Abstract
This review gives an overview of the mathematical modelling of lactation curves in dairy cattle. Over the last ninety years, the development of this field of study has followed the main requirements of the dairy cattle industry. Non-linear parametric functions have represented the preferred tools for modelling average curves of homogeneous groups of animals, with the main aim of predicting yields for management purposes. The increased availability of records per individual lactations and the genetic evaluation based on test day records has shifted the interest of modellers towards more flexible and general linear functions, as polynomials or splines. Thus the main interest of modelling is no longer the reconstruction of the general pattern of the phenomenon but the fitting of individual deviations from an average curve. Other specific approaches based on the modelling of the correlation structure of test day records within lactation, such as mixed linear models or principal component analysis, have been used to test the statistical significance of fixed effects in dairy experiments or to create new variables expressing main lactation curve traits. The adequacy of a model is not an absolute requisite, because it has to be assessed according to the specific purpose it is used for. Occurrence of extended lactations and of new productive and functional traits to be described and the increase of records coming from automatic milking systems likely will represent some of the future challenges for the mathematical modelling of the lactation curve in dairy cattle.
Introduction
Why modelling the lactation curve?
The mathematical representation of milk production during the lactation period represents one of the most successful applications of mathematical modelling in agriculture (CitationFrance and Thornley, 1984). Tools that can mimic fundamental processes of milk production in different scenarios are of great help for physiologists, nutritionists and geneticists for studying and testing hypothesis on the behavior of the mammary gland machinery. Models able to forecast future milk yields supply useful information for management decisions dealing with time: lactation curve functions are currently implemented in dairy farm management softwares (CitationDe Vries, 2006). At cow level, lactation curve modeling is of help for monitoring individual yields for diet planning, early detection of diseases before the appearance of clinical signs and for selecting animals to be culled (CitationGipson and Grossman, 1989). Beside management, breeding is another important field of application. Lactations in progress of daughters of young bulls running progeny test can be projected up to 305-d in order to anticipate first crop evaluation (CitationSchaeffer and Jamrozik, 1996). Finally, lactation curve functions are used to fit additive genetic effects in random regression test day models (CitationSchaeffer, 2004).
Since the early work of CitationBrody et al. (1923) who proposed a decreasing exponential function to fit the declining phase of the lactation pattern in dairy cattle, studies on lactation curve modelling have experienced a rather waving pattern. Periods of great scientific contribution and discussion have been followed by relative decreases of interest. The development of the different approaches has been driven by specific needs of the dairy industry, by the scientific and technological achievements of lactation physiologists that provided the biological and theoretical foundations, and by computational sciences that supplied the mathematical tools.
Main features of the variable to be modelled
Milk production pattern along the lactation is the result of physiological processes of synthesis and captation from the blood stream by specialized epithelial cells of the mammary gland (CitationMepham, 1987). The core process of milk production is represented by the variation of mammary epithelial cell number and secretion activity (CitationCapuco et al., 2003). Cell number variation is the result of two events: cell proliferation, that starts in early pregnancy reaching a maximum immediately after parturition; cell remodelling and apoptosis that begins in early lactation and lasts till the dry-off (CitationHurley, 1989; CitationKnight and Wilde, 1993). For a healthy cow within a given milking regime, the variation of the cell secretory activity appears to be very little, although studies on this aspect of lactation are very limited (CitationKnight et al., 1998).
The phenotypic expression of these biological processes is represented by the so called standard shape of the lactation curve (). It is characterized by a first ascending phase from parturition till a maximum, the lactation peak, followed by a second declining slope that ends with the dry off of the animal. The curve depicted in represents the pattern that can be reconstructed by using the standard availability of data (usually one daily record per month, for an average of 8–10 per lactation).
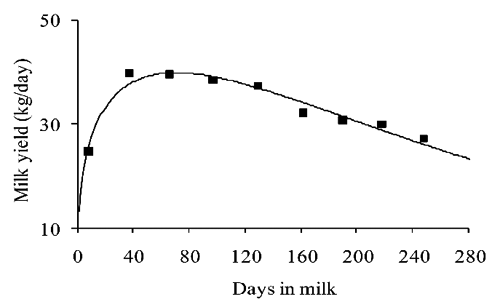
Basic traits of the curve are represented by the rate of increase of milk production in the first phase, time at peak occurrence, peak production and the rate of decline of milk yield in the second phase. The term lactation curve usually refers to the above mentioned deterministic, regular and continuous pattern depicted by the solid line in . However, a second stochastic and less predictable component, represented by individual deviations from the standard shape, exists. It can be ascribed to several factors as genetic variation between animals, feeding, farming system, environmental conditions and health status of the animal (CitationMacciotta et al., 2008a).
Lactation curve shapes different from the standard may occur, especially when individual patterns are fitted: i) a reversed shape, with an intial decreasing phase to a minimum followed by an increase, that is common for fat and protein contents; ii) a continuously increasing curve; a continuously decreasing pattern without the lactation peak i.e., the so-called atypical curve for milk yield (CitationCongleton and Everett, 1980). The most frequent (about 20–30% of cases) is the atypical shape (CitationMacciotta et al., 2005; CitationOlori et al., 1999; CitationRekik and Ben Gara, 2004). Actually, it represents mainly a computational issue due to the interaction between the mathematical structure of the model used and the combinations of test day values and distribution along the lactation trajectory (CitationCongleton and Everett, 1980; CitationMacciotta et al., 2005). In the most common dairy recording systems the milk is recorded once a month and peak yield can be easily missed. When daily data are available, as in the case of automatic milking systems, the occurrence of atypical curves is expected to be markedly smaller. However, a role of the biological differences among cows should not be excluded. In fact, an effect of calving season and parity on atypical curves occurrence has been reported (CitationRekik and Ben Gara, 2004). Another exception to the standard shape is represented by the existence of a second lactation peak in cows calving in autumn in pasture-based faming systems (CitationGarcía and Holmes, 2001). In this case, the variation in shape has to be ascribed mainly to seasonal effects, in particular to improved pasture availability in spring.
Describing and/or explaining the milk production pattern: empirical mechanistic approach. Does this differentiation make sense?
Mathematical models are a subclass of theoretical models, i.e. those aimed at representing a specific sector of reality based on its basic properties and relationships. When system features are represented by quantities, the model becomes a mathematical model. In applied biology, models are often classified into empirical or mechanistic. An empirical a model has a theory that refers just to the level of reality at which the considered phenomenon is expressed, whereas a mechanistic model is characterized by deeper theoretical assumptions. Thus, in the former the descriptive component is prevalent whereas in the latter the explicative aspects are the most important. Such a basic taxonomy is sometimes misused in agriculture, as in the specific case of lactation curve modelling. It is often assumed that empirical models are just a mere quantitative representation of the phenomenon, without any theoretical assumption, whereas all the theory is reserved to mechanistic models. Actually, some authors pointed out that a rigid distinction cannot be made (CitationNestorov et al., 1999; CitationVetharaniam et al., 2003) since each model, also the mathematically simplest and description-oriented, is based on theoretical assumptions.
A part from the theoretical foundation, that should be clearly stated, other features of a mathematical model of the lactation curve are: i) a specific mathematical structure; ii) a well defined field of application; iii) a set of potentialities but also defined borders, out of which the model can be not useful or is even misleading.
Modelling the lactation curve
Most of mathematical functions proposed to fit lactation patterns in dairy cattle are mainly aimed at describing the phenomenon. Their basic assumption is that lactation is characterized by a continuous and deterministic component with an increasing phase till a maximum followed by a decreasing slope. The mathematical tool used in this approach is represented by an analytical function of time:
where yt is the daily milk production recorded at time t. reports some of the most popular mathematical functions used to fit lactation curve data. An increase in the number of parameters, a shift from non-linear to linear equations and from models specifically conceived to fit the standard shape of the lactation curve to more general functions can be observed across years (). Such an evolution has been probably driven by several reasons, as advancements in computing or larger availability of data, although the most relevant is represented by the specific requirements of the dairy cattle industry.
Table 1 Some of the most popular empirical models used to fit the lactation curve in dairy cattle.
Early models paid more attention to the deterministic component of the lactation pattern, being essentially aimed at describing average lactation curves of homogeneous groups of animals for management purposes. An efficient model was therefore asked to disentangle the general frame of the process from environmental perturbations and to predict milk yield with good accuracy. Due to the fairly large number of records available per average curves and to the consequent regular pattern, non-linear estimation procedures could be quite easily implemented.
The Wood incomplete gamma function (CitationWood, 1967) is probably the most popular empirical model of the lactation curve. It generates the standard shape of the lactation curve as the product of a constant, a power function and an exponential decay function (). The partitioning of Wood equation into its components underlines the direct relationship of its parameters with main features of lactation curve shape (): a has a scale meaning and tends to increase with parity, ie. according to the production level of animals; b controls the rate of increase to the lactation peak; finally, c expresses the rate of decline after the peak and its absolute value tends to increase with parity. Moreover, Wood parameters can be used to estimate lactation curve traits, time at peak (tm), peak yield (ym) and lactation persistency (p):
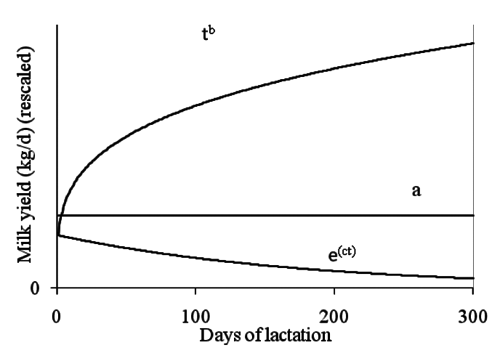
Table 2 Parameters and lactation curve traits for dairy cattle of different parities estimated with the Wood model (lactation traits have been calculated on the basis of parameter values reported in the articles).
Values for lactation curve traits reported in confirm some assessed results about relationships between lactation curve characteristics and parity number in cattle: first calving cows show a later peak occurrence, a lower peak production and a higher persistency of lactation, compared to later parities.
Limitations of the Wood model are also well known. Overestimation of daily milk yield in the first part of the curve, underestimation around and after the peak have been reported by several authors (CitationCongleton and Everett, 1980; CitationSherchand et al., 1995; CitationDematawewa et al., 2007; CitationDijkstra et al., 2010). Essentially, due to its multiplicative structure, the model is characterized by a high degree of correlation among parameters (ranges 0.70–0.90, CitationMacciotta et al., 2005) that results in a marked rigidity and in a great sensitivity to data distribution (CitationSilvestre et al., 2006). On the other hand, the model possesses some other interesting features as the ability to fit four different shapes including the atypical curves. Several modifications of the incomplete gamma function have been proposed to improve the ability of describing lactation peaks close to parturition (CitationCappio-Borlino et al., 1995) or to adequately account for seasonal variations (CitationGrossman et al., 1986). In spite of assessed limitations, the Wood model is still the most widely used function for modelling lactation curves (CitationDijkstra et al., 2010). Moreover it has been used to describe traits other than milk yield as for example milk fatty acid pattern (CitationCraninx et al., 2008), and has been used also for emerging issues, as the fitting of extended lactations (CitationDematawewa et al., 2007). Around the 1980’s, some important changes occurred in the dairy cattle industry. Individual feeding, health status monitoring, development of farm management softwares and, later on, the use of daily records instead of cumulated lactation yields for genetic evaluations, put emphasis on individual curve modelling. Mean lactation patterns of large groups of animals usually have a shape that is sufficiently regular to be fitted with a parametric function. On the contrary, large variation that occurs in individual curve shapes requires larger flexibility, easy computation, and, specifically for genetic models, an easy implementation in mixed model framework. (CitationOlori et al., 1999; CitationVerbyla et al., 1999; CitationWhite et al., 1999). Furthermore, the huge individual variation and the reduced number of records per subject (8–10 tests) resulted in a difficult use of non linear estimation procedures and thus linear or linearizable algorithms have represented an almost obliged choice, at least for routine implementations. The combined exponential and linear model of CitationWilmink (1987a) and the polynomial regression of CitationAli and Schaeffer (1987) () can be regarded as a transition between early and newer models. The Wilmink function consists of three terms that are combined additively, thus enhancing flexibility. Moreover, it can be easy linearized by setting the k parameter to a suitable fixed value (CitationBrotherstone et al., 2000) and its parameters still maintain a relationship with lactation curve shape. The Ali and Schaeffer model has a larger number of coefficients that allow for fitting a wider range of shapes although its parameters have no longer a technical meaning. Both models have been successfully used to fit individual curves (CitationMacciotta et al., 2005; CitationSilvestre et al., 2006; CitationOlori et al., 1999) and implemented in the earlier versions of random regression models (CitationDruet et al., 2003; CitationSchaeffer et al., 2000; CitationSchaeffer, 2004). These two functions have been also used for modelling traits other than milk yield as, for example, dry matter and water intake in Holsteins (CitationKramer et al., 2009) and to estimate gene effect on dairy traits (CitationStrucken et al., 2011). Although they usually outperform the Wood function, especially in different scenarios of data distribution (CitationSilvestre et al., 2006), these two models tend to yield mathematical artifacts such as negative or too high predicted values of milk yield at the beginning or at the end of lactation (CitationDruet et al., 2003; CitationMacciotta et al., 2005; CitationSilvestre et al., 2006). Above mentioned failures in describing specific stages of the lactation trajectory can be analyzed from different standpoints. First of all, it should be pointed out that less records are usually available at the edges of the lactation compared to middle stages. Under these conditions, shortcomings of extrapolating relationships between two variables beyond their known range of validity may occur when models are fitted (CitationMead et al., 2002). On the other hand, CitationGrossman and Koops (1988) stressed a further theoretical issue, common to several lactation models, represented by the consideration of the whole lactation as a single process. Starting from the assumption that lactation is the sum of different overlapping phases, these authors developed a model by adding different logistic functions (). The diphasic model, that fits two lactation phases described by two linear and two quadratic logistic functions, has been indicated as the most suitable for describing mean lactation curves for milk yield. Estimated theoretical durations of the two overlapping phases were of approximately 200 and 410 days. In general, multiphasic models are characterized by a large number of parameters (three for each phase) and therefore they require a greater number of tests to be conveniently fitted, as in case of average curves and extended lactations (CitationDematawewa et al., 2007; CitationVargas et al., 2000).
Looking for extra-flexibility: polynomials and non-parametric functions
The need for fitting individual deviations from a mean pattern led animal scientists towards more flexible mathematical tools. A common criticism to parametric models, i.e. those whose coefficients show a direct relationship with curve traits, is their high sensitivity to data distribution which may markedly affect the whole shape of the curve. As possible alternatives to parametric models, general functions not specifically conceived to fit lactation curve shape have been proposed from the early 90’. Legendre orthogonal polynomials (LP) (CitationKirkpatrick et al., 1990) or regression splines (CitationWhite et al., 1999) () are characterized by a large number of parameters, a linear additive mathematical structure and a remarkable capacity to fit a great range of shapes. Theoretically, a LP of order n is able to describe 2n different shapes: for example, a fourth-order LP has been able to recognize 18 out of the 32 theoretical shapes, represented by specific deformations of the two basic shapes, standard and atypical (CitationMacciotta et al., 2005). Legendre polynomials are currently used to fit random effects (genetic additive and permanent environment) in random regression model. In any case they still show a remarkable sensitivity to data distribution, especially at the edges of the lactation trajectory (border effects) (CitationDruet et al., 2003; CitationLópez-Romero and Carabaño, 2003). Splines are a type of segmented regression in which the curve is divided into different intervals of the dependent variable, joined at points named knots, each fitted with different polynomials (CitationGuo and White, 2005). Thus individual observations influence only a part of the function (CitationJamrozik et al., 2010). Cubic splines regressions resulted a good compromise between fitting performances, data sensitivity, smoothness and parametrisation in fitting average curves (CitationDruet et al., 2003; CitationSilvestre et al. 2006). Moreover, they are able to fit local sudden variations as in the case of individual somatic cell pattern around an event of clinical mastitis (Citationde Haas et al., 2002). A technical issue in spline fitting is represented by the optimization of the number and location of knots. Some authors recommend that knots should be as many as possible, placed at points of maximum concentration of records (CitationSilvestre et al., 2006; CitationDruet et al., 2003; CitationMisztal, 2006), even if such a criteria necessarily increases the number of function parameters. In several papers, knot number and position are fixed a priori, usually evenly spaced (CitationDruet et al., 2003; CitationHickson et al., 2006; CitationSilvestre et al., 2006). As an alternative, knot positions can be estimated by considering them as additional independent variables in a non linear estimation procedure (CitationFadel, 2004; CitationMacciotta et al., 2010). reports results of a non linear estimation of knot position for linear, quadratic and cubic splines fitted to average lactation curves of first and third calving Canadian Holsteins (CitationMacciotta et al., 2008c). Knots are located mainly in the first part of lactation, around the peak, i.e. where main curvatures occurs. Such non linear estimation, however, becomes problematic for higher order splines and large number of knots. A recent paper of CitationJamrozik et al. (2010) pointed out that the optimal knot position varies markedly depending on the effect to be modelled (genetic, permanent, environmental), the trait and the population considered. If the main aim of empirical modelling is to disentangle the deterministic component of lactation pattern, the rush for extra flexibility can have a further theoretical implication. reports the lactation curve of a third calving cow fitted with the Wood model (a), a fourth order Legendre polynomials (b) and a cubic regression spline (c) with three knots placed at 16, 69 and 252 days in milk (DIM), respectively. It can be noticed that the Wood function, although not able to fit all the local variation of test records, is able to reconstruct the shape of the curve. On the other hand, Legendre polynomials and splines are slightly efficient in fitting waves in the middle of lactation but their ability to disentangle the main traits of the phenomenon is rather questionable.
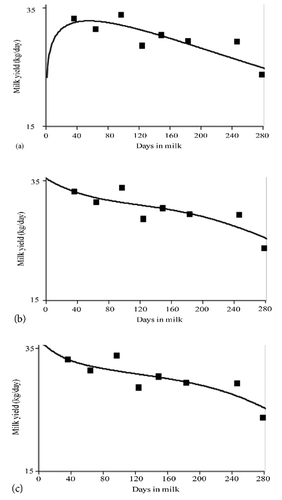
Table 3 Knot positions (days in milking) and Mean Squared Error of average lactation for regression splines of different orders fitted to average lactation curves of cows of three parities by using a non linear regression procedure.
Evaluation of factors affecting curve shape: the use of mixed linear models
Variation in lactation curve shape could be caused by several external factors such as calving season, feeding, health status, climate (CitationDijkstra et al., 2010). Under specific farming conditions, as in the case of pasture-based systems of temperate regions, seasonal and physiological effects on lactation curve shape are confounded (CitationGarcía and Holmes, 2001). In the functional approach, effects of environmental factors are evaluated either by estimating average curves of cows grouped according to sources of variation (i.e. different calving seasons or areas of production) or by comparing parameter values and (co)variances of individual curves (CitationWood, 1976; CitationShanks et al., 1981; CitationFerris et al., 1985; CitationStrucken et al., 2011). However, results of these analyses have to be carefully handled, considering the great variation that characterizes parameter values estimated on individual data (CitationOlori et al., 1999) and the different meaning of the parameters within different curve shapes (CitationMacciotta et al., 2005).
Moreover, an implicit assumption of continuous functions of time is that environmental factors average out over lactation. This is correct if effects are the same for all test day records but it is well known that some factors of variation, such as pregnancy status, weather conditions, change of management groups, are not constant throughout the lactation (CitationJamrozik and Schaeffer, 1997). Mixed linear models represent a mathematical tool more suitable for evaluating environmental effects, being able to account for factors that could affect each TD differently. The basic structure of mixed linear models used to fit daily milk yields along the lactation trajectory, usually named as test day models, is the following:
[1] where y is the daily milk yield, HTD is the interaction between the herd and the date of the test which accounts for the effects peculiar to a specific date; F represents some kind of fixed factors (i.e. calving season, region of production, parity); DIM is the fixed effect of days in milk classes, whose least squares solutions allow to generate lactation curves corrected for other effects included in the model (CitationStanton et al., 1992); L is the random effect of the individual cow and it is associated to a variance component σ2L; e is the random residual, associated at the variance component σ2L.
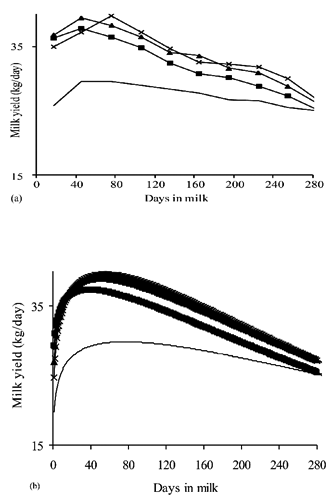
The use of levels of a fixed factor (DIM) to model lactation curve has some advantages. No previous assumptions on the shape of the curve are made and the influence of data is local because a specific TD record in a certain class does not affect the estimate for any other class (CitationDruet et al., 2003). Specific curves for different factors can be estimated by nesting the DIM class into the specific source of variation (CitationStanton et al., 1992). For example, A represents average lactation curves of dairy cattle of three different parities estimated with test day model in which the DIM factor is nested within parity. The difference with the continuous functional approach can be clearly observed in B, that reports the same curves reconstructed by fitting the Wood model to daily yields averaged by parities.
The mathematical modelling of lactation curve by mixed linear models supplied the theoretical frame for genetic evaluations based on daily yields developed during the 90’s. Structure [1] with the animal (L) effect split into genetic additive and permanent environment, represents the basic form of the test day repeatability models used for dairy cattle (CitationPtak and Schaeffer, 1993). The further inclusion of continuous functions (as the Wilmink model or LP polynomials) to fit random additive and environmental effects resulted in the development of random regression models (CitationSchaeffer, 2004). Pregnancy has a negative effect on milk production in dairy cattle. When comparing cows managed for 12 or 18 calving intervals (i.e. with pregnancies occurring at about 85 and 250 DIM, respectively), CitationBertilsson et al. (1997) found similar milk production patterns in the first 24–32 weeks. Beyond this period, the milk production of cows at the 12-month cycle tended to drop quickly, whereas that of cows at the 18-month cycle maintained a constant slope. Different lactation curve shapes have also been reported by CitationBrotherstone et al. (2004) for cows within different days open period. The lactation curve patterns for pregnant cows at different DIM and non-pregnant ones were similar until around 200 DIM. Afterwards, between 200 and 250 DIM, the decrease of production was more rapid in cows with short calving interval than in those with longer calving intervals. This confirms the reduction in milk yield and the change in milk composition that occur around month 6 of pregnancy. Indeed, it has been estimated a reduction of about 5 kg of ECM/d at day 305 of lactation for cows which had become pregnant at 85 DIM compared to those pregnant at 225 DIM (CitationAtzori et al., 2010).
A part from genetic evaluations, the use of mixed linear models to fit the lactation curve has had other applications. One is represented by the prediction of daily milk yields from a limited number of recorded TDs (CitationPool and Meuwissen, 2000; CitationVasconcelos et al., 2004). Mixed models are also used in longitudinal studies carried out on dairy animals (CitationTempelman, 2009), where they are commonly used to test research hypothesis and to evaluate effects of feeding treatments, genotypes at candidate loci, health status on milk production traits (CitationWilson et al., 2004; CitationMacciotta et al., 2008b; CitationSmith et al., 2009).
Exploring the correlation structure among test day records along the lactation
From a statistical standpoint, daily milk records taken at different time intervals from parturition are a case of repeated measures design. In particular, they can be defined as a split-plot in time design (CitationDiggle et al., 1994): the animal is the whole plot and time intervals at which milk measurements are taken represent the sub-plots. Such a data frame is characterized by a complex (co)variance structure within and between subjects, as depicted in . Milk yields close in time are more correlated than those far apart in time. Such a covariance pattern can be explained, at least in part, by common environmental factors that affect adjacent tests as weather conditions and feed (CitationAli and Schaeffer, 1987). Moreover, measures recorded on the same cow are more correlated than those recorded on different animals because they share a common contribution from the same individual.
Table 4 Sample variances (kg2), on the diagonal, and correlations, off diagonal, of test day records for milk yield along the lactation estimated with a mixed linear model without any assumption of (co)variance structure.
Variances express a rather waving behaviour, with highest values at the lactation edges. This pattern has been frequently highlighted in genetic (co)variance functions estimated with random regression models on milk yield (CitationJamrozik et al., 1997; CitationJaffrezic et al., 2000). Correlations among TD pairs at different time distance show a decreasing pattern, although values lagged the same number of observation apart show different values. Actually, heterogeneity of variance and serial correlation are common features of longitudinal data (CitationLittell et al., 1998; CitationNúñez-Antón and Zimmerman, 2000).
The complex covariance structure among test day yields must be adequately modelled because it may affect the rate of type I and II errors of tests for fixed effects (CitationLittell et al., 1998; CitationWang and Goonewardene, 2004). Variances and correlations reported in are estimated without assumptions of the underlying model. Such an unstructured (co)variance requires the estimation of t(t+1)/2 parameters, being t the number of time intervals considered. Several models have been used to fit adequately the data reducing at the same time model parametrisation. The most common structure, known as compound symmetry (CS), assumes constant variances and covariances of test day records along the lactation. Such a structure implies the estimation of 2 (co)variance parameters: σ2L (between subjects) and σ2e (within subjects). The individual lactation variance can be used for calculating of the average correlation, or repeatability, among TD records within lactation as (σ2L (σ2L + σ2e)). Repeatability values for daily milk yield in cows have been estimated to be between 0.3–0.6 (CitationStanton et al., 1992; CitationMacciotta et al., 2008b). These figures are of the same magnitude of repeatabilities estimated with BLUP models that use the pedigree relationship matrix to structure the genetic covariance between animals. Actually in this approach the σ2L component could be approximately considered as the sum of the genetic effect and the permanent environmental effect.
Some authors have proposed the use of first order autoregressive structure (AR1) to model permanent environmental variance along the lactation using test day models (Carvalheira et al., 1998; CitationWilson et al., 2008). In this structure the variance is constant whereas the correlation between two observation lagged k is ρk. CitationNúñez-Antón and Zimmerman (2000) indicated the ante-dependence structure, that fits both heterogeneous variances and covariances, as the best model for milk production data. Their results are in agreement with those obtained by CitationWang and Goonewardene (2004) on growth curves of steers. In are reported (co)variance parameters estimated on the same data set of but with the above mentioned (co)variance structures together with two popular statistics of goodness of fit, the Akaike’s Information Criterion (AIC) and the Bayesian Information Criterion (BIC).
Table 5 Comparison between different (co)variance structure used in mixed models to fit milk test day yields using Akaike’s Information Criterion (AIC) and Bayesian Information Criterion (BIC).
As far as the goodness of fit is concerned, the best value (i.e. the lowest) of the AIC is for the UN structure because, obviously, it gives the closest representation of data. The ante-dependence structure gets the best score if the BIC criterion, more penalizing for the number of parameters, is considered. The value of the autoregressive parameter reported in is of the same order of magnitude of those reported by CitationVasconcelos et al. (2004) for Portuguese Holstein cattle.
The correlation structure between TD pairs along the lactation has been largely exploited for prediction purposes. Methods currently used to predict total lactation yields for genetic evaluation purposes are based on correlation between tests in different stages of lactation (CitationNorman et al., 1999). reports correlation between actual and predicted lactation yields in first parity cattle by using different number of tests available and different approaches. Methods considered are a multiple trait approach based on Wood’s model (WMTP) (CitationSchaeffer and Jamrozik, 1996), an autoregressive moving average model (ARMA) (CitationMacciotta et al., 2002) and an autoregressive test day model (ATD) (CitationVasconcelos et al., 2004). It can be seen that all approaches are able to predict cumulated yields highly correlated with actual yields (>0.90) also when two or three tests are available at the beginning of lactation.
Table 6 Correlations among actual and predicted lactation yields in first parity cows with different approaches and different plans of missing data.
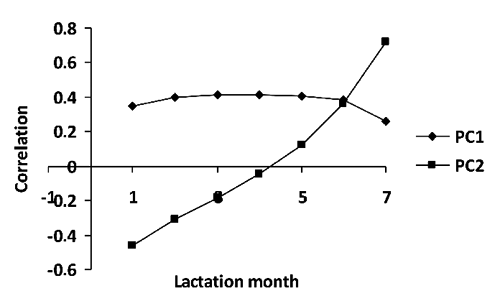
The correlation structure between milk yield test records has been investigated also using multivariate statistical techniques. The marked difference observed between values of partial and Pearson correlations can be interpreted as an evidence of an underlying latent structure of TD records (CitationMacciotta et al., 2004). The principal component analysis performed on the correlation matrix of TD records is able to extract two new variables, able to explain about 90% of the original variance, whose meaning can be inferred by structure of the correspondent eigenvectors (). The first component, explaining 75% of the original variance, is equally related to all milk tests whereas the second (about 15% of the original variance) has negative relationship with the first and positive with the second part of the lactation respectively. These new variables can be used as lactation curve traits, expressing average lactation yield and lactation curve shape, respectively (CitationMacciotta et al., 2006; CitationWilmink, 1987b) The same result has been also obtained at genetic level, by performing eigenvalue decomposition on genetic covariance matrix of TD (CitationVan der Werf et al., 1998; CitationDruet et al., 2003) or the genetic (co)variance matrix of random regression test day models (CitationJamrozik et al., 2002). A part from supplying new variables that may be of help for the genetic improvement of lactation curve traits, results of the multivariate analysis that underline the existence of a latent structure made of two latent variable seems to give an indirect confirmation of the hypothesis of CitationGrossman and Koops (1988) of a diphasic structure of the lactation pattern.
Extended lactations, a challenge for lactation curve modelling
During the last decades, an increase in the average milk yield per lactation in specialized dairy breeds has occurred due to genetic selection and improved management practices (CitationLucy, 2001; CitationSorensen et al., 2008). One side effect of the assessed strategy to increase milk yield by maximizing lactation peak and minimizing calving intervals (CitationGrossman and Koops, 2003) has been the modification of the lactation curve shape. Actually high producing breeds tend to have higher lactation peaks, a later peak occurrence and show high persistency ().
Table 7 Lactation curve traits estimated in different cattle breeds.
Production improvement, however, has resulted in an increase of fertility and health problems. The period around lactation peak is of a high risk for culling due to health problems (CitationBohmanova et al., 2009) and the negative energy balance affects reproductive efficiency (CitationFriggens et al., 2007). A straightforward consequence is the increase of average lactation length observed in many countries, with more than 50% of cows exceeding the 305 d period (CitationVargas et al., 2000; CitationGonzález-Recio et al., 2006; CitationCole et al., 2009).
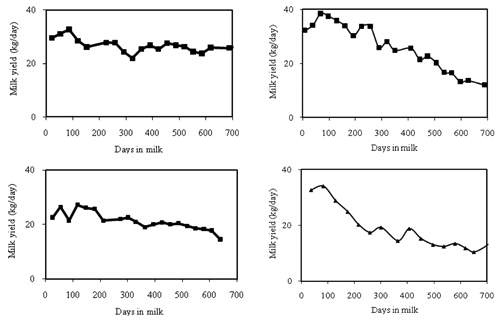
Till the 90’s, modelers paid attention almost exclusively to lactations developed within the standard length of 305 days. Actually, the extension of the lactation period does not seem to alter basic curve traits. No substantial differences in peak yields and peak occurrence were observed between extended and standard lactations (CitationVanRaden et al., 2006; CitationDematawewa et al., 2007; CitationSteri et al., 2009). Moreover, a high genetic correlation (>0.70) between milk yield in the first 5 month of lactation and production after 300 days has been reported (CitationHaile-Mariam and Goddard, 2008). Individual patterns of extended lactations usually show a relevant variability (CitationSteri et al., 2009), as reported for 305d curves. reports individual extended lactation curve patterns of Italian Holstein ( A,B) and Italian Simmental ( C,D) cows. It can be observed the occurrence of both flat ( A,C), standard ( B) and a sort of descending plus asymptotic ( D) patterns. It must be said that all these results have been obtained on extended lactations that have been caused essentially by fertility problems.
The adequacy of mathematical models to describe extended lactations has been evaluated essentially on fitting performances. CitationVargas et al. (2000) recommended the use of the diphasic function to fit extended lactations, even though they underline a relevant variability of parameter estimates. CitationGrossman and Koops (2003) developed a specific multiphasic approach, linking a first ascending phase of milk yield with a series of three descending phases of yield. A more recent comparison performed by CitationDematawewa et al. (2007) confirmed the good fitting performances of the multiphasic approach but also the computational problems related to the estimation of a high number of parameters. Authors concluded in favour of simpler models, as the Wood or the Rook () function, at least for routine use.
Even though most of extended lactations are the result of reproduction failures, a voluntary delay of insemination may increase the profitability of cows both in conventional and pasture-based farming systems (CitationRotz et al., 2005; CitationButler et al., 2010). Thus, in this specific field of application, a suitable mathematical model should be able to support decision, for example, on the economical convenience in keep milking a cow. An example is represented by the use of a modified version of the Dijkstra model implemented by CitationVanRaden et al. (2006) in the calculation of the lifetime profit for US Holstein cows:
In this formula, the β0 parameter estimates a baseline that represents a sort of threshold for milk yield profitability. Thus, the suitability of the model for extended lactations depends not only on its general goodness of fit but also on the ability of describing the asymptotic phase that often occurs approximately after 350 DIM (). Values of this parameter have been estimated in about 20 kg and 19 kg 12 for US and Italian Holsteins respectively (CitationSteri et al., 2009; CitationVanRaden et al., 2006).
Lactation curve modelling for milk composition
Lactation curves for dairy traits have received little attention compared to milk yield. However they represent an important aspect of milk production trait not only for milk production but also because milk constituents are related to the energy expenditure and has a relevant importance for dairy cattle management especially at the debut of lactation. A trait of interest is the fat:protein ratio proposed by CitationButtchereit et al. (2010) as a trait able to differentiate between cows that can or cannot adapt to the challenge of early lactation.
Also for composition traits the correlation between different stages of lactation show a progressively decreasing pattern (CitationCaccamo et al., 2008).
Conclusions
During about ninety years of documented career, the mathematical description of the lactation curve in cattle has played a fundamental role in supporting animal scientists and technicians to face several scenarios of data structure for different purposes. In general, model evolution has been characterized by a relative increase of the complexity of the mathematical structure of the algorithms used. A driven force has been probably represented by the increase of data available per individual pattern that has shifted the focus of modelling from average patterns to individual deviations. However, it is clear that a function suitable for all the fields of applications hardly exists and that model validity should be assessed on the basis of its sustainability for a particular purpose (CitationTedeschi, 2006). In any case, continuous functions with a limited number of parameters and specifically conceived for describing the standard shape of the lactation curve are still preferred in many cases.
The prediction of future challenges for lactation curve modelling is an hard task to perform. Most of literature deal with milk yield whereas other conventional dairy traits as fat, protein and somatic cell have received little attention. However there have been examples of modelling of new traits related to milk nutritional quality or health status, as fatty acid composition and water intake (CitationCraninx et al., 2008; CitationKramer et al., 2009), that are becoming of great importance for breeding and management strategies. Being difficult and expensive to be measured routinely, tools able to describe underlying patterns from few data and with a relevant predictive ability will be presumably required for their modelling.
The importance of the genetic modification of the lactation curve shape in an economically desirable direction has been pointed out by several researchers (CitationDekkers et al., 1998; CitationTogashi and Lin, 2003; CitationWeller et al., 2006). Cows with curves characterized by lower peaks and higher persistency are less exposed to health and fertility problems and can exploit efficiently cheaper feeds (CitationJakobsen et al., 2002). Several models for measuring these traits have been proposed in literature but none of them has been widely accepted (CitationMacciotta et al., 2006; CitationRekaya et al., 2001). Lactation curve traits are characterised by low to moderate values of heritability, even though a a relevant variation could be observed in estimates reported in literature. Ali and Schaeffer, for example, reported h2 values of 0.12, 0.17 and 0.23 for the three parameters of the Wood model, respectively, using a maximum likely-hood multiple trait approach. Varona et al. (1998) for the same parameters found values of 0.43, 0.17 and 0.40 using a Bayesian procedure. The issue is not only mathematical but it involves breeding strategies. The increased occurrence of extended lactations may offer a different perspective to the problem. CitationKnight (2005) demonstrated that the milk yielded in three years with two (extended) or three (conventional) lactations is the same if persistency is improved of the 1% in extended lactations. Thus models able to give an early estimate of persistency in lactation in progress may represent useful tools for both breeding and management strategies.
A huge increase of daily records per animal is likely to occur due to the diffusion of automatic milking systems in cattle farming (CitationNixon et al., 2009). The modelling of large amount of individual data will undoubtedly increase the amount of information available with positive consequences for management and breeding applications. However, modelling will be asked to address new issues: control of large variation within individuals, data correction for the great variation in milk intervals and frequencies between cows, evaluation of individual deviations from expected patterns for monitoring nutritional and health status. Some early papers used simple model as the Wilmink function (CitationNielsen et al., 2010). However other non conventional methods, like time series analysis, with their ability to disentangle deterministic and periodic components from the white noise of random variation may represent a suitable alternative for addressing these issues (CitationDeluyker et al., 1990; CitationMacciotta et al., 2002).
Acknowledgements:
the authors would like to thank Ana Helena Dias Francesconi for revising and editing the manuscript.
References
- AliT.E. SchaefferL.R. 1987 Accounting for covariances among test day milk yields in dairy cows Can. J. Anim. Sci 67 637 644
- AndersenF. OsterasØ. ReksenO. GröhnY.T. 2011 Mastitis and the shape of the lactation curve in Norwegian dairy cows J. Dairy Res 78 23 31
- AtzoriA.S. SteriR. RotondoF. DimauroC. MacciottaN.P.P. CannasA. PulinaG. 2010 Bio-economic model for lactation length simulations in dairy cows Proc. 26th World Buiatrics Congr. Santiago, Chile Available from: http://www.kenes.com/buiatrics/cd/pdf/536.pdf
- BertilssonJ. BerglundB. RatnayakeG. Svennersten-SjaunjaK. WiktorssonH. 1997 Optimising lactation cycles for the high-yielding dairy cow A European perspective. Livest. Prod. Sci 50 5 13
- BohmanovaJ. MigliorF. JamrozikJ. 2009 Use of test-day records beyond three hundred five days for estimation of three hundred five-day breeding values for production traits and somatic cell score of Canadian Holsteins J. Dairy Sci 92 5314 5325
- BrodyS. RagsdaleA.C. TurnerC.W. 1923 The rate of decline of milk secretion with the advance of the period of lactation J. Gen. Physiol 5 441 444
- BrotherstoneS. ThompsonR. WhiteI.M.S. 2004 Effects of pregnancy on daily milk yield of Holstein-Friesian dairy cattle Livest. Prod. Sci 87 265 269
- BrotherstoneS. WhiteI.M.S. MeyerK. 2000 Genetic modelling of daily milk yield using orthogonal polynomials and parametric curves Anim. Sci 70 407 415
- ButlerS.T. ShallooL. MurphyJ.J. 2010 Extended lactations in a seasonal-calving pastoral system of production to modulate the effects of reproductive failure J. Dairy Sci 93 1283 1295
- ButtchereitN. StamerE. JungeW. ThallerG. 2010 Evaluation of five lactation curve models fitter for fat-protein ratio of milk and daily energy balance J. Dairy Sci 93 1702 1712
- CaccamoM. VeerkampR.F. de JongG. PoolM.H. PetriglieriR. LicitraG. 2008 Variance components for test-day milk, fat and protein yield, and somatic cell score for analyzing management information J. Dairy Sci 91 3268 3276
- Cappio-BorlinoA. PulinaG. RossiG. 1995 A non-linear modification of Wood's equation fitted to lactation curves of Sardinian dairy ewes Small Ruminant Res 18 75 79
- CapucoA.V. EllisS.E. HaleS.A. LongE. ErdmanR.A. ZhaoX. PaapeM.J. 2003 Lactation persistency: insights from mammary cell proliferation studies J. Anim. Sci 81 (Suppl. 3) 18 31
- CarvalheiraJ.G.V. BlakeR.W. PollakE.J. QuaasR.L. Duran-CastroC.V. 1998 Application of an autoregressive process to estimate genetic parameters and breeding values for daily milk yield in a tropical herd of Lucerna cattle and in United States Holstein herds J. Dairy Sci 81 2738 2751
- CobbyJ.M. Le DuY.L.P. 1978 On fitting curves to lactation data Anim. Prod 26 127 133
- ColeJ.B. NullD.J. 2009 Genetic evaluation of lactation persistency for five breeds of dairy cattle J. Dairy Sci 92 2248 2258
- ColeJ.B. NullD.J. VanRadenP.M. 2009 Best prediction of yields for long lactations J. Dairy Sci 92 1792 1810
- CongletonW.R.Jr. EverettR.W. 1980 Error and bias in using the incomplete gamma function to describe lactation curves J. Dairy Sci 63 101 108
- CraninxM. SteenA. Van LaarH. Van NespenT. Marím-TeresoJ. De BaetsB. FievezV. 2008 Effect of lactation stage on the odd- and branched-chain milk fatty acids of dairy cattle under grazing and indoor conditions J. Dairy Sci 91 2662 2677
- CruzG.R.B. RibeiroM.N. Pimenta FilhoE.C. 2009 Estimativas de parâmetros de curva de lactação de bovinos Arch. Zootec 58 695 704
- CunhaD.N.F.V. PereiraJ.C. SilvaF.F. CamposO.F. BragaJ.L. MartuscelloJ.A. 2010 Selection of models of lactation curves to use in milk production simulation systems Rev. Bras. Zootecn 39 891 902
- de HaasY. BarkemaH.W. VeerkampR.F. 2002 The effect of pathogen-specific clinical mastitis on the lactation curve for somatic cell count J. Dairy Sci 85 1314 1323
- DekkersJ.C.M. Ten HagJ.H. WeersinkA. 1998 Economic aspects of persistency of lactation in dairy cattle Livest. Prod. Sci 53 237 252
- DeluykerH.A. ShumwayR.H. WeckerW.E. AzariA.S. WeaverL.D. 1990 Modeling daily milk yield in Holstein cows using time series analysis J. Dairy Sci 73 539 548
- DematawewaC.M.B. PearsonR.E. VanRadenP.M. 2007 Modeling extended lactations of Holsteins J. Dairy Sci 90 3924 3936
- De VriesA. 2006 Economic value of pregnancy in dairy cattle J. Dairy Sci 89 3876 3885
- DiggleP.J. LiangK.Y. ZegerS.L. 1994 Analysis of longitudinal data Clarendon Press Oxford, UK
- DijkstraJ. LopezS. BanninkA. DhanoaM.S. KebreabE. OdongoN.E. Fathi NasriM.H. BeheraU.K. Hernandez-FerrerD. FranceJ. 2010 Evaluation of a mechanistic lactation model using cow, goat and sheep data J. Agr. Sci 148 249 262
- DruetT. JaffrézikF. BoichardD. DucroqV. 2003 Modeling lactation curves and estimation of genetic parameters for first lactation test-day records of French Holstein cows J. Dairy Sci 86 2480 2490
- FadelJ.G. 2004 Technical note: estimating parameters of nonlinear segmented models J. Dairy Sci 87 169 173
- FerrisT.A. MaoI.L. AndersonC.R. 1985 Selecting for lactation curve and milk yield in dairy cattle J. Dairy Sci 68 1438 1448
- FranceJ. ThornleyJ.H.M. 1984 Mathematical models in agriculture Butterworths London, UK
- FriggensN.C. BergP. TheilgaardP. KorsgaardI.R. IngvartsenK.L. LøvendahlP. JensenJ. 2007 Breed and parity effects on energy balance profiles through lactation: evidence of genetically driven body energy change J. Dairy Sci 90 5291 5305
- GarcíaS.C. HolmesC.W. 2001 Lactation curves of autumn- and spring-calved cows in pasture-based dairy systems Livest. Prod. Sci 68 189 203
- GipsonT.A. GrossmanM. 1989 Diphasic analysis of lactation curves in dairy goats J. Dairy Sci 72 1035 1044
- González-RecioO. AlendaR. ChangY.M. WeigelK.A. GianolaD. 2006 Selection for female fertility using censored fertility traits and investigation of the relationship with milk production J. Dairy Sci 89 4438 4444
- GrossmanM. KoopsW.J. 1988 Multiphasic analysis of lactation curves in dairy cattle J. Dairy Sci 71 1598 1608
- GrossmanM. KoopsW.J. 2003 Modeling extended lactation curves of dairy cattle: a biological basis for the multiphasic approach J. Dairy Sci 86 988 998
- GrossmanM. KuckA.L. NortonH.W. 1986 Lactation curves of purebred and crossbred dairy cattle J. Dairy Sci 69 195 203
- GuoQ. WhiteR.E. 2005 Cubic spline regression for the open-circuit potential curves of a lithium-ion battery J. Electrochem. Soc 152 A343 A350
- Haile-MariamM. GoddardM.E. 2008 Genetic and phenotypic parameters of lactations longer than 305 days (extended lactations) Anima 2 325 335
- HicksonR.E. Lopez-VillalobosN. DalleyD.E. ClarkD.A. HolmesC.W. 2006 Yields and persistency of lactation in Friesian and Jersey cows milked once daily J. Dairy Sci 89 2017 2024
- HurleyW.L. 1989 Mammary gland function during involution J. Dairy Sci 72 1637 1646
- JaffrezicF. WhiteI.M.S. ThompsonR. HillW.G. 2000 A link function approach to model heterogeneity of residual variances over time in lactation curve analyses J. Dairy Sci 83 1089 1093
- JakobsenJ.H. MadsenP. JensenJ. PeersenJ. CristiensenL.G. SorensenD.A. 2002 Genetic parameters for milk production and persistency for Danish Holsteins estimated in random regression models using REML J. Dairy Sci 85 1607 1616
- JamrozikJ. BohmanovaJ. SchaefferL.R. 2010 Selection of locations of knots for linear splines in random regression test-day models J. Anim. Breed. Genet 127 87 92
- JamrozikJ. KistemakerG.J. DekkersJ.C.M. SchaefferL.R. 1997 Comparison of possible covariates for use in a random regression model for analyses of test day yields J. Dairy Sci 80 2550 2556
- JamrozikJ. SchaefferL.R. 1997 Estimates of genetic parameters for a test day model with random regressions for yield traits of first lactation Holsteins J. Dairy Sci 80 762 770
- JamrozikJ. SchaefferL.R. WeigelK.A. 2002 Estimates of genetic parameters for single and multiple-country test-day models J. Dairy Sci 85 3131 3141
- KelloggD.W. Scott UrquhartN. OrtegaA.J. 1977 Estimating Holstein lactation curves with a Gamma Curve J. Dairy Sci 60 1308 1315
- KirkpatrickM. LofsvoldD. BulmerM. 1990 Analysis of the inheritance, selection and evolution of growth trajectories Genetics 124 979 993
- KnightC.H. 2005 Extended lactation: turning theory into reality Adv. Dairy Technol 17 113 123
- KnightC.H. PeakerM. WildeC.J. 1998 Local control of mammary development and function Rev. Reprod 3 104 112
- KnightC.H. WildeC.J. 1993 Mammary cell changes during pregnancy and lactation Livest. Prod. Sci 35 3 19
- KramerE. StamerE. SpilkeJ. ThallerG. KrieterJ. 2009 Analysis of water intake and dry matter intake using different lactation curve models J. Dairy Sci 92 4072 4081
- LittellR.C. HenryP.R. AmmermanC.B. 1998 Statistical analysis of repeated measures data using SAS procedures J. Anim. Sci 76 1216 1231
- López-RomeroP. CarabañoM.J. 2003 Comparing alternative random regression models to analyse first lactation daily milk yield data in Holstein-Friesian cattle Livest. Prod. Sci 82 81 96
- LucyM.C. 2001 Reproductive loss in high-producing dairy cattle: where will it end? J. Dairy Sci 84 1277 1293
- MacciottaN.P.P. DimauroC. SteriR. Cappio-BorlinoA. 2008a Mathematical modelling of goat lactation curves CannasA. PulinaG. Dairy goats feeding and nutrition CAB International Wallingford, UK 31 46
- MacciottaN.P.P. MeleM. ConteG. SerraA. CassandroM. Dal ZottoR. Cappio-BorlinoA. PagnaccoG. SecchiariP. 2008b Association between a polymorphism at the stearoyl CoA desaturase locus and milk production traits in Italian Holsteins J. Dairy Sci 91 3184 3189
- MacciottaN.P.P. MigliorF. Cappio-BorlinoA. SchaefferL.R. 2008c Issues in modelling lactation curves with regression splines J. Dairy Sci 91 (E-Suppl.1) 544 544 abstr.
- MacciottaN.P.P. MigliorF. DimauroC. SchaefferL.R. 2010 Comparison of parametric, orthogonal, and spline functions to model individual lactation curves for milk yield in Canadian Holsteins Ital. J. Anim. Sci 9 460 464
- MacciottaN.P.P. VicarioD. Cappio-BorlinoA. 2005 Detection of different shapes of lactation curve for milk yield in dairy cattle by empirical mathematical models J. Dairy Sci 88 1178 1191
- MacciottaN.P.P. VicarioD. Cappio-BorlinoA. 2006 Use of multivariate analysis to extract latent variables related to level of production and lactation persistency in dairy cattle J. Dairy Sci 89 3188 3194
- MacciottaN.P.P. VicarioD. DimauroC. Cappio-BorlinoA. 2004 A multivariate approach to modelling shapes of individual lactation curves in cattle J. Dairy Sci 87 1092 1098
- MacciottaN.P.P. VicarioD. PulinaG. Cappio-BorlinoA. 2002 Test day and lactation yield predictions in Italian simmental cows by ARMA methods J. Dairy Sci 85 3107 3114
- MeadR. CurnowR.N. HastedA.M. 2002 Statistical methods in agriculture and experimental biology Chapman and Hall/CRC London, UK
- MephamT.B. 1987 Physiology of lactation Open University Press Philadelphia, PA, USA
- MisztalI. 2006 Properties of random regression models using linear splines J. Anim. Breed. Genet 123 74 80
- NestorovI. RowlandM. HadjitodorovS.T. PetrovI. 1999 Empirical versus mechanistic modelling: comparison of an artificial neural network to a mechanistically based model for quantitative structure pharmaco-kinetic relationships of a homologous series of barbiturates AAPS Pharmsci 1 5 13
- NielsenP.P. PetterssonG. Svennersten-SjaunjaK.M. NorellL. 2010 Technical note: variation in daily milk yield calculations for dairy cows milked in an automatic milking system J. Dairy Sci 93 1069 1073
- NixonM. BohmanovaJ. JamrozikJ. SchaefferL.R. HandK. MigliorF. 2009 Genetic parameters of milking frequency and milk production traits in Canadian Holsteins milked by an automated milking system J. Dairy Sci 92 3422 3430
- NormanD.H. VanRadenP.M. WrightJ.R. SmithL.A. 1999 Mathematical representations of correlations among yield traits J. Dairy Sci 82 2205 2211
- Núñez-AntónV. ZimmermanD.L. 2000 Modeling nonstationary longitudinal data Biometrics 56 699 705
- OloriV.E. BrotherstoneS. HillW.G. McGuirkB.J. 1999 Fit of standard models of the lactation curve to weekly records of milk production of cows in a single herd Livest. Prod. Sci 58 55 63
- PoolM.H. MeuwissenT.H.E. 2000 Reduction of the number of parameters needed for a polynomial random regression test day model Livest. Prod. Sci 64 133 145
- PtakE. SchaeffernL.R. 1993 Use of test day yields for genetic evaluation of dairy sires and cows Livest. Prod. Sci 34 23 34
- RekayaR. WeigelK.A. GianolaD. 2001 Hierarchical nonlinear model for the persistency of milk yield in the first three lactations of Holsteins Livest. Prod. Sci 68 181 187
- RekikB. Ben GaraA. 2004 Factors affecting the occurrence of atypical lactations for Holstein-Friesian cows Livest. Prod. Sci 87 245 250
- RotzC.A. ZartmanD.L. CrandallK.L. 2005 Economic and environmental feasibility of a perennial cow dairy farm J. Dairy Sci 88 3009 3019
- SchaefferL.R. 2004 Application of random regression models in animal breeding Livest. Prod. Sci 86 35 45
- SchaefferL.R. JamrozikJ. 1996 Multiple-trait prediction of lactation yields for dairy cows J. Dairy Sci 79 2044 2055
- SchaefferL.R. JamrozikJ. KistemakerG.J. Van DoormaalB.J. 2000 Experience with a test-day model J. Dairy Sci 83 1135 1144
- ShanksR.D. BergerP.J. FreemanA.E. DickinsonF.N. 1981 Genetic aspects of lactation curves J. Dairy Sci 64 1852 1860
- SherchandL. McNewR.W. KelloggD.W. JohnsonZ.B. 1995 Selection of a mathematical model to generate lactation curves using daily milk yields of Holstein cows J. Dairy Sci 78 2507 2513
- SilvestreA.M. Petim-BatistaF. ColaçoJ. 2006 The accuracy of seven mathematical functions in modeling dairy cattle lactation curves based on test-day records from varying sample schemes J. Dairy Sci 89 1813 1821
- SilvestreA.M.D. AlmeidaJ.C.M. SantosV.A.C. FontesP.J.P. AlvesV.C. 2010 Modeling lactation curves of “Barrosã” beef cattle with Wood's model Ital. J. Anim. Sci 9 243 247
- SmithR.L. GrohnY.T. PradhanA.K. WhitlockR.H. Van KesselJ.S. SmithJ.M. WolfgangD.R. SchukkenY.H. 2009 A longitudinal study on the impact of Johne's disease status on milk production in individual cows 92 J. Dairy Sci 2653 2661
- SorensenA. MuirD.D. KnightC.H. 2008 Extended lactation in dairy cows: effects of milking frequency, calving season and nutrition on lactation persistency and milk quality J. Dairy Res 75 90 97
- StantonT.L. JonesL.R. EverettR.W. KachmanS.D. 1992 Estimating milk, fat, and protein lactation curves with a test day model J. Dairy Sci 75 1691 1700
- SteriR. NicolazziE.L. GaspaG. CanavesiF. DimauroC. MacciottaN.P.P. 2009 Modeling technical parameters of individual extended lactation curves in Italian Holsteins J. Dairy Sci 92 (A-Suppl.1) 124 (abstr.)
- StruckenE.M. de KoningD.J. RahmatallaS.A. BrockmannG.A. 2011 Lactation curve models for estimating gene effects over a timeline J. Dairy Sci 49 442 449
- TedeschiL.O. 2006 Assessment of the adequacy of mathematical models Agr. Syst 89 225 247
- TempelmanR.J. 2009 Invited review: Assessing experimental designs for research conducted on commercial dairies J. Dairy Sci 92 1 15
- TogashiK. LinC.Y. 2003 Modifying the lactation curve to improve lactation milk and persistency J. Dairy Sci 86 1487 1493
- Val-ArreolaD. KebreabE. DijkstraJ. FranceJ. 2004 Study of the lactation curve in dairy cattle on farms in central Mexico J. Dairy Sci 87 3789 3799
- Van der WerfJ.H.J. GoddardM.E. MeyerK. 1998 The use of covariance functions and random regressions for genetic evaluation of milk production based on test day records J. Dairy Sci 81 3300 3308
- VanRadenP.M. DematawewaC.M.B. PearsonR.E. TookerM.E. 2006 Productive life including all lactations and longer lactations with diminishing dredits J. Dairy Sci 89 3213 3220
- VargasB. KoopsW. J. HerreroM. Van ArendonkJ.A. 2000 Modeling extended lactations of dairy cows J. Dairy Sci 83 1371 1380
- VaronaL. MorenoC. Garcia CortesL.A. AltarribaJ. 1996 Bayesian analysis of Wood's lactation curve for spanish dairy cows J. Dairy Sci 81 1469 1478
- VasconcelosJ. MartinsA. Petim-BatistaM.F. ColaçoJ. BlakeR.W. CarvalheiraJ. 2004 Prediction of daily and lactation yields of milk, fat, and protein using an autoregressive repeatability test day model J. Dairy Sci 87 2591 2598
- VerbylaA.P. CullisB.R. KenwardM.G. WelhamS.J. 1999 The analysis of designed experiments and longitudinal data by using smoothing splines Appl. Stat-J. Roy. St. C 48 269 311
- VetharaniamI. DavisS.R. UpsdellM. KolverE.S. PleasantsA.B. 2003 Modeling the effect of energy status on mammary gland growth and lactation J. Dairy Sci 86 3148 3156
- WangZ. GoonewardeneL.A. 2004 The use of MIXED models in the analysis of animal experiments with repeated measures data Can. J. Anim. Sci 84 1 11
- WellerJ.I. EzraE. LeitnerG. 2006 Genetic analysis of persistency in the Israeli Holstein population by the multitrait animal model J. Dairy Sci 89 2738 2746
- WhiteI.M.S. ThompsonR. BrotherstoneS. 1999 Genetic and environmental smoothing of lactation curves with cubic splines J. Dairy Sci 82 632 638
- WilminkJ.B.M. 1987a Adjustment of test-day milk, fat and protein yield for age, season and stage of lactation Livest. Prod. Sci 16 335 348
- WilminkJ.B.M. 1987b Comparison of different methods of predicting 305-day milk yield using means calculated from within-herd lactation curves Livest. Prod. Sci 17 1 17
- WilsonD.J. GonzálezR.N. HertlJ. SchulteH.F. BennetG.J. SchukkenY.H. GröhnY.T. 2004 Effect of clinical mastitis on the lactation curve: a mixed model estimation using daily milk weights J. Dairy Sci 87 2073 2084
- WilsonD.J. GrohnY.T. BennettG.J. GonzálezR.N. SchukkenY.H. SpatzJ. 2008 Milk production change following clinical mastitis and reproductive performance compared among J5 vaccinated and control dairy cattle J. Dairy Sci 91 3869 3879
- WoodP.D.P. 1967 Algebraic model of the lactation curve in cattle Nature 216 164 165
- WoodP.D.P. 1976 Algebraic models of the lactation curves for milk, fat and protein production, with estimates of seasonal variation Anim. Prod 22 35 40