
Abstract
In areas isolated from direct human impact, such as Spitsbergen, environmental changes result mainly from natural processes, which include nutrient enrichment caused by seabirds. The objective of this study was to evaluate the degree of nutrient enrichment of organic soils fertilized by seabirds, indicated by the phosphorus content and transformations. This study encompassed two areas on the Bellsund coast. A profile without the influence of seabirds and profiles located at different distances from the colony of birds (0–150 m) were analysed. A sequential phosphorus fractionation method was used, and three inorganic P (Pi) fractions were obtained as a result: Pi-L (labile P), Pi-FeAl (P associated with Fe and Al) and Pi-CaMg (P associated with Ca and Mg); and two fractions of organic P (Po): Po-HuAc (P associated with humic acids) and Po-Res (residual P). Polar organic soils not subjected to the direct seabird impact contained amounts of total phosphorus (Pt) similar to organic soils in other climate zones. The presence of the seabird colony increased the Pt content and changed the distribution between the ratio of organic and inorganic P fractions. Within the inorganic P fraction, the Pi–CaMg component was dominant and its distribution was modified by the fertilizing effect of bird droppings. The nutrient enrichment of organic soils by birds in the polar zone was therefore very strong.
The transformations affecting the soil environment of Svalbard result mainly from natural processes on account of the archipelago's remote location. Spitsbergen's soil cover has formed differently than the soil cover of southerly locations, primarily due to its climate and the impact of glaciers. The present soil cover of Spitsbergen has been influenced by humidity, weathering and cryogenic processes in addition to the relief and lithology of the Bedrock (Forman & Miller Citation1984; Mann et al. Citation1986; Klimowicz & Uziak Citation1988; Uziak Citation1992; Blümel et al. Citation1993; Klimowicz et al. Citation1993; Melke & Chodorowski Citation2006; Klimowicz et al. Citation2008).
In very humid tundra, vegetation develops in local depressions of coastal plains or on hill slopes and at the foot of the slopes. The formation of peat soils in these habitats is significantly influenced by seabird colonies that increase the nutrient content in the soils in the coastal zone. Many species of birds foraging in the sea deliver large quantities of faeces rich in macro- and micro-elements directly to the soils in the neighbourhood of their breeding sites (Speir & Cowling Citation1984; Tatur Citation1989, Citation2002; Godzik Citation1991; Headley Citation1996; Anderson & Polis Citation1999; Ligęza & Smal Citation2003; Hawke & Newman Citation2004). Thus, the seabirds directly influence the chemical properties of the soils forming there and, indirectly, the vegetation cover (Hogg & Morton Citation1983; Myrcha & Tatur Citation1991; Mun Citation1997; Breuning-Madsen et al. Citation2008; Mulder et al. Citation2011).
Organic and organic-mineral soils are quite common in the Bellsund area, but they occupy small areas associated with water stagnation and the presence of animals. Peat soil profiles are usually shallow, rarely exceeding 50 cm (Eurola Citation1971; Zelikson Citation1971; Låg Citation1980; Göttlich & Hornburg Citation1982; Klimowicz et al. Citation1997).
The chemical composition of organic soils on Spitsbergen has not been well researched; only a few publications mention it (Dobrovolsky Citation1990; Jóźwik & Magierski Citation1992; Klimowicz et al. Citation1997; Melke Citation1999). The literature also lacks studies mentioning the content of various forms of phosphorus and their mutual relationships in the organic soils of the Arctic area. These data can be obtained by means of phosphorus speciation analysis. Such studies focus on using various patterns of sequential phosphorus fractionation in order to examine the resources and transformations of phosphorus quantitatively and qualitatively (Bedrock et al. Citation1994; Cross & Schlesinger Citation1995, Citation2001; Ivanoff et al. Citation1998; Robinson et al. Citation1998; Schlichting et al. Citation2002; Litaor et al. Citation2004; Condron & Newman Citation2011).
Phosphorus is an essential nutrient for plants because of the role it plays in biochemical processes. Its deficiency in soil and water environments is a factor that limits primary production, and its excess leads to eutrophication of ecosystems (Smil Citation2000; Elser et al. Citation2007). Hence, it determines the fertility of water and land ecosystems. It is relatively stable in the soil, and its secondary accumulation occurs in organic deposits (Dowding et al. Citation1981; Richardson & Marshall Citation1986; Craft & Richardson Citation1993; Wang et al. Citation2012). The determination of total phosphorus (Pt) content as well as the content of its particular forms can help identify fertility changes taking place in the environment.
The objective of this research was to study the content of various forms of phosphorus in organic soils subjected to the varying impacts of the seabirds; determine the distribution of various forms of P in the bulk P pool; and to identify the relationship between the inorganic and organic forms of phosphorus along with their location in the soil profiles. On that basis, an attempt was made to assess the impact of the seabirds on the nutrient enrichment of the soils in the immediate vicinity of a breeding colony.
Study area
The study included two sites located at the southern shore of Bellsund, in the north-western part of Wedel Jarlsberg Land in western Spitsbergen ().
Fig. 1 Location of the study area and survey points. (Map based on Zagórski Citation2005.)
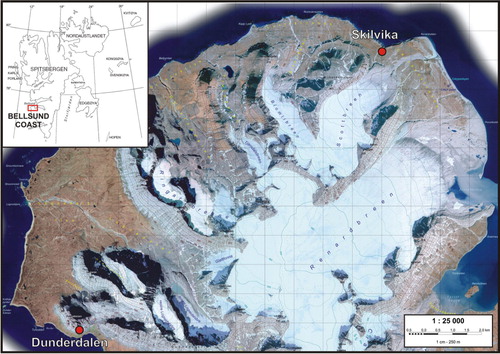
The first site was a reference profile of organic soil not subjected to the direct impact of the seabirds, referred to as the Skilvika profile. It is located within Calypsostranda, on the Skilvika Bay near a seashore cliff (). The site lies within a system of raised coastal terraces whose topographic surface reaches 40–65 m a.s.l. The area where peat occurs is a flat plain dissected by polygons of varying sizes, covered by various moss species and, to a lesser extent, by grass clumps and willow shrubs (Salix polaris).
The other site was the Dunderdalen transect, located on the southern slope of Dundrabeisen, at 30–90 m a.s.l. (). A peat bog of the slope type occurs here, a short distance away from the sea coast, within a talus fan. Above the peat bog, there is a nearly vertical cliff, devoid of vegetation but having hundreds of seabird nests, inhabited mostly by the black-legged kittiwake (Rissa tridactyla). The peat bog under study is small in size (well below 1 ha), not more than 100 m in width; its inclination is about 30°. The entire peat bog is overgrown by various moss species.
Materials and methods
The Skilvika profile was collected from a crevice filled with mossy peat. Such crevices are usually 40–80 cm wide and more than 50 cm deep. Permafrost lies below. The site where the material was collected had similar dimensions and lay on a flat surface fed by precipitation. The peat, along with the mineral substrate, was extracted to a depth of 67 cm and divided into 27 samples (1.5–2.5 cm thick; the mineral substrate sample was 7 cm thick).
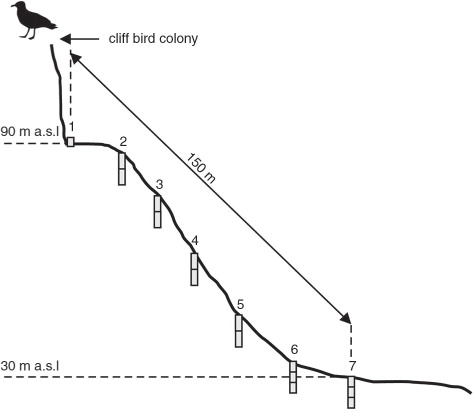
The material in the Dunderdalen area was collected at seven points along a transect from the rock cliff to the foot of the slope (). The first point was a surface sample located immediately underneath the seabird nests, composed of dry plant remains, bird droppings and feathers. Points from 2 to 7 were soil samples, where the distance between points was about 25 m. The last point of transect was 150 m away from the seabird colony. The thickness of the organic horizon varied, but generally did not exceed 0.5 m. The organic horizon was underlain by rock debris of varying degrees of fragmentation. Sampling locations 2–7 consisted of two or three samples (5 or 10 cm thick). A total of 15 samples were obtained from the entire transect.
Fig. 2 The location of sample collection points at the Dunderdalen transect.
For the soil samples collected, a morphological description was prepared (Table , 2 ). After drying and grinding the material, the following were determined: hygroscopic water using the oven-drying method; organic matter and mineral ingredient content using the incandescence method (Sapek & Sapek Citation1997); carbonate content using the Scheibler method (Lityński et al. Citation1976); and pH in 0.01 M CaCl2 using the potentiometric method (Thomas Citation1996).
Table 1 Basic properties and phosphorus sequential extract ion for the Skilvika profile.
Table 2 Basic properties and phosphorus sequential extraction for the Dunderdalen transect.
Phosphorus fractionation was carried out for all samples from the Dunderdalen transect and from 13 samples from the Skilvika profile (selected according to the morphology and basic properties). There is a great variety of methods for determining various forms of phosphorus in mineral soils (Cross & Schlesinger Citation1995). Based on these methods, modifications for organic soils have been developed. One of them is a modification by Reddy et al. (Citation1998) based on the Chang & Jackson method (Citation1957). This sequential phosphorus fractioning method is widely used in American studies for soils in boggy areas (Ivanoff et al. Citation1998; Reddy et al. Citation1998; Ann et al. Citation2000). Its scheme is presented in .
Fig. 3 Scheme used to determine the phosphorus fraction, developed by Reddy et al. Citation1998 (Ziółek Citation2007).
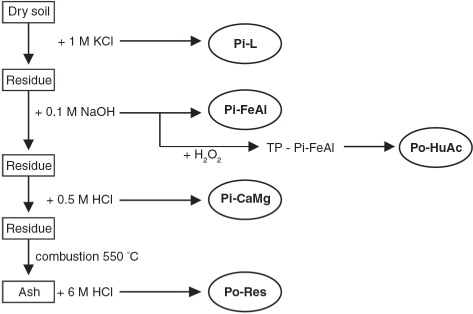
The particular phosphorus fractions were determined by means of the ascorbic acid procedure of Murphy & Riley (Kuo Citation1996), on a Lambda 12 spectrophotometer (Perkin Elmer, Waltham, MA, USA). The total inorganic phosphorus (Pi) was calculated as the sum of the following fractions: labile, easily soluble phosphorus (Pi–L), phosphorus associated with Fe and Al (Pi–FeAl), and inorganic phosphorus associated with Ca and Mg (Pi–CaMg). The total organic phosphorus (Po) is the sum of the phosphorus associated with humic acids (Po–HuAc) and phosphorus in residue fractions (Po–Res). The sum of all forms of phosphorus determined is referred to as Pt.
Results
Soil characteristics
The basic physicochemical properties of the organic soils under study were variable, which resulted from the varying morphological structure, organic matter content and ash content. The organic part of the Skilvika profile (mossy peat of varying degrees of decomposition) reached a depth of 57.5 cm (). It was underlain by a clayey mineral substrate. The organic matter content in the surface part exceeded 80% and decreased with depth. The pH of the studied soil was slightly acidic (with the exception of neutral pH in the sample surface).
In the Dunderdalen peat transect, all the studied samples were organic (organic matter content ranged from 30.5 to 78.3%) overlying rock debris (). The surface samples (0–10 or 0–5 cm in the remaining six sample locations) had a greater organic matter content than those lying deeper. The pH of the studied organic deposits ranged from slightly acidic to basic. The highest pH (7.5) was exhibited by a surface sample located immediately underneath the seabird colony, whereas the lowest pH (5.6) was represented by a sample from the neighbouring point 2. On both sites, the studied deposits did not contain carbonates.
Total P: content and distribution
The soils of the sites under study were characterized by a high Pt content which varied considerably between the two sites (Table , 2 ). Very large quantities of Pt were determined in the Dunderdalen transect (from 1189.5 to 16157.0 mg/kg of dry matter); the greatest quantities occurred in points closest to the breeding seabird colony; these quantities fell as the distance from the colony increased (). Much lower and less varied Pt content (from 899.3 to 1457.6 mg/kg of dry matter) was found in the Skilvika profile (). The smallest quantities were found in surface samples and they increased with depth.
Distribution of average values of P forms
compares the averaged values for all measured P forms from the Dunderdalen transect against the reference of averaged values for all measured P forms from the Silvika profile. Average values of P for Dunderdalen transect were calculated from all samples for all points (depth of 0–20 or 0–25 cm). Average values for Skilvika were obtained from surface samples with corresponding depth (0–20 cm; ).
Fig. 4 Averaged values of P forms for the Dunderdalen transect plotted against Silvika reference values.
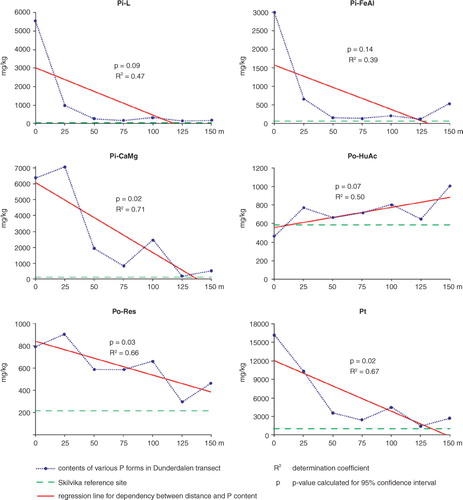
Generally, the content of all P forms in the entire Dunderdalen transect were higher than values obtained for the Skilvika. It means that the fertilizing impact of the bird colony covered 150 m of transect and may have reached even further because of the steep slope. The highest values of Pi–L, Pi–FeAl, Pi–CaMg, Po–Res forms and, accordingly, Pt, occurred near the seabird colony (points 1 and 2 of the transect); they decreased in distance away from it, as shown by a regression line. The Po–HuAc form was an exception as its values increased in distance away from the seabird colony. Calculated determination coefficient R 2 showed that the correlation between the distance from the colony and P contents for the three P forms was strong (R 2 above 0.6). For the rest of the forms there was a weaker correlation between the distance from the colony and P contents (R 2 between 0.4 and 0.5; ). Although the analysis was done for only seven sampling locations, for most of the forms the correlation is explicit.
Proportion of various phosphorus forms in the Pt
Inorganic phosphorus—labile
The contribution of the Pi–L (easily soluble) fraction in Pt varied in the organic soils studied (). For samples from the Dunderdalen transect, the contribution ranged from 2.8 to 34.4%, whereas for the Skilvika profile it ranged from 0.03 to 29.7% (, ). In Dunderdalen, this fraction had the highest contribution in the surface sample below the seabird colony (more than 34%). In the other points of the transect, the percentage contribution of this form was lower, reaching maximum values (about 10%) in surface samples (). In the Skilvika profile, the maximum contribution of the Pi–L fraction also occurred in the surface sample (nearly 30%), and it decreased rapidly with depth (to less than 1%; ).
Fig. 5 Skilvika profile. Proportional participation of different forms of soil P in the total P pool. Terms are abbreviated as follows: labile inorganic P (Pi–L); inorganic P bound with Fe and Al (Pi–FeAl); inorganic P bound with Ca and Mg (Pi–CaMg); organic P associated with humus acids (Po–HuAc); and residual organic P (Po–Res).

Fig. 6 Dunderdalen transect. Proportional participation of different forms of soil P in total P pool. Terms are abbreviated as follows: labile inorganic P (Pi–L); inorganic P bound with Fe and Al (Pi–FeAl); inorganic P bound with Ca and Mg (Pi–CaMg); organic P associated with humus acids (Po–HuAc); and residual organic P (Po–Res).
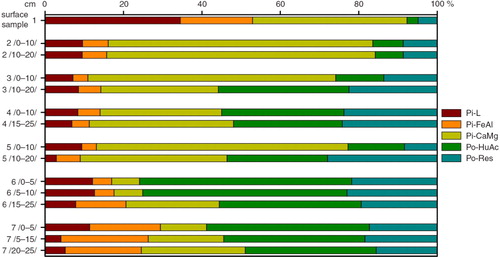
Table 3 Skilvika profile and Dunderdalen transect. The mean and standard deviation (SD) for the percentage contribution of different P forms in the total P pool.
Inorganic phosphorus associated with iron and aluminium
The contribution of the Pi–FeAl fraction to the Pt was low, and minor variation occurred between sites (). In the Skilvika profile this contribution ranged from 4.4 to 10.4% of Pt, reaching the highest value in the surface sample (). For the Dunderdalen transect, the range was from 3.7 to 22.2% of Pt, with a higher contribution in samples from the last point of the transect, and in the surface sample from below the seabird colony ().
Inorganic phosphorus associated with calcium and magnesium
The percentage contribution and distribution of the Pi–CaMg fraction varied considerably between the two sites (). The contribution of this phosphorus form in the Skilvika profile ranged from 3.6 to 36.6% of Pt, and its distribution was characterized by values that increased with depth (). In the Dunderdalen transect, the Pi–CaMg fraction was found to have a considerably greater contribution in the Pt pool, ranging from 7.1 to 68.5% (). The distribution of this form of phosphorus along the transect varied, and was different than in the Skilvika profile; the highest values were found in the samples close to the seabird colony. This form dominated in the total P in points 1, 2 and 3 of the transect.
Total inorganic phosphorus
In the organic soils under study, Pi accounted for 20.4–92.3% of the Pt pool (Table , 2 ). The greatest contribution of Pi, ranging from 24.1 to 92.3% of Pt was found in samples from the Dunderdalen transect (). The highest values of inorganic forms occurred in the surface sample underneath the seabird colony (>92%) and in the samples in point 2 (>83%). In the Skilvika profile, the contribution of Pi forms was smaller and ranged from 20.4 to 43.7% of Pt (). Pi had the highest percentage contribution in the surface sample and the sample from the mineral substrate. Pi–CaMg was the dominant fraction in the total Pi in both sites under study: up to 68.5% of the Pi pool in the Dunderdalen transect, and up to 36.6% of the Pi pool in the Skilvika profile (, ). The labile form of phosphorus, Pi–L, also played a significant role in Pi.
Organic phosphorus associated with humic acids (humic and fulvic)
The organic form Po–HuAc content varied considerably in the organic soils under study (). This fraction had very high percentage contributions in the entire Skilvika profile, ranging from 45.9 to 62.0% of Pt (). In the Dunderdalen transect samples, the contribution of the Po–HuAc form in the Pt pool was lower and more varied, ranging from 2.8 to 54.1% (). The lowest values of this form occurred near the seabird colony where they increased along the transect.
Organic phosphorus in residue
Po content in residue showed a very similar range of values for both sites: for the Skilvika profile they ranged from 6.4 to 27.4% of Pt, whereas for the Dunderdalen transect they ranged from 4.9 to 28.0% of Pt (, , ). In the Skilvika profile, the lowest values, below 10%, occurred in the surface sample and the deepest mineral sample, whereas the maximum values occurred in the upper part of the profile (7–12.5 cm). In the Dunderdalen transect, the lowest percentage contribution of the Po–Res fraction occurred near the seabird colony.
Total organic phosphorus
In the Skilvika profile, the Po pool ranged from 56.3 to 79.6% of Pt; its contribution was quite high and evenly spread throughout the profile (). In the Dunderdalen transect, Po content varied considerably: from 7.7 to 75.9% of Pt (). The lowest value of Po occurred in the upper part of the transect, close to the seabird colony. The Po–HuAc fraction dominated among the Po forms in the soils of Spitsbergen (with the exception of the beginning of the Dunderdalen transect, where the Po–Res form had a greater contribution).
Po/Pi ratio
The ratio of the Po to Pi quantities was low for the soil in the Dunderdalen transect fertilized by seabirds, reaching the lowest values near the seabird colony (from 0.1 to 1.3) (). In the Skilvika profile, not subjected to the direct impact of seabirds, the Po/Pi ratio was considerably higher (from 1.3 to 3.9; ).
Discussion
The organic deposits under study were characterized by high Pt content relative to values reported by other authors for different areas of the world (Parent & Khiari Citation2003). These values were considerably higher than values found for mineral soils, suggesting that the capacity of organic soils to take in and absorb phosphorus is greater than that of mineral soils (Craft & Richardson Citation1993; Reddy et al. Citation1998; Wang et al. Citation2012). There are also studies showing poor retention of P by organic soils (Hawke & Newman Citation2004). The phosphorus content and its distribution in organic soils are closely linked to the overlapping or joint effect of various factors and phenomena occurring during the development of these soils.
The highest concentrations of phosphorus occurred directly below the seabird colony, indicating that seabirds have a crucial impact on the content of phosphorus in soils. The gregarious seabirds foraging in the sea fertilize soils close to their breeding grounds with large quantities of seabird droppings rich in phosphorus compounds (Godzik Citation1991; Myrcha & Tatur Citation1991; Klimowicz et al. Citation1997; Anderson & Polis Citation1999; Ligęza & Smal Citation2003; Breuning-Madsen et al. Citation2008). Phosphorus delivered with seabird droppings are poorly soluble, accumulating intensively in the soil (Comerford Citation1998). However, despite their relatively high capacity to accumulate P, organic soils cannot retain all of the large quantity of P delivered by seabirds, resulting in Pt dispersion. This is demonstrated by a steady decrease in Pt along the transect (). At the Skilvika site, which was not subject to the direct impact of seabirds, the Pt quantities recorded were similar to those characteristic of subalpine peat bogs (Wang et al. Citation2012) as well as organic soils of moderate climate zones not subjected to strong anthropogenic fertilization (Ziółek Citation2007).
An examination of the distribution of Pt at Dunderdalen showed that as one moved away from the seabird colony along the slope, phosphorus content decreased approximately by a factor of 8 (at points 3, 4 and 7 of the transect; ). The fertilization of the soil by seabird droppings in the Dunderdalen transect was the strongest near the seabird nests, where intensive direct accumulation occurred, and decreased with the distance away from the seabird colony. The decrease in Pt content along the slope with a considerable inclination could also result from the gradual accumulation of this element in the upper, drained horizon of organic soil (Schlichting et al. Citation2002; Litaor et al. Citation2004).
The sequential phosphorus fractionation made it possible to discover the relative proportions of the various forms of P in the organic soils of the Bellsund area. The distribution of the Pi pool between its various fractions clearly indicated a significant impact of seabirds on two forms: Pi–L and Pi–CaMg. The very high concentrations of Pi–L near the seabird colony resulted from the direct delivery of seabird droppings rich in this element. The considerable contribution of Pi–L in the upper part of the Skilvika profile could result from remineralization of plant litter. The contribution of the Pi–CaMg form and its distribution in the soils at the study sites indicated a clear relationship with the fertilizing effect of the seabird colony which was probably due to the high content of Ca and Mg in the bird guano (Fugler Citation1985). However the tendency for the Pi–CaMg form to accumulate near the nutrient enrichment (eutrophication) source was also noted in American studies on bog soils subjected to strong fertilization (Reddy et al. Citation1998). The contribution of this form of phosphorus increased with the depth in the Skilvika profile which was not subject to concentrated fertilization. Such a relationship in the distribution of this form was also observed in the peat soils of the moderate zone not subjected to direct fertilization (Ziółek Citation2007). The seabirds seemed not to have an impact on the contribution of Pi–FeAl in the Pt pool. The greater contribution of this form in the sample underneath the seabird colony at Dunderdalen indicated the direct fertilization effect of the seabirds, but it had no significant influence on the distribution of P among fractions. At both sites, the Pi–CaMg fraction dominated in the Pi pool, which could result from a high pH (6–7) of these soils. Such a correlation is confirmed by the findings of many studies (Ivanoff et al. Citation1998; Ann et al. Citation2000; Schlichting et al. Citation2002; Litaor et al. Citation2004).
Most of the soil phosphorus is stored in organic forms, which has been reported in numerous studies (Richardson & Marshall Citation1986; Ivanoff et al. Citation1998; Reddy et al. Citation1998; Robinson et al. Citation1998; Schlichting et al. Citation2002; Weintraub Citation2011; Wang et al. Citation2012). Such a situation occurred in the soils of the Skilvika profile. The Po–HuAc fraction played a significant role among the organic forms of phosphorus. At the Skilvika site, this form dominated in the Pt pool, which indicated the highly natural character of this site. Research indicated that the greater the contribution of the organic P fraction associated with humic acids and the smaller the contribution of the inorganic fraction associated with Fe and Al in the Pt pool, the smaller the extent of peat soil degradation (Schlichting et al. Citation2002). Very low values of the Po–HuAc form occurred near the seabird colony and increased along the Po–HuAc transect. This indicates a strong degradation of the organic soil by the seabirds. Although most of the total soil phosphorus pool was stored in organic forms, mineral phosphorus dominated in the samples near the seabird colony, reaching 92% of the Pt pool. Large quantities of phosphorus from seabird droppings were delivered there. This delivery was a mixture of inorganic P and labile organic P that was readily converted to inorganic P by soil microbes. Due to the short polar summer and consequently the short growing season, the vegetation showed a lower rate of mineral phosphorus compound intake (Dowding et al. Citation1981; Myrcha & Tatur Citation1991; Weintraub Citation2011). Accordingly, the fertilizing effect of the seabirds in the polar climate is very pronounced indeed. The other fraction of Po, Po–Res, indicated the presence of phosphorus associated with the highly durable organic plant material (Ivanoff et al. Citation1998). The similar percentage contribution of this fraction in the Pt pool at both sites under study indicates that the accumulation of this form was not linked to the impact of the seabirds.
The ratio of the Po to Pi quantities (Po/Pi) is an indicator of the natural character of peat soils. Schlichting et al. (Citation2002) concludes that the large contribution of organic P and small contribution of inorganic P indicate that peats are less degraded and less mineralized. This is also confirmed by American studies (Koch & Reddy Citation1992; Reddy et al. Citation1998). In samples near the seabird colony, the ratio was the lowest (from 0.1 to about 1.2), which resulted from the very intensive delivery of Pi into the soils by the seabirds. At the Silvika site, not subjected to the intensive impact of the seabirds, the Po/Pi ratio was considerably higher (from 1.3 to 3.9), which indicates the highly oligotrophic character of these deposits.
Conclusions
The content of Pt and its various fractions show a large diversity in the organic soils of Spitsbergen due to the fertilizing impact of seabirds. Based on the Pt content, which is a measure of the intensity of seabirds’ impact, it can be concluded that the nutrient enrichment of organic soils by birds in the polar zone is very strong. Polar organic soils not subjected to the direct fertilizing seabirds’ impact contain amounts of Pt similar to organic soils in other climate zones.
The dominant fraction in the total Pi pool in the organic soils on Spitsbergen is the phosphorus fraction associated with calcium and magnesium, regardless of the impact of the seabirds. However, its distribution depends strongly on the distance from the seabird colony. The strong impact of the breeding seabird colony in the polar climate zone modifies the distribution of phosphorus among its organic and inorganic fractions, causing the “shifting” of phosphorus from organic to inorganic fractions. The delivery of large quantities of inorganic P disrupts its natural proportions in the soil phosphorus pool (primarily a reduced proportion of the Po–HuAc fraction in the Pt pool). The considerable contribution of the labile phosphorus fraction in the organic soils results from the direct fertilizing effect of seabirds.
Acknowledgements
The authors thank the reviewers for their valuable comments and suggestions to improve the initial manuscript.
Related Research Data
References
- Anderson W.B., Polis G.A. Nutrient fluxes from water to land: seabirds affect plant nutrient status on Gulf of California islands. Oecologia. 1999; 118: 324–332.
- Ann Y., Reddy K.R., Delfino J.J. Influence of chemical amendments on phosphorus immobilization in soils from a constructed wetland. Ecological Engineering. 2000; 14: 157–169.
- Bedrock C.N., Cheshire M.V., Chudek J.A., Goodman B.A., Shand C.A. Use of 31P-NMR to study the forms of phosphorus in peat soils. Science of the Total Environment. 1994; 152: 1–8.
- Blümel W.D., Eberle J., Weber L. Verwitterung, Genese und Bodenverbreitung im Liefdefjord/Bockfjordgebiet (NW-Spitzbergen)—Untersuchungsmethoden und erste Ergebnisse. (Weathering, genesis and soil distribution in Liefdefjord/Bockfjord region [NW Spitsbergen]—research methods and first results.). Basler Beiträge zur Physiogeographie. 1993; 15: 65–70.
- Breuning-Madsen H., Ehlers C.B., Borggaard O.K. The impact of perennial cormorant colonies on soil phosphorus status. Geoderma. 2008; 148: 51–54.
- Chang S.C., Jackson M.L. Fractionation of soil phosphorus. Soil Science. 1957; 84: 133–144.
- Comerford N.B. Lynch J.P., Deikman J. Soil phosphorus bioavailability. Phosphorus in plant biology. 1998; Rockville, MD: American Society of Plant Physiologists. 136–147.
- Condron L.M., Newman S. Revisiting the fundamentals of phosphorus fractionation of sediments and soils. Journal Soils Sediments. 2011; 11: 830–840.
- Craft C.B., Richardson C.J. Peat accretion and phosphorus accumulation along a eutrophication gradient in the northern Everglades. Biogeochemistry. 1993; 22: 133–156.
- Cross A.F., Schlesinger W.H. A literature review and evaluation of the Hedley fractionation: applications to the biogeochemical cycle of soil phosphorus in natural ecosystems. Geoderma. 1995; 64: 197–214.
- Cross A.F., Schlesinger W.H. Biological and geochemical controls on phosphorus fractions in semiarid soils. Biogeochemistry. 2001; 52: 155–172.
- Dobrovolsky V.V. Geohimija počv Špicbergena. (Geochemistry of Spitsbergen soils.). Počvovedenie. 1990; 2: 5–20.
- Dowding P., Chapin F.S. III, Wielgolaski F.E., Kilfeather P. Bliss L.C. Nutrients in tundra ecosystems. Tundra ecosystems: a comparative analysis. 1981; Cambridge: Cambridge University Press. 647–683.
- Elser J.J., Bracken M.E.S., Cleland E.E., Gruner D.S., Harpole W.S., Hillebrand H., Ngai J.T., Seabloom E.W., Shurin J.B., Smith J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecology Letters. 2007; 10: 1135–1142.
- Eurola S. The middle Arctic mire vegetation in Spitsbergen. Acta Agralia Fennica. 1971; 123: 87–107.
- Forman S.L., Miller G.H. Time-dependent soil morphologies and pedogenic processes on raised beaches, Bröggerhalvöya, Spitsbergen, Svalbard Archipelago. Arctic and Alpine Research. 1984; 16: 381–394.
- Fugler S.R. Siegfried W.R. Chemical composition of guano of burrowing petrel chicks (Procellariidae) at Marion Island. Antarctic nutrient cycles and food webs. 1985; Berlin: Springer. 169–172.
- Godzik B. Heavy metals and macroelements in the tundra of southern Spitsbergen: the effect of little auk Alle alle (L.) colonies. Polar Research. 1991; 9: 121–131.
- Göttlich K., Hornburg P. Ein Zeuge wärmzeitlicher Moore im Adventdalen auf Spitzbergen (Svalbard-Archipel). (The peatbog in Adventdalen on Spitsbergen [Svalbard Archipeligo] as a witness of warm period. Telma. 1982; 12: 253–260.
- Hawke D.J., Newman J. Inventories and elemental accumulation in peat soils of forested seabird breeding islands, southern New Zealand. Australian Journal of Soil Research. 2004; 42: 45–48.
- Headley A.D. Heavy metal concentrations in peat profiles from the High Arctic. Science of the Total Environment. 1996; 177: 105–111.
- Hogg E.H., Morton J.K. The effect of nesting gulls on the vegetation and soil of islands in the Great Lakes. Canadian Journal of Botany. 1983; 61: 3240–3254.
- Ivanoff D.B., Reddy K.R., Robinson S. Chemical fractionation of organic phosphorus in selected histosols. Soil Science. 1998; 163: 36–45.
- Jóźwik Z., Magierski J. Repelewska-Pękalowa J., Pękala K. V. Trace elements in plants and soils of coastal plains of south Bellsund (western Spitsbergen). Polar Session. Arctic natural environment problems. 1992; Lublin: Maria Curie-Skłodowska University Press. 161–169.
- Klimowicz Z., Melke J., Uziak S. The influence of relief and lithology on soil formation in west Spitsbergen. Permafrost, Sixth International Conference Proceedings. Vol. 1. 1993; Beijing, China: South China University of Technology Press. 350–355.
- Klimowicz Z., Melke J., Uziak S. Peat soils in the Bellsund region, Spitsbergen. Polish Polar Research. 1997; 18: 25–39.
- Klimowicz Z., Melke J., Uziak S., Chodorowski J. Soils of north-west part of Wedel Jarlsberg Land against a background of the Spitsbergen natural environment. 2008; Lublin: Maria Curie-Skłodowska University Press.
- Klimowicz Z., Uziak S. Soil-forming processes and soil properties in Calypsostranda, Spitsbergen. Polish Polar Research. 1988; 9: 61–71.
- Koch M.S., Reddy K.R. Distribution of soil and plant nutrients along a trophic gradient in the Florida Everglades. Soil Science Society of America Journal. 1992; 56: 1492–1499.
- Kuo S. Sparks D.L. Phosphorus. Methods of soil analysis. Part 3. Chemical methods. SSSA Book Series No. 5. 1996; Madison: Soil Science Society of America, American Society of Agronomy. 869–918.
- Låg J. Special peat formation in Svalbard. Acta Agriculturae Scandinavica. 1980; 30: 205–210.
- Ligęza S., Smal H. Accumulation of nutrients in soils affected by perennial colonies of piscivorous birds with reference to biogeochemical cycles of elements. Chemosphere. 2003; 52: 595–602.
- Litaor M.I., Reichmann O., Auerswald K., Haim A., Shenker M. The geochemistry of phosphorus in peat soils of a semiarid altered wetland. Soil Science Society of America Journal. 2004; 68: 2078–2085.
- Lityński T., Jurkowska H., Gorlach E. Analiza chemiczno-rolnicza. (Agricultural-chemistry analysis.). 1976; Warszawa: PWN.
- Mann O.H., Sletten R.S., Ugolini F.C. Soil development at Kongsfjorden, Spitsbergen. Polar Research. 1986; 4: 1–16.
- Melke J. Repelewska-Pękalowa J. Preliminary studies of fissure and slope peats in the Calypsostranda and Lyellstranda Region, west Spitsbergen. Polish Polar Studies. 26th International Polar Symposium. 1999; Lublin: Maria Curie-Skłodowska University Press. 171–175.
- Melke J., Chodorowski J. Formation of Arctic soils in Chamberlindalen, Bellsund, Spitsbergen. Polish Polar Research. 2006; 27: 119–132.
- Mulder C.P.H., Jones H., Kameda K., Palmborg C., Schmidt S., Ellis J.C., Orrock J.L., Wait D.A., Wardle D.A., Yang L., Young H., Croll D.A., Vidal E. Mulder C.P.H. Impacts of seabirds on plant and soil properties. Seabird islands: ecology, invasion, and restoration. 2011; New York: Oxford University Press. 135–176.
- Mun H.T. Effects of colony nesting of Adrea cinerea and Egretta alba modesta on soil properties and herb layer composition in Pinus densiflora forest. Plant and Soil. 1997; 197: 55–59.
- Myrcha A., Tatur A. Ecological role of the current and abandoned penguin rookeries in the land environment of the maritime Antarctic. Polish Polar Research. 1991; 12: 3–24.
- Parent L.E., Khiari L. Parent L.E., Ilnicki P. Nitrogen and phosphorus balance indicators in organic soils. Organic soils and peat materials for sustainable agriculture. 2003; Boca Raton, FL: CRC Press. 105–136.
- Reddy K.R., Wang Y., DeBusk W.F., Fisher M.M., Newman S. Forms of soil phosphorus in selected hydrologic units of the Florida Everglades. Soil Science Society of America Journal. 1998; 62: 1134–1147.
- Richardson C.J., Marshall P.E. Processes controlling movement, storage, and export of phosphorus in a fen peatland. Ecological Monographs. 1986; 56: 279–302.
- Robinson J.S., Johnston T.C., Reddy K.R. Combined chemical and 31P-NMR spectroscopic analysis of phosphorus in wetland organic soils. Soil Science. 1998; 163: 705–713.
- Sapek A., Sapek B. 1997; Falenty, Poland: Institute for Land Reclamation and Grassland Farming. . Metody analizy chemicznej gleb organicznych. Materialy Instruktażowe 115. (Methods for chemical analysis of organic soils. Instructional Materials 115.).
- Schlichting A., Leinweber P., Meissner R., Altermann M. Sequentially extracted phosphorus fractions in peat-derived soils. Journal of Plant Nutrition and Soil Science. 2002; 165: 290–298.
- Smil V. Phosphorus in the environment: natural flows and human interferences. Annual Review Energy Environment. 2000; 25: 53–88.
- Speir T.W., Cowling J.C. Orthogenetic soils of the Cape Bird Adelie penguin rookeries, Antarctica. Polar Biology. 1984; 2: 207–212.
- Tatur A. Ornithogenic soils of the maritime Antarctic. Polish Polar Research. 1989; 10: 481–532.
- Tatur A. Beyer L., Bölter M. Ornithogenic ecosystems in the maritime Antarctic—formation, development and disintegration. Geoecology of terrestrial Antarctic ice-free coastal landscapes, ecological studies 154. 2002; Berlin: Springer. 161–184.
- Thomas G.W. Sparks D.L. Soil pH and Soil Acidity. Methods of soil analysis. Part 3. Chemical methods. SSSA Book Series No. 5. 1996; Madison: Soil Science Society of America, American Society of Agronomy. 869–918.
- Uziak S. Polish pedological studies on Spitsbergen. A review. Geographia Polonica. 1992; 60: 67–78.
- Wang G., Bao K., Yu X., Zhao H., Lin Q., Lu X. Forms and accumulation of soil P in a subalpine peatland of Mt. Changbai in northeast China. Catena. 2012; 92: 22–29.
- Weintraub M.N. Bünemann E.K. Biological P cycling in Arctic and alpine soils. Phosphorus in action. Biological processes in soil phosphorus cycling. 2011; Berlin: Springer. 295–316.
- Zelikson E.M. Neustadt M.I. Palinologičeskoe issledovanie golocenovogo torfjanika na Špicbergene. Palinologija golocena. (Holocene palynology. 1971; Moscow: Academy of Science of the SSSR. 199–212. (Palynological investigation of a Holocene peat deposit from Spitsbergen.).
- Zagórski P. NW part of Wedel Jarlsberg Land (Spitsbergen, Svalbard, Norway), Orthophotomap, scale 1:25000. 2005; Lublin: Department of Geomorphology, Maria Curie-Skłodowska University/Tromsø: Norwegian Polar Institute.
- Ziółek M. Phosphorus forms in organic soils with a varying degree of transformation (on the example of the Lublin Polesie region). Polish Journal of Soil Science. 2007; 40: 179–194.