100 mg per kg body weight of pure tryptophan appear to have no adverse effects. No changes of the urinary excretion of 5-hydroxyindoleacetic acid (the end product of serotonin metabolism) or of indican (the end product of indole metabolism) indicate that such a fairly large dose is within the limit of what the body can absorb and metabolise. Normally, however, the intestinal microbiota is supplied with tryptophan in the form of proteins in our food, and the anaerobic bacteria in the colon, in particular, produce indole from undigested proteins, a decomposition process that involves the enzyme tryptophanase.
Escherichia coli is a typical indole producer, and indole produced by E. coli functions as a ‘quorum-sensing’ signal that regulates the virulence and biofilm formation of both E. coli and other bacteria. Indole is, for example, important for the pathogenicity of enterohaemorrhagic E. coli (EHEC); while indole itself inhibits biofilm formation of EHEC, oxidised indole stimulates it (Citation7). While indole decreases E. coli biofilms, it increases those of pseudomonads (Citation8) and the growth of the cholera bacterium (Vibrio cholerae; (Citation9)). Perhaps for this reason V. cholerae thrives in the colon. In contrast, V. anguillarum, a bacterium that causes major damage in aquaculture facilities, loses its virulence when exposed to indole and stays clear of indole-contaminated water (Citation10). In humans, microbial tryptophanase and indole are essential for our defence against intestinal worms and the regulation of the plasmid content of microbes (Citation11, Citation12).
In general, indole has a beneficial effect on the host's intestinal function. Irritable bowel syndrome patients have a weakened mucosal barrier, and endogenous serotonin cannot repair the problem (Citation13). Thus, it is interesting to notice that indole strengthens the barrier function of the mucous membrane by repairing ‘tight junctions’, both in vitro in human epithelial cell cultures (Citation14) and in vivo in bacteria-free mice (Citation15). However, no studies have yet clearly shown whether indole also can strengthen the mucosal barrier in vivo in humans. The indole-based signal system can be manipulated with the aid of drugs, dietary changes, and other bacteria, and recent research aims to exploit these signal systems. A relative shortage of Faecalibacterium prausnitzii in the faecal flora appears to be a reproducible finding in patients with inflammatory bowel disease (IBD; (Citation16, Citation17)). Therefore, it is worth noting that culture supernatants of this strictly anaerobic, indole-producing bacterium strengthens the mucosal barrier and protects against experimental colitis in mice (Citation18, Citation19). Also, odorants present in the luminal environment of the gut may stimulate serotonin release via olfactory receptors present in human enterochromaffin cells. Olfactory receptors are, therefore, potential novel targets for the treatment of gastrointestinal motility disorders (Citation20, Citation21). Other receptors may be involved when the enterochromaffin cells secrete serotonin and glucagon-like peptide-1 (GLP-1) in response to short-chain fatty acids and indole, respectively (Citation22, Citation23).
A recent study shows that microbes that produce indole-like substances may have beneficial effects even in persons with alcohol addiction and increased intestinal permeability. In these persons, increased permeability is associated with the craving for alcohol as well as mental problems, such as anxiety and depression. Interestingly, when the permeability improves after abstinence, it may serve as a predictor for a concomitant reduction in the craving for alcohol and the likelihood of relapse of drinking (Citation24).
Hydrogen peroxide
Apparently, indole plays a key role in the orchestration of several microbial processes in the bowel and for the functions of the epithelial cells in the intestine, but the underlying mechanisms are largely unknown. Indole is obviously not the only signal substance that ensures microbial homeostasis in the bowel; it is involved in the interaction with numerous other substances. It is tempting to speculate whether indole works through activation of the peroxisome proliferator-activated receptor (PPAR)-γ, since this is an endogenous regulator of intestinal inflammation, and a lack of this ligand-activated transcription factor is associated with chronic inflammation, such as IBD (Citation25). Interestingly, indole-based PPAR agonists are developed, albeit not for this purpose (Citation26).
Hydrogen peroxide (H2O2)-producing lactobacilli can provide important homeostatic support to indole, because H2O2 activates PPAR-γ. In contrast to strictly anaerobic bacteria, the lactobacilli are not killed by H2O2 or O2, and they can thus find their niches in aerobic pockets in close contact with the intestinal epithelium (Citation27). Although lactobacilli produce no catalase, they have a high tolerance of H2O2 and it is conceivable that they produce sufficient amounts of H2O2 in close contact with the epithelium to ensure continuous activation of PPAR-γ.
The redox potential in the alimentary tract goes from being positive in the proximal sections of the small intestine to being negative in the large bowel. In the proximal sections of the intestine, the microbes will therefore use O2 as an electron acceptor, while the facultative anaerobes, which can use other electron acceptors [e.g. nitrate, nitrite, thiocyanate, sulphate, and trimethylaminoxide (TMAO)], become more dominant as the O2 tension recedes in the distal direction. The redox system thus promotes different floras in the small and large intestines. The redox system also helps in regulating intestinal permeability, immune defence, gene expression, healing of wounds, and stem cell proliferation (Citation28). It is, thus, obvious that all processes that may interfere with the redox values can affect the composition of the bacterial flora and the function of the epithelial cells in the intestine. For example, if the redox potential in the large intestine is insufficiently negative (free of O2), this may have an effect on the anaerobic bacteria contained within it. It is conceivable that a lack of strictly anaerobic, indole-producing bacteria (for example F. prausnitzii in the case of IBD) may be the result of an insufficiently negative redox potential distally in the bowel. A sufficiently negative redox potential (an O2-free environment) may also prevent oxidative conversion of indole to hydroxyindole derivates that do not have the same beneficial effect on bowel health as indole.
Hydrogen peroxide can also be produced by the enzyme ‘dual oxidase’ (DUOX) within the epithelial cells in the small intestine and the large bowel. DUOX activity is induced by bacteria, and most likely has an effect on the redox potential locally and clearance of other microbes (Citation28). However, whereas H2O2 production from lactobacilli adherent to the mucosal epithelium may have a preventive effect against intestinal diseases, H2O2 production from DUOX may be a part of an immune response.
Indoleamine dioxygenase
The two enzymes, tryptophan 2,3-dioxygenase (TDO) and indoleamine 2,3-dioxygenase (IDO), catabolise (decyclizate) tryptophan to N-formylkynurenine. Activation of innate immunity with the release of interferon-γ (INF γ) stimulates IDO. The result is the depletion of tryptophan and production of metabolites of the kynurenine pathway, which both influence the normal flora and intestinal permeability. In addition, the production of metabolites can provide a source of nicotinamide dinucleotide (NAD+; (Citation29)), and superoxide radicals produced during IDO-catalysed tryptophan oxidation will generate highly reactive peroxynitrite in the presence of NO. IDO is present in many cells in the bowel, especially antigen-presenting cells such as macrophages and dendritic cells in the Peyer's patches in the ileum are strongly IDO positive (Citation30). Studies have shown that IDO is inhibited (oxidised) by hydrogen peroxide and that this could be a method whereby lactic acid bacteria influence mucosal barrier and immunity (Citation31).
Melatonin
Melatonin, N-acetyl-5-methoxytryptamine, is a highly conserved indoleamine found in bacteria, plants, and animals. Its production by photosynthetic bacteria show that melatonin as well as indole are both probably phylogenetically very ancient as signal substances, possibly developed to protect aerobic organisms against the toxic effects of oxygen (Citation32). Melatonin produced by the pineal gland is known as our major regulator of the ‘internal clock’. The fact that melatonin may also originate from the gastrointestinal tract was first reported in 1976, when it was discovered that serotonin-rich enterochromaffin cells produce melatonin (Citation33). In 1992, it was shown in rats that oral administration of tryptophan, the precursor of melatonin, causes a rapid elevation of circulating melatonin, reaching much higher levels than those recorded after parenteral administration of this amino acid (Citation34). The total amounts of melatonin produced daily within the gastrointestinal tract may be about 500 times higher than those secreted by the pineal gland (Citation35). Melatonin from the enterochromaffin cells is released into the portal vein and metabolised by the liver. The metabolites are excreted in bile and urine. Intriguingly, urinary excretion is abnormally low in patients with irritable bowel syndrome, suggesting altered melatonin production or metabolism in this group of patients (Citation36).
Melatonin acts as a direct free radical scavenger in vitro. In vivo, it may reduce oxidative damage of excessive free radical generation (Citation32, Citation37). Consistently, it attenuates lipopolysaccharide-induced hepatotoxicity (Citation38) and stress-induced gastrointestinal motility disorder and gastric stress ulcers in rats (Citation39), and melatonin or its precursor tryptophan may be effective in the treatment of oesophagitis and gastroduodenal ulcers in human beings (Citation40). Chronic fatigue syndrome is associated with high oxidative stress levels (Citation41) and unexplained abdominal complaints (Citation42), and several studies indicate that the administration of melatonin may improve abdominal pain in these patients (Citation43).
Having a common precursor (tryptophan), the production of both serotonin and melatonin is affected by the availability of tryptophan. Together with GLP-1, another signalling substance from the enteroendocrine cells, they play important roles in gastrointestinal physiology, including immunological, inflammatory, and oxidative defence (Citation44). Whether gut microbes promote their production through effects of indole or short-chain fatty acids is an interesting possibility (Citation22, Citation23).
Indole: an archetypical hormone?
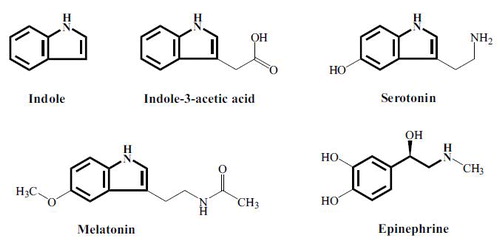
The indole nucleus, found in human hormones such as serotonin and melatonin (; 8), is also found in the most important growth-regulating hormone in plants, indoleacetic acid (IAA). Indole formed from tryptophan by microbes in the bowel regulates growth, virulence, and formation of biofilm within this ecosystem (Citation8), while also regulating the functioning of the intestinal epithelium favourably. This is a striking parallel to what happens in the interaction between plants and soil microbes. Indole produced by microbes in intimate contact with plant roots regulates microbial growth in this ecosystem (the rhizosphere), while indole also stimulates the development of the plants’ root system and intervenes in the regulation of plant growth and development (Citation45). Plants can ‘smell’ indole and react positively to it even in volatile form. Therefore, the role of indole in plants supports the idea of indole as a very ancient, archetypical hormone, which evolution has preserved to help regulate the interaction between microbes and their host organisms in the vegetable as well as in the animal kingdom.
Fig. 1 Indole as an archetypical hormone. The indole nucleus is the basis for a number of important signal molecules in both plants and animals. Reproduced with permission from (8).
Manure: a valuable material
Microbe-rich and indole-smelling manure is obviously a valuable material. Plants communicate with bacteria with the aid of indole (Citation45), and we have all seen that manure may reinvigorate ailing plants. Perhaps, it is unsurprising that patients who ‘wither’ may also recover when supplied with fresh faeces that have the right smell (Citation46)?
Conflict of interest and funding
The authors have not received any funding or benefits from industry or elsewhere to conduct this study.
Related Research Data
References
- Roth J , LeRoith D , Shiloach J , Rosenzweig JL , Lesniak MA , Havrankova J . The evolutionary origins of hormones, neurotransmitters, and other extracellular chemical messengers: implications for mammalian biology. N Engl J Med. 1982; 306: 523–7.
- Lyte M . Microbial endocrinology and the microbiota–gut–brain axis. Adv Exp Med Biol. 2014; 817: 3–24.
- Berstad A , Raa J , Valeur J . Tryptophan: ‘essential’ for the pathogenesis of irritable bowel syndrome?. Scand J Gastroenterol. 2014; 49: 1493–8.
- Tremaroli V , Backhed F . Functional interactions between the gut microbiota and host metabolism. Nature. 2012; 489: 242–9.
- Li G , Young KD . Indole production by the tryptophanase TnaA in Escherichia coli is determined by the amount of exogenous tryptophan. Microbiology. 2013; 159(Pt 2): 402–10.
- Michael AF , Drummond KN , Doeden D , Anderson JA , Good RA . Tryptophan metabolism in man. J Clin Invest. 1964; 43: 1730–46.
- Lee J , Bansal T , Jayaraman A , Bentley WE , Wood TK . Enterohemorrhagic Escherichia coli biofilms are inhibited by 7-hydroxyindole and stimulated by isatin. Appl Environ Microbiol. 2007; 73: 4100–9.
- Lee J , Jayaraman A , Wood TK . Indole is an inter-species biofilm signal mediated by SdiA. BMC Microbiol. 2007; 7: 42.
- Mueller RS , Beyhan S , Saini SG , Yildiz FH , Bartlett DH . Indole acts as an extracellular cue regulating gene expression in Vibrio cholerae. J Bacteriol. 2009; 191: 3504–16.
- Li X , Yang Q , Dierckens K , Milton DL , Defoirdt T . RpoS and indole signaling control the virulence of Vibrio anguillarum towards gnotobiotic sea bass (Dicentrarchus labrax) larvae. PLoS One. 2014; 9: e111801.
- Anyanful A , Dolan-Livengood JM , Lewis T , Sheth S , Dezalia MN , Sherman MA , etal. Paralysis and killing of Caenorhabditis elegans by enteropathogenic Escherichia coli requires the bacterial tryptophanase gene. Mol Microbiol. 2005; 57: 988–1007.
- Chant EL , Summers DK . Indole signalling contributes to the stable maintenance of Escherichia coli multicopy plasmids. Mol Microbiol. 2007; 63: 35–43.
- Keszthelyi D , Troost FJ , Jonkers DM , van Eijk HM , Lindsey PJ , Dekker J , etal. Serotonergic reinforcement of intestinal barrier function is impaired in irritable bowel syndrome. Aliment Pharmacol Ther. 2014; 40: 392–402.
- Bansal T , Alaniz RC , Wood TK , Jayaraman A . The bacterial signal indole increases epithelial-cell tight-junction resistance and attenuates indicators of inflammation. Proc Natl Acad Sci USA. 2010; 107: 228–33.
- Shimada Y , Kinoshita M , Harada K , Mizutani M , Masahata K , Kayama H , etal. Commensal bacteria-dependent indole production enhances epithelial barrier function in the colon. PLoS One. 2013; 8: e80604.
- Cao Y , Shen J , Ran ZH . Association between Faecalibacterium prausnitzii reduction and inflammatory bowel disease: a meta-analysis and systematic review of the literature. Gastroenterol Res Pract. 2014; 2014: 872725.
- Sokol H , Pigneur B , Watterlot L , Lakhdari O , Bermudez-Humaran LG . Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc Natl Acad Sci USA. 2008; 105: 16731–6.
- Martin R , Chain F , Miquel S , Lu J , Gratadoux JJ , Sokol H , etal. The commensal bacterium Faecalibacterium prausnitzii is protective in DNBS-induced chronic moderate and severe colitis models. Inflamm Bowel Dis. 2014; 20: 417–30.
- Carlsson AH , Yakymenko O , Olivier I , Hakansson F , Postma E , Keita AV , etal. Faecalibacterium prausnitzii supernatant improves intestinal barrier function in mice DSS colitis. Scand J Gastroenterol. 2013; 48: 1136–44.
- Kidd M , Modlin IM , Gustafsson BI , Drozdov I , Hauso O , Pfragner R . Luminal regulation of normal and neoplastic human EC cell serotonin release is mediated by bile salts, amines, tastants, and olfactants. Am J Physiol Gastrointest Liver Physiol. 2008; 295: G260–72.
- Braun T , Voland P , Kunz L , Prinz C , Gratzl M . Enterochromaffin cells of the human gut: sensors for spices and odorants. Gastroenterology. 2007; 132: 1890–901.
- Chimerel C , Emery E , Summers DK , Keyser U , Gribble FM , Reimann F . Bacterial metabolite indole modulates incretin secretion from intestinal enteroendocrine L cells. Cell Rep. 2014; 9: 1202–8.
- Reigstad CS , Salmonson CE , Rainey JF III , Szurszewski JH , Linden DR , Sonnenburg JL , etal. Gut microbes promote colonic serotonin production through an effect of short-chain fatty acids on enterochromaffin cells. FASEB J. 2015; 29: 1395–403.
- Leclercq S , Matamoros S , Cani PD , Neyrinck AM , Jamar F , Starkel P , etal. Intestinal permeability, gut-bacterial dysbiosis, and behavioral markers of alcohol-dependence severity. Proc Natl Acad Sci USA. 2014; 111: E4485–93.
- Voltan S , Martines D , Elli M , Brun P , Longo S , Porzionato A , etal. Lactobacillus crispatus M247-derived H2O2 acts as a signal transducing molecule activating peroxisome proliferator activated receptor-gamma in the intestinal mucosa. Gastroenterology. 2008; 135: 1216–27.
- Gim HJ , Li H , Jeong JH , Lee SJ , Sung MK , Song MY , etal. Design, synthesis, and biological evaluation of a series of alkoxy-3-indolylacetic acids as peroxisome proliferator-activated receptor gamma/delta agonists. Bioorg Med Chem. 2015; 23: 3322–36.
- Hertzberger R , Arents J , Dekker HL , Pridmore RD , Gysler C , Kleerebezem M , etal. H(2)O(2) production in species of the Lactobacillus acidophilus group: a central role for a novel NADH-dependent flavin reductase. Appl Environ Microbiol. 2014; 80: 2229–39.
- Kim SH , Lee WJ . Role of DUOX in gut inflammation: lessons from Drosophila model of gut-microbiota interactions. Front Cell Infect Microbiol. 2014; 3: 116.
- Ball HJ , Jusof FF , Bakmiwewa SM , Hunt NH , Yuasa HJ . Tryptophan-catabolizing enzymes – party of three. Front Immunol. 2014; 5: 485.
- Dai X , Zhu BT . Indoleamine 2,3-dioxygenase tissue distribution and cellular localization in mice: implications for its biological functions. J Histochem Cytochem. 2010; 58: 17–28.
- Valladares R , Bojilova L , Potts AH , Cameron E , Gardner C , Lorca G , etal. Lactobacillus johnsonii inhibits indoleamine 2,3-dioxygenase and alters tryptophan metabolite levels in BioBreeding rats. FASEB J. 2013; 27: 1711–20.
- Manchester LC , Poeggeler B , Alvares FL , Ogden GB , Reiter RJ . Melatonin immunoreactivity in the photosynthetic prokaryote Rhodospirillum rubrum: implications for an ancient antioxidant system. Cell Mol Biol Res. 1995; 41: 391–5.
- Raikhlin NT , Kvetnoy IM . Melatonin and enterochromaffine cells. Acta Histochem. 1976; 55: 19–24.
- Huether G , Poeggeler B , Reimer A , George A . Effect of tryptophan administration on circulating melatonin levels in chicks and rats: evidence for stimulation of melatonin synthesis and release in the gastrointestinal tract. Life Sci. 1992; 51: 945–53.
- Celinski K , Konturek PC , Slomka M , Cichoz-Lach H , Gonciarz M , Bielanski W , etal. Altered basal and postprandial plasma melatonin, gastrin, ghrelin, leptin and insulin in patients with liver cirrhosis and portal hypertension without and with oral administration of melatonin or tryptophan. J Pineal Res. 2009; 46: 408–14.
- Radwan P , Skrzydlo-Radomanska B , Radwan-Kwiatek K , Burak-Czapiuk B , Strzemecka J . Is melatonin involved in the irritable bowel syndrome?. J Physiol Pharmacol. 2009; 60(Suppl 3): 67–70.
- Reiter RJ , Tan DX , Manchester LC , Qi W . Biochemical reactivity of melatonin with reactive oxygen and nitrogen species: a review of the evidence. Cell Biochem Biophys. 2001; 34: 237–56.
- Sewerynek E , Melchiorri D , Reiter RJ , Ortiz GG , Lewinski A . Lipopolysaccharide-induced hepatotoxicity is inhibited by the antioxidant melatonin. Eur J Pharmacol. 1995; 293: 327–34.
- Zhang L , Gong JT , Zhang HQ , Song QH , Xu GH , Cai L , etal. Melatonin attenuates noise stress-induced gastrointestinal motility disorder and gastric stress ulcer: role of gastrointestinal hormones and oxidative stress in rats. J Neurogastroenterol Motil. 2015; 21: 189–99.
- Celinski K , Konturek SJ , Konturek PC , Brzozowski T , Cichoz-Lach H , Slomka M , etal. Melatonin or L-tryptophan accelerates healing of gastroduodenal ulcers in patients treated with omeprazole. J Pineal Res. 2011; 50: 389–94.
- Kennedy G , Spence VA , McLaren M , Hill A , Underwood C , Belch JJ . Oxidative stress levels are raised in chronic fatigue syndrome and are associated with clinical symptoms. Free Radic Biol Med. 2005; 39: 584–9.
- Berstad A , Undseth R , Lind R , Valeur J . Functional bowel symptoms, fibromyalgia and fatigue: a food-related triad?. Scand J Gastroenterol. 2012; 47: 914–19.
- Song GH , Leng PH , Gwee KA , Moochhala SM , Ho KY . Melatonin improves abdominal pain in irritable bowel syndrome patients who have sleep disturbances: a randomised, double blind, placebo controlled study. Gut. 2005; 54: 1402–7.
- Khan WI , Ghia JE . Gut hormones: emerging role in immune activation and inflammation. Clin Exp Immunol. 2010; 161: 19–27.
- Bailly A , Groenhagen U , Schulz S , Geisler M , Eberl L , Weisskopf L . The inter-kingdom volatile signal indole promotes root development by interfering with auxin signalling. Plant J. 2014; 80: 758–71.
- van Nood E , Vrieze A , Nieuwdorp M , Fuentes S , Zoetendal EG , de Vos WM , etal. Duodenal infusion of donor feces for recurrent Clostridium difficile . N Engl J Med. 2013; 368: 407–15.