
Abstract
Palsa mires are sub-Arctic peatland complexes, vulnerable ecosystems with patches of permafrost. Permafrost thawing in palsa mires occurs throughout Fennoscandia, probably due to local climatic warming. In palsa mires, permafrost thaw alters hydrological conditions, vegetation structure and microhabitat composition with unknown consequences for invertebrate fauna. This study's objectives were to examine the role of microhabitat heterogeneity and the effects of permafrost dynamics and thaw on oribatid mite communities in palsa mires. Oribatid mites were sampled in two palsa mires in Finland and Norway. Three different types of microhabitats were examined: graminoid-dominated wet sites, herb-dominated small hummocks and evergreen shrub-dominated permafrost-underlain palsa hummocks. The results indicate that permafrost dynamics are an important factor structuring oribatid mite communities in palsa mires. The community composition of oribatid mites differed remarkably among microhabitats. Six species were significantly more abundant in permafrost-underlain microhabitats in relation to non-permafrost microhabitats. None of the species identified occurred exclusively in permafrost-underlain microhabitats. Findings suggest that permafrost thaw may not have an impact on species diversity but may alter community composition of oribatid mites in palsa mire ecosystems.
Palsa mires are sub-Arctic mire complexes, found in northern parts of Fennoscandia, Siberia, Canada and Alaska. A determinative character of palsa mire is permafrost, which occurs in patches within the mire. The optimum areas for palsa mire occurrence in northern Europe are areas of low precipitation (<450 mm), where the mean annual air temperature is between −3°C and −5°C (Luoto, Fronzek et al. Citation2004). In Finnish Lapland, the southern limit of the palsa region coincides with the −1°C mean annual air temperature (Seppälä Citation2011). In Fennoscandia, palsa mires are found at the limits of the permafrost zone and are therefore very sensitive to climatic fluctuations (King & Seppälä Citation1987; Seppälä Citation1988). Palsa mires are good climate change indicators and identified as “early warning” ecosystems (Fronzek et al. Citation2009).
Throughout their distribution range in northern Finland and Scandinavia, palsa mires are degrading (Matthews et al. Citation1997; Sollid & Sørbel Citation1998; Zuidhoff & Kolstrup Citation2000; Luoto & Seppälä Citation2003; Fronzek et al. Citation2006; Åkerman & Johanssen Citation2008; Hofgaard Citation2009), probably because of regional climatic warming. Palsa mires are highly valued in terms of nature conservation (Luoto, Heikkinen et al. Citation2004; Fronzek et al. Citation2009), and listed as a priority habitat type by European Union (Anonymous Citation2007).
Permafrost occurs in palsa mires in the form of palsas, which are large peat hummocks with a permanently frozen core, formed by frost upheaval. The formation and degradation of palsas are natural cyclic processes, which maintain microhabitat diversity in palsa mires. The degradation process of permafrost causes palsa hummocks to collapse, forming remnants such as peat ridges, thermokarst pools, depressions and smaller hummocks (Seppälä Citation1986; Zuidhoff Citation2003; Zuidhoff & Kolstrup Citation2005).
Palsa hummocks create a mosaic of microhabitats within the mire because both microenvironment and vegetation on top of palsa hummocks differ from the wet surroundings. The microenvironments provide habitats for a wide range of flora and fauna and serve as important breeding areas for many bird species (Luoto, Heikkinen et al. Citation2004). Currently, in many sub-Arctic peatlands, more palsas are collapsing than are formed. This leads to changes in surface structures, hydrological conditions and vegetation. This is occurring in palsa mires in Finnish Lapland (Luoto & Seppälä Citation2003; Luoto, Heikkinen et al. Citation2004; Seppälä Citation2005), northern Sweden (Zuidhoff & Kolstrup Citation2000; Zuidhoff Citation2003) and Norway (Sollid & Sørbel Citation1998; Hofgaard Citation2003). Malmer et al. (Citation2005) used aerial images together with ground surveys of plant communities and determined that during the last three decades, the drier sites, which are dominated by ombrotrophic hummock vegetation such as mosses and evergreen dwarf shrubs, had declined, and the wet sites, dominated by tall graminoids, expanded in palsa mire ecosystems. A similar trend consisting of a decrease in hummock areas and an increase in wet area was also documented by Zuidhoff (Citation2003). Recently, Fronzek (Citation2013) evaluated that the area where climatic conditions are suitable for palsa occurrence will decrease remarkably during the 21st century.
Permafrost thaw can lead to homogenization of habitats in palsa mire ecosystems (Luoto, Heikkinen et al. Citation2004). Habitat heterogeneity is known to maintain diverse communities of invertebrates through an increase in niche differentiation and resource availability (e.g., Anderson Citation1978; Berg et al. Citation1998; Hansen Citation2000; Verberk Citation2006). Therefore, thawing of permafrost in palsa mires may not only result in changes in plant species composition and vegetation structure, but in invertebrate species composition and diversity, too. However, the effects of permafrost thaw on invertebrates inhabiting palsa mires are poorly known.
In this study, relationships between oribatid mite (Acari: Oribatida) community composition and permafrost dynamics in two palsa mire ecosystems in northern Finland and Norway were investigated. Oribatid mites are microarthropods living in various types of habitats, including in soil and dumps, the littoral, decaying wood and tree canopies (Weigmann Citation2006). Oribatid mites are one of the most diverse and numerous invertebrate groups in peatlands, where they are an important prey for higher insects (Behan-Pelletier & Bisset Citation1994; Murvanidze & Kvavadze Citation2010). Oribatid mites are significant in ecosystem functioning, due to their importance in decomposition and nutrient cycling processes (Seastedt Citation1984). In high-latitude ecosystems, the role of microarthropods is particularly important, because larger soil macro-fauna are absent or have low densities and species diversity (Swift et al. Citation1979).
Several studies show that microhabitat and resource heterogeneity are important factors structuring oribatid mite communities (Hansen & Coleman Citation1998; Migge et al. Citation1998; Hansen Citation2000; Mumladze et al. 2013) and that Oribatida have a high degree of microhabitat preferences and food-niche partitioning (Schneider et al. Citation2004; Erdmann et al. 2007; Maraun et al. Citation2011; Nielsen et al. Citation2012). However, there is also growing evidence that no clear relationships between plant and oribatid mite diversity exist (Kaneko et al. Citation2005; St. John et al. Citation2006; Osler et al. Citation2006; Erdmann et al. Citation2012). In peatlands, moisture is an important factor affecting the spatial distribution of oribatid mites with intermediate moisture conditions being the optimum for most species (Tarras-Wahlberg Citation1961; Behan-Pelletier & Bisset Citation1994).
To my knowledge, there are no published studies regarding the communities and microhabitat preferences of Oribatida in Fennoscandian palsa mires. This study's objectives were to investigate the effects of permafrost dynamics on oribatid mite communities in a microhabitat mosaic. The following questions were addressed. (1) What kind of oribatid mite communities inhabit palsa mires? (2) Are there distinct, characteristic oribatid mite communities in different microhabitats within the mire? (3) Are there species that are found only in permafrost-underlain microhabitats? (4) How are oribatid mite communities of palsa mires affected by permafrost thaw?
Materials and methods
Study areas
Data were collected from two palsa mires: Ferdesmyra mire in east Finnmark, Norway, and Vaisjeäggi palsa mire in Utsjoki, Finland. Ferdesmyra is a ca. 15 km2 peatland area 30 km west of Kirkenes. The mean annual temperature for the area is −1°C, and the mean annual rainfall is 470 mm (Norwegian Meteorological Institute; www.eklima.no). Permafrost dynamics and thaw in Ferdesmyra mire have been surveyed by the Norwegian Institute for Nature Research. A decrease in number, size and height of palsa hummocks was documented in 1970–2008 (Hofgaard Citation2009). Today, palsa hummocks are found only in two restricted areas of the mire, and all are in decay. Sampling in Ferdesmyra was conducted in the northern part of the mire (69°48′N; 27°14′E, 71 m a.s.l), where decaying palsa hummocks still exist.
The Vaisjeäggi study area was situated in a palsa mire segment—ca. 2 km in length and 1 km in width—at the south-western end of the larger Vaisjeäggi peatland area (69°44′N; 29°16′E, 282 m a.s.l). In this area, the mean annual temperature recorded at the Kevo Subarctic Research Station (12 km south-west from the mire and at 75 m a.s.l.) is −1.6°C, and the mean annual rainfall is 415 mm (Kevo weather station; www.kevo.utu.fi). In the area where study site was located, mature palsa hummocks reach heights of 1–2 m.
Sampling
Three microhabitats in both mires were sampled. Microhabitats were selected according to field observations of dominant vegetation and presence/absence of permafrost. The presence of permafrost was detected by a vegetation survey and by detection with an iron probe. Microhabitats were: (1) permafrost-underlain palsa hummocks dominated by evergreen dwarf shrubs; (2) non-permafrost hummocks dominated by herbs; (3) adjacent wet microhabitats dominated by graminoids. Because palsa hummocks in Ferdesmyra mire are degrading, whereas they are in a mature stage in Vaisjeäggi, the study areas differed in some of the microhabitat characteristics. In Ferdesmyra, sampled palsa hummocks were between 30 cm and 1.5 m and in Vaisjeäggi between 1 and 1.5 m in height. In Vaisjeäggi, the wet habitats sampled were internal meadows dominated by Carex spp. At the Ferdesmyra study site, there were no internal meadow types of microhabitat adjacent to palsa hummocks, but thermokarst pools were abundant. These pools were formed by thawing permafrost, and have been overgrown by Sphagnum spp. Eriophorum spp. was a dominant vascular plant in the pools.
At the Ferdesmyra study site, study plots were first established in all existing palsa hummocks (N=5). After that, next to each palsa hummock, one non-permafrost hummock plot and one thermokarst pool plot were chosen. One moss/soil sample of 15 cm in diameter and 5–6 cm thick, was taken in each study plot, resulting in five replicates from all three microhabitat types, 15 samples in total. Samples were taken on 8 August 2012.
In Vaisjeäggi, 15 sampling plots, representing the three microhabitats, were established along a transect set at the south-western edge of the mire. First, study plots were established on top of five separate palsa hummocks. After that, five internal meadow sites and five non-permafrost hummocks were chosen in a random manner, resulting in 15 study plots. Two 10 cm diameter and 5–6 cm thick peat samples were taken in each study plot. From the internal meadow microhabitat in Vaisjeäggi, only four samples were analysed because one sample was damaged. Samples were taken on 6 August 2011 and 9 August 2012. Samples were originally collected for two different projects, but later on data sets from the two mires were combined. For this reason, sample sizes were different between the Vaisjeäggi and Ferdesmyra study areas.
Samples were transported to Turku, Finland, where extraction in Tullgren-Berlese type funnels started within 48 h after sampling. Funnels were run for 8 days, until all samples were dry. Oribatid mites were sorted from samples under a microscope, and stored in 75% ethanol. Adult mites were identified to species or genus level using light microscopy. They were cleaned in lactic acid before identification. Identified material was stored in the Zoological Museum of the University of Turku. The nomenclature used in this paper follows Weigmann (Citation2006).
Field measurements
The percentage of vegetation coverage around the sampling plots was measured by using vegetation quadrats of 0.5 m2 in size. Moisture content was detected from a soil/peat sample taken from each study plot. Samples were dried in 35°C and moisture content was calculated as the difference in sample weight before and after drying (fresh weight minus dry weight). The sample pH was measured in the laboratory using dried and sieved soil (one sample per sampling plot) mixed with distilled water solution (100 ml water, 10 ml dried soil) using a Precisa 900 pH meter (Dietikon, Switzerland). Soil pH could not be detected for thermokarst pool samples because samples contained solely Sphagnum, and water pH measurements were not available.
Data analysis
To compare moisture content and pH between microhabitats, analyses of variance was performed. Differences in vegetation coverage, moisture and pH between microhabitats are shown in Table and 2 . Differences in abundance of species/genera between the microhabitats and study locations (Vaisjeäggi and Ferdesmyra) were compared using a generalized linear mixed model (GLMM) Proc Glimmix procedure with Tukey-Kramer post-hoc multiple comparisons for microhabitat pairs. Because the sample size was different in the two study areas, a standardized abundance data—number of individuals +0.01/100 cm2—was used in the analysis. A small number (0.01) was added to the original values to omit the zero abundance values. Because the data was not normally distributed, and did not fit Poisson or negative binomial distribution, a gamma distribution was used in the analyses. Species/genera, which occurred in six or more study plots were included in the analyses. In Vaisjeäggi, the Proc Glimmix procedure was also used to test yearly variation in species abundance. In this analysis, original values and negative binomial distribution were used.
Table 1 Microhabitat characteristics: mean moisture content per 1 g sample and pH with standard error of the mean, dominant vegetation and mean percentage of coverage of open area covered by Sphagnum or Cladina and mosses common in hummocks (Polytrichum strictum, Dicranum spp.).
Table 2 Analyses of variance test results for pH and moisture content comparisons between microhabitats.
To illustrate the differences in oribatid mite community composition between microhabitats, non-metric multidimensional scaling (NMDS) was performed. Bray-Curtis similarity index was used. To minimize the effect of different sample sizes in the two study areas, relative abundance (% of total) of each taxon was calculated for each sample and the relative abundances were used in the analyses.
NMDS analysis and microhabitats characteristics comparisons were done with PAST version 2.17 software (http://folk.uio.no/ohammer/past/). Other analyses were performed with SAS version 6.1 software.
Results
The data consisted of 8101 specimen of Oribatida, which were identified to species or genus level. Altogether, 28 species and 30 genera were identified. These belonged to 20 families. The number or genera was bigger than number of species because some of the Oribatida were identified only to genus level.
The mean overall abundance of oribatid mites was highest in palsa hummocks, second highest in non-permafrost hummocks, and lowest in wet microhabitats (). Differences in overall abundance were significant between palsa and wet microhabitats (Tukey-Kramer test p=0.003; ). There was no difference in Oribatida abundance between the years in Vaisjeäggi (GLMM; p>0.05). One species, Scheloribates initialis, was more abundant in 2011 than in 2012 (GLMM; p=0.04).
Table 3 Mean abundance of Oribatida, 1000 individuals/m2 ±SE, in three microhabitat types, the results of Tukey-Kramer test of multiple comparison of microhabitats, and a test of differences in abundances between two sampling areas.
Six species—Carabodes labyrinthicus, Tectocepheus velatus, Neoribates aurantiacus, Ceratozetes thienemanni, Eueremaeus silvestris and Camisia biurus—were more numerous in palsa microhabitats than in non-permafrost hummocks (Tukey-Kramer test; ). Conchogneta traegardhi, Carabodes subarcticus, Carabodes marginatus and Euphthiracarus spp. were significantly more abundant in non-permafrost hummocks than in palsas (Tukey-Kramer test; ).
Palsa hummock and non-permafrost hummock microhabitats were both dominated by T. velatus. Carabodes labyrinthicus was the second most abundant species in palsas (; ). In Vaisjeäggi, Melanozetes mollicomus was the most abundant species in non-permafrost hummock microhabitats (23.5%), but in Ferdesmyra, it was more numerous in palsas than in non-permafrost hummocks. Tectocepheus velatus was the most common species in the study areas. In Ferdesmyra, the relative abundance of T. velatus was 50% of all specimens identified from palsa and 24% from non-permafrost hummocks (). In Vaisjeäggi, 29.5% of individuals identified from palsa microhabitats were T. velatus and 24.4% were C. labyrinthicus. Wet microhabitats were dominated by Limnozetes ciliatus, Platynothrus peltifer and Oppiella spp. (; ).
Fig. 1 Dominance structure of Oribatida in (a) Ferdesmyra mire and (b) in the Vaisjeäggi mire.
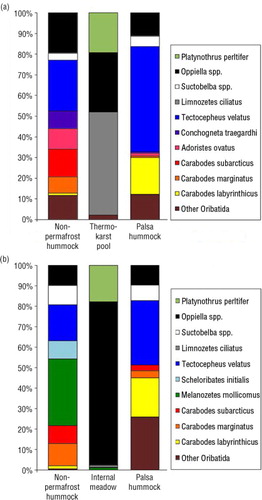
Conchogneta traegardhi, T. velatus, and C. biurus were significantly more abundant in Ferdesmyra than in Vaisjeäggi (). Melanozetes mollicomus, M. meridianus, Chamobates borealis, C. thienemanni and S. initialis were more abundant in Vaisjeäggi (). For S. initialis, the differences in abundance are simply the effect of sampling year, as it was more abundant in 2011.
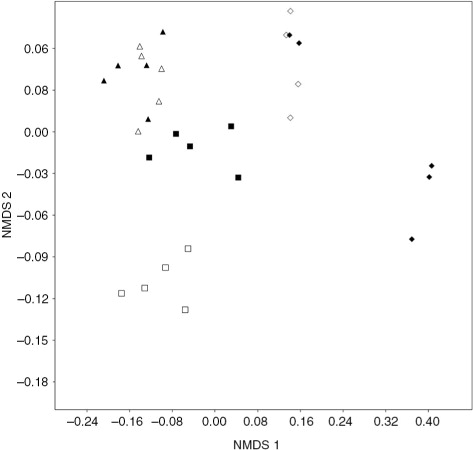
On the basis of visual interpretation of the NMDS configuration, there are distinct Oribatida communities in permafrost-underlain microhabitats in palsa mires. This is clear in Vaisjeäggi: Oribatida assemblages of palsa hummocks are located at the upper left corner of the configuration and samples taken from non-permafrost hummocks at the lower left corner. In the Ferdesmyra study area, assemblages of non-permafrost hummocks are closer to palsa hummock microhabitat assemblages than in Vaisjeäggi ().
Fig. 2 The non-metric multidimensional scaling ordination configuration based on the Oribatida data collected in two mires. Symbols represent sampling plots. Triangles represent sampling plots in palsa hummock microhabitats, squares non-permafrost hummocks and diamonds wet microhabitats. Black symbols represent Ferdesmyra and white symbols Vaisjeäggi. Stress: 0.11.
Discussion
In palsa mires, climate change is likely to change the microhabitat ratio. Karlgård (Citation2008), Bosiö et al. (Citation2012) and Fronzek (Citation2013) have predicted that the area covered by dry hummock microhabitats, such as palsas, will decrease remarkably during the coming decades in palsa mires. During the 21st century, a shift toward graminoid and carpet moss vegetation dominance is expected (Bosiö et al. Citation2012), and wet areas dominated by graminoids are predicted to increase to cover 80–87% of palsa mire areas by 2050 (Karlgård Citation2008). The area where climatic conditions are suitable for palsa occurrence will shrink remarkably (Fronzek Citation2013). If annual mean temperatures rise by 4°C in comparison to the 1961–1990 period, palsa hummocks will disappear entirely (Fronzek Citation2013).
Numerous studies address the effects of climate change in oribatid mite communities in the Arctic and sub-Arctic regions, for example, the direct effects of warming and fertilization (Coulson et al. Citation1996; Sjursen et al. Citation2005a; Hågvar & Klanderut Citation2009), the impact of altered freeze–thaw cycles (Sjursen et al. Citation2005b; Konestabo et al. Citation2008), and ice thaw in glaciers (Hågvar et al. Citation2009; see also Ingimarsdottir et al. Citation2012). Palsa mires are identified as “early warning” ecosystems vulnerable to changes in climate (Luoto, Heikkinen et al. Citation2004). Therefore the present study can add a deeper understanding of the overall effects of climate change on sub-Arctic soil faunal communities.
Even though interpretation of ordination figures is subjective (Quinn & Keough Citation2002), the NMDS analysis strengthens the idea that permafrost dynamics affect the structure and species composition of Oribatida communities (). This was particularly clear in Vaisjeäggi. Differences in community composition are also demonstrated in , which shows the dominance structure of the Oribatida communities in different microhabitats. The NMDS stress value was relatively high (0.11), but still usable (see Clarke Citation1993). It should be noted that wet microhabitats appear homogeneous because Oppiella, which were abundant in wet habitats, were identified only to genus level.
The overall abundance of Oribatida was highest in palsa microhabitats. The differences were significant only in comparison with wet microhabitats (). Therefore, a loss of palsa hummocks may only have minor impacts in overall abundance of oribatid mites in palsa mires. Mean abundances were higher than values of Oribatida abundance in Russian raised bogs (Zaitsev Citation2013), similar to abundances recorded from Caucasus lowland bogs (Murvanidze & Kvavadze Citation2010) and lower than in oligotrophic bogs in Norway (Solhøy Citation1979).
None of the species identified in this study occurred exclusively in the palsa hummock microhabitat, which suggests that a loss of permafrost would have a minor effect on oribatid mite species diversity in the studied sites. However, not all Oribatida were identified at a species level, and further conclusions should not be made. Even though most of the significant test results in habitat effects were caused by differences in species abundances between wet microhabitats and hummock habitats (palsas and non-permafrost hummocks), the results also indicate differences between permafrost-underlain and non-permafrost hummock habitats. Nine species showed microhabitat preferences between palsa and non-permafrost hummocks habitats (). Sampling location had also an effect in abundance of Limnozetes ciliatus, Nanhermannia sellnicki, and Platynothrus peltifer. This may be caused by the differences in wet microhabitats sampled, as in Ferdesmyra wet microhabitat were more pool-like than in Vaisjeäggi due to extensive permafrost thaw ().
Tectocepheus velatus and Carabodes labyrinthicus were the most numerous species in palsa hummock microhabitats (; ). Tectocepheus velatus, which was also a dominant species in non-permafrost hummocks, is a cosmopolitan species with wide habitat preferences (Weigmann Citation2006). It is a pioneer species in many habitats, such as glacier forelands and nunataks (Solhøy & Solhøy Citation2000; Hågvar et al. Citation2009; Ingimarsdottir et al. Citation2012; Flø & Hågvar Citation2013), and the most widespread species in Holarctic peatlands (Mumladze et al. Citation2013; see also Sidorchuk Citation2008). Carabodes labyrinthicus has Holarctic distribution (Karppinen & Krivolutsky Citation1982; Weigmann Citation2006) and is found in abundance in alpine heath (Solhøy & Skartveit Citation1975), under tree bark (Volker Citation1986), in tree canopies (Behan-Pelletier & Walter Citation2000), in the forest floor and in decaying wood (Siira-Pietikäinen et al. Citation2008). It is abundant in Cladina-lichen in mountain ecosystems (Materna Citation2000) and in tundra (Sidorchuk Citation2009). Other species, which were significantly more numerous in palsas than in other microhabitats, are forest dwellers (Ceratozetes thienemanni, Eueremaeus silvestris), forest-meadow dwellers (Neoribates aurantiacus) or prefer variety of habitats (Camisia biurus). Species, which were significantly more abundant in non-permafrost hummocks, are found both in forests and peatlands (Carabodes subarcticus, C. margninatus), or mainly in forests (Conchogneta traegardhi; Weigmann Citation2006; Huhta et al. Citation2012). Carabodes labyrinthicus, N. aurantiacus and E. silvestris were more numerous in palsa hummocks despite the sampling location (). In palsa mires, these species may have permafrost-indicator potential, and are also most likely to be affected by a loss of permafrost-underlain microhabitats.
The main differences in community composition between permafrost-underlain and non-permafrost hummocks were caused by a high number of C. labyrinthicus in palsas. This finding is of interest because in high-latitude ecosystems, which are relatively simple from a biodiversity perspective, species loss or gain can have strong impacts on ecosystem functions (Chapin & Körner Citation1995). Even though the presence of permafrost hummocks in palsa mires may not be an important factor maintaining oribatid mite diversity, their loss can have important, yet unknown, consequences in ecosystem functioning. Single species can have an irreplaceable role in soil systems (Laakso & Setälä Citation1999a, Citationb). Siepel & Maaskamp (Citation1994) reported that presence of C. labyrinthicus had negative effect on decomposition rates and CO2 production in microcosm experiments. This finding is linked to species traits. C. labyrinthicus is a herbofungivore, which cannot digest chitin of fungal cell-walls. Feeding solely on fungal cell contents had negative impact on fungal activity, which resulted in lower decomposition rates (Siepel & Maaskamp Citation1994). However, microcosm experiments lack multiple species interactions. Also, spatial and temporal effects, which may have a minor role in microcosm, are very important in nature (Siepel & Maaskamp Citation1994). The role of individual species in ecosystem functions is often ecosystem-specific (Bardgett & Wardle Citation2010). Whether individual oribatid mite species have effect on the ecosystem functions such as decomposition and CO2 production in palsa mires, remains to be studied.
Oribatid mites are known to show a high degree of microhabitat preferences and trophic niche partitioning (Schneider et al. Citation2004; Erdmann et al. Citation2007; Maraun et al. Citation2011; Nielsen et al. Citation2012). Schneider et al. (Citation2004) reported that species in the genus Carabodes had distinct stable isotope signals of nitrogen, indicating the species to occupy different trophic niches. The finding that morphologically similar species can have very different diets, highlights the importance of species-level identifications in Oribatida studies. Recently Mumladze et al. (Citation2013) concluded that the composition of the peat bog oribatid community at a local scale is a function of inter-specific interactions, in particular food-niche differentiation. Permafrost thaw is likely to alter these interactions because environmental changes can cause shifts in functional space occupation in a community (see Mouillot et al. Citation2013). After environmental change, species with traits that are poorly adapted to the new environment may disappear, and colonization by better-adapted species is likely to occur (Mouillot et al. Citation2013). In palsa mires, permafrost thaw and decrease of permafrost-underlain microhabitats may lead to dominance of generalist, peatland-specialist and hydrophilous oribatid mite species.
Finally, several studies have reported a high degree of unknown patterns in spatial variability of oribatid mites (Caruso et al. Citation2012; Nielsen et al. Citation2012; Franklin et al. Citation2013), and that sampling effort has remarkable effect on biodiversity investigations of Oribatida (Franklin et al. Citation2013). Therefore, a study with finer microhabitat resolution (separation among lichen, ericaceous plants, different moss types etc.) with all Oribatida individuals identified to species level would be necessary to determine the small-scale distribution patterns of oribatid mites in palsa mires, and to study the links between oribatid mite community composition and vegetation structure. A comparative study of oribatid mite communities of palsa mires and other sub-Arctic mire types, especially aapa mires, would also contribute to our understanding of the effects of permafrost thaw in oribatid mite diversity in northern peatlands. This study revealed that permafrost dynamics are an important factor structuring oribatid mite communities in sub-Arctic palsa mires.
Acknowledgements
I am grateful to Tero Klemola and Jouko Katajisto for their help with statistics. Steve Coulson, and an anonymous referee are acknowledged for their valuable comments. Thank you to Pekka Niemelä, Ritva Penttinen and Minna Turunen for commenting on the manuscript. Kevo Subarctic Research Station staff is acknowledged for providing facilities for fieldwork, and Pekka Markkula for assistance in sampling. This study has been funded by Oskar Öflund Foundation, Societas pro Flora et Fauna Fennica and Vuokon Luonnonsuojelusäätiö. Permission for sampling in Ferdesmyra was provided by the County Governor of Finnmark. Robert M. Badeau of Aura Professional English Consulting has provided the language content editing for this manuscript.
Related Research Data
References
- Åkerman H.J., Johansson M. Thawing permafrost and thicker active layers in sub-Arctic Sweden. Permafrost and Periglacial Processes. 2008; 19: 279–292.
- Anderson J.M. Inter- and intra-habitat relationships between woodland cryptostimgata species diversity and the diversity of soil and litter microhabitats. Oecologia. 1978; 32: 341–348.
- Anonymous. Interpretation manual of European Union habitats. EUR 27. 2007; Brussels: European Commission, DG Environment.
- Bardgett R.D., Wardle D.A. Aboveground–belowground linkages. Biotic interactions, ecosystem processes, and global change. 2010; Oxford: Oxford University Press.
- Behan-Pelletier V.M., Bissett B. Oribatida of Canadian peatlands. Memoirs of Entomological Society of Canada. 1994; 169: 73–88.
- Behan-Pelletier V.M., Walter D.E. Coleman D.C., Hendrix P.F. Biodiversity of oribatid mites (Acari: Oribatida) in tree canopies and litter. Invertebrates as webmasters in ecosystems. 2000; London: CABI Publishing. 187–195.
- Berg M.P., Kniese J.P., Bedaux J.M., Verhoef H.A. Dynamics and stratification of functional groups of micro- and mesoarthropods in the organic layer of a Scots pine forest. Biology and Fertility of Soils. 1998; 26: 268–284.
- Bosiö J., Johansson M., Callaghan T.V., Johanse B., Christensen T.R. Future vegetation changes in thawing subarctic mires and implications for greenhouse gas exchange—a regional assessment. Climate Change. 2012; 115: 379–398.
- Caruso T., Taormina M., Migliorini M. Relative role of deterministic and stochastic determinants of soil animal community: a spatially explicit analysis of oribatid mites. Journal of Animal Ecology. 2012; 81: 214–221.
- Chapin F.S. III, Körner C. Arctic and alpine biodiversity: patterns, causes and ecosystem consequences. 1995; Berlin: Springer.
- Clarke K.R. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology. 1993; 18: 117–143.
- Coulson S.J., Hodkinson I.D., Wooley C., Webb N.R., Block W., Worland M.R., Strathdee J.S. Effects of experimental temperature elevation on High-Arctic soil microarthropod populations. Polar Biology. 1996; 16: 147–153.
- Erdmann G., Otte V., Langel R., Scheu S., Maraun M. The trophic structure of bark-living oribatid mite communities analysed with stable isotopes (15N, 13C) indicates strong niche differentiation. Experimental and Applied Acarology. 2007; 41: 1–10.
- Erdmann G., Scheu S., Maraun M. Regional factors rather than forest type drive the community structure of soil living oribatid mites (Acari, Oribatida). Experimental and Applied Acarology. 2012; 57: 157–169.
- Flø D., Hågvar S. Aeral dispersal of mosses and invertebrates close to receding alpine glacier in southern Norway. Arctic, Antarctic and Alpine Research. 2013; 45: 481–490.
- Franklin E., de Moraes J., Landeiroc V.L., de Souzab J.L.P., Pequenod P.A.C.L., Magnussona W.E., de Moraisa J.W. Geographic position of sample grid and removal of uncommon species affect multivariate analyses of diverse assemblages: the case of oribatid mites (Acari: Oribatida). Ecological Indicators. 2013; 34: 172–180.
- Fronzek S. Climate change and the future distribution of palsa mires: ensemble modelling, probabilities and uncertainties. 2013. PhD thesis, University of Helsinki.
- Fronzek S., Johansson M., Christensen T.R., Carter T.R., Friborg T., Luoto M. Climate change impacts on sub-Arctic palsa mires and greenhouse gas feedbacks. 2009; Helsinki: Syke.
- Fronzek S., Luoto M., Carter T.R. Potential effect of climate change on the distribution of palsa mires in Subarctic Fennoscandia. Climate Research. 2006; 32: 1–12.
- Hågvar S., Klanderut K. Effects of simulated environmental change on alpine soil arthropods. Global Change Biology. 2009; 15: 2972–2980.
- Hågvar S., Solhøy T., Mong C.E. Primary succession of soil mites (Acari) in a Norwegian glacier foreland, with emphasis on Oribatid species. Arctic, Antarctic, and Alpine Research. 2009; 41: 219–227.
- Hansen R.A. Effects of litter habitat complexity and composition on a diverse litter microarthropod assemblage. Ecology. 2000; 81: 1120–1132.
- Hansen R.A., Coleman D.C. Litter complexity and composition are determinants of the diversity and species composition of oribatid mites (Acari: Oribatida) in litterbags. Applied Soil Ecology. 1998; 9: 17–23.
- Hofgaard A. Effects of climate change on the distribution and development of palsa peatlands: background and suggestions for a national monitoring project. NINA Project Report 21. 2003; Trondheim: Norwegian Institute for Nature Research.
- Hofgaard A. Overvåking av palsmyr. Forstegangsundersokelse i Ferdesmyra, Ost-Finnmark 2008. (Monitoring palsa mires. Initial survey of Ferdesbyra, east Finnmark 2008.) NINA Rapport 476. 2009; Trondheim: Norwegian Institute for Nature Research.
- Huhta V., Penttinen R., Pitkänen E. Cultural factors in the distribution of soil mites in Finland. Memoranda Society of Fauna and Flora Fennica. 2012; 88: 52–70.
- Ingimarsdottir M., Caruso T., Ripa J., Mangusdottir O.B., Migliorini M., Hedlund K. Primary assembly of soil communities: disentangling the effect of dispersal and local environment. Oecologia. 2012; 170: 745–754.
- Kaneko N., Yasunori S., Toshizumi M., Hasegawad M. Oribatid mite community structure and tree species diversity: a link?. Pedobiologia. 2005; 49: 521–528.
- Karlgård J. Degrading palsa mires in northern Europe: changing vegetation in an altering climate and its potential impact on greenhouse gases. 2008. Master's thesis, Lund University.
- Karppinen E., Krivolutsky D.A. List of oribatid mites (Acarina, Oribatei) of northern Palaearctic region. I. Europe. Acta Entomologica Fennica. 1982; 41: 1–32.
- King L., Seppälä M. Permafrost thickness and distribution in Finnish Lapland—results of geoelectrical soundings. Polarforschung. 1987; 57: 127–147.
- Konestabo H., Michelsen A., Holmstrup M. Responses of springtail and mite populations to prolonged periods of soil freeze–thaw cycles in a sub-Arctic ecosystem. Applied Soil Ecology. 2008; 36: 136–146.
- Laakso J., Setälä H. Sensitivity of primary production to changes in the architecture of belowground food webs. Oikos. 1999a; 87: 57–64.
- Laakso J., Setälä H. Population- and ecosystem-effects of predation on microbial-feeding nematodes. Oecologia. 1999b; 120: 279–286.
- Luoto M., Fronzek S., Zuidhoff F.S. Spatial modelling of palsa mires in relation to climate in northern Europe. Earth Surface Processes and Landforms. 2004; 29: 1373–1387.
- Luoto M., Heikkinen R.K., Carter T.R. Loss of palsa mires in Europe and biological consequences. Environmental Conservation. 2004; 31: 30–37.
- Luoto M., Seppälä M. Thermokarst ponds as indicators of the former distribution of palsas in Finnish Lapland. Permafrost and Periglacial Processes. 2003; 14: 19–27.
- Malmer N., Johansson T., Olsrud M., Christensen T. R. Vegetation, climatic changes and net carbon sequestration in a north-Scandinavian Subarctic mire over 30 years. Global Change Biology. 2005; 11: 1895–1909.
- Maraun M., Erdmann G., Fischer B.M., Pollierer M.M., Norton R.A., Schneider K., Scheu S. Stable isotopes revisited: their use and limits for oribatid mite trophic ecology. Soil Biology and Biochemistry. 2011; 43: 877–882.
- Materna J. Oribatid communities (Acari: Oribatida) inhabiting saxicolous mosses and lichens in the Krkonoše Mts. (Czech Republic). Pedobiologia. 2000; 44: 40–62.
- Matthews J.A., Dahl S.-O., Berrisfjord M.S., Nesje A. Cyclic development and thermokarst degradation of palsas in the mid-alpine zone at Leipullan, Dovrefjell, southern Norway. Permafrost and Periglacial Processes. 1997; 8: 107–122.
- Migge S., Maraun M., Scheu S., Schaefer M. The oribatid mite community (Acarina) of pure and mixed stands of beech (Fagus sylvatica) and spruce (Picea abies) of different age. Applied Soil Ecology. 1998; 9: 115–121.
- Mouillot D., Graham N.A.J., Villeger S., Mason N.W.H., Bellwood D.R. A functional approach reveals community responses to disturbances. Trends in Ecology and Evolution. 2013; 1621: 167–177.
- Mumladze L., Murvanidze M., Behan-Pelletier V. Compositional patterns in Holarctic peat bog inhabiting oribatid mite (Acari: Oribatida) communities. Pedobiologia. 2013; 56: 41–48.
- Murvanidze M., Kvavadze E. Sabelis M.W., Bruin J. The inventory of oribatid mites, the main decomposers in bogs of Colchic Lowland (Caucasus, Georgia). Trends in acarology. Proceedings of the 12th International Congress. 2010; Berlin: Springer. 175–178.
- Nielsen U.N., Osler G.H.R., Campbell C.D., Burslem D.F.R.P., van der Wal R. Predictors of fine-scale spatial variation in soil mite and microbe community composition differ between biotic groups and habitats. Pedobiologia. 2012; 55: 83–91.
- Osler G.H.R., Cole L., Keith A.M. Changes in oribatid mite community structure associated with the succession from heather (Calluna vulgaris) moorland to birch (Betula pubescens) woodland. Pedobiologia. 2006; 50: 323–330.
- Quinn G.P., Keough M.J. Experimental design and data analysis for biologists. 2002; Cambridge: Cambridge University Press.
- Schneider K., Migge S., Norton R.A., Scheu S., Langel R., Reineking A., Maraun M. Trophic niche differentiation in soil microarthropods (Oribatida, Acari): evidence from stable isotope ratios (15N/14N). Soil Biology and Biochemistry. 2004; 36: 1769–1774.
- Seastedt T.R. The role of microarthropods in decomposition and mineralization processes. Annual Review of Entomology. 1984; 29: 25–46.
- Seppälä M. The origin of palsas. Geografiska Annaler. 1986; 68: 141–147.
- Seppälä M. Clark M.J. Palsas and related forms. Advances in periglacial geomorphology. 1988; Chichester: John Wiley. 247–278.
- Seppälä M. Seppälä M. Periglacial environment. The physical geography of Fennoscandia. 2005; Oxford: Oxford University Press. 349–364.
- Seppälä M. Synthesis of studies of palsa formation underlining the importance of local environmental and physical characteristics. Quaternary Research. 2011; 75: 366–370.
- Sidorchuk E.A. Oribatid mites (Acari, Oribatei) of three fens in the northern part of European Russia. Entomological Review. 2008; 88: 485–490.
- Sidorchuk E.A. New data on the fauna of Oribatid mites (Acari, Oribatida) from the Polar Urals. Entomological Review. 2009; 89: 554–563.
- Siepel H., Maaskamp F. Mites of different feeding guilds affect decomposition of organic matter. Soil Biology and Biochemistry. 1994; 26: 1389–1394.
- Siira-Pietikäinen A., Penttinen R., Huhta V. Oribatid mites (Acari: Oribatida) in boreal forest floor and decaying wood. Pedobiologia. 2008; 52: 111–118.
- Sjursen H., Michelsen A., Holmstrup M. Effects of long-term soil warming and fertilisation on microarthropod abundances in three sub-Arctic ecosystems. Applied Soil Ecology. 2005a; 30: 148–161.
- Sjursen H., Michelsen A., Holmstrup M. Effects of freeze–thaw cycles on microarthropods and nutrient availability in a sub-Arctic soil. Applied Soil Ecology. 2005b; 28: 79–93.
- Solhøy T. Oribatids (Acari) from an oligotrophic bog in western Norway. Fauna Norvegica Serie B. 1979; 26: 91–94.
- Solhøy T., Skartveit A. Wielgolaski F.E. Influence of various temperature approximations on estimates of field respiration in a mountain community. Fennoscandian tundra ecosystems. 1975; Berlin: Springer. 111–116.
- Solhøy I.W., Solhøy T. The fossil oribatid mite fauna (Acari: Oribatida) in late-glacial and early-Holocene sediments in Kråkenes Lake, western Norway. Journal of Paleolimnology. 2000; 23: 35–47.
- Sollid J.L., Sørbel L. Palsa bogs as a climate indicator—examples from Dovrefjell, southern Norway. Ambio. 1998; 27: 287–291.
- St John M.G., Wall D.H., Behan-Pelletier V. Does plant species co-occurrence influence soil mite diversity?. Ecology. 2006; 87: 625–633.
- Swift M.J., Heal O.W., Anderson J.M. Decomposition in terrestrial ecosystems. 1979; Oxford: Blackwell Science.
- Tarras-Wahlberg N. Oribatei of a central Swedish bog and their environment. Oikos. 1961; 4: 166–171.
- Verberk W. Importance of landscape heterogeneity for the conservation of aquatic macroinvertebrate diversity in bog landscapes. Journal of Nature Conservation. 2006; 14: 78–90.
- Volker N. The bark of trees: thermal properties, microclimate and fauna. Oecologia. 1986; 69: 148–160.
- Weigmann G. Hornmilben (Oribatida). 2006; Keltern, Germany: Goecke and Evers.
- Zaitsev A. Oribatid mite communities (Acari: Oribatida) in different habitats of the Polistovsky Nature Reserve (Pskov Region, Russia). Estonian Journal of Ecology. 2013; 62: 276–286.
- Zuidhoff F.S. Palsa growth and decay in northern Sweden. Comprehensive Summaries of Uppsala Dissertations from the Faculty of Science and Technology 813. 2003; Uppsala: Uppsala University.
- Zuidhoff F.S., Kolstrup E. Changes in palsa distribution in relation to climate change in Laivadalen, northern Sweden, especially 1960–1997. Permafrost and Periglacial Processes. 2000; 11: 55–69.
- Zuidhoff F.S., Kolstrup E. Palsa development and associated vegetation in northern Sweden. Arctic, Antarctic and Alpine Research. 2005; 37: 49–60.