Abstract
The effects of metabolizable energy levels were evaluated on nutrient intake and ingestive behaviour in Morada Nova lambs. Forty Morada Nova lambs were used with an initial weight of 12.2±2.05 kg. Five treatments were defined according to the metabolizable energy levels (0.96, 1.28, 1.72, 2.18 and 2.62 Mcal/kg DM). The experimental model was a randomized block design. Linear effect (P<0.0001) of ME levels was determined for dry matter intake (DMI), organic matter (OM), crude protein (CP), total carbohydrates (TC), non-fibrous carbohydrates (NFC) and total digestible nutrients (TDN) in g/day. Quadratic effect was determined for NDF (P<0.017) and carbohydrate intake (P<0.003). Linear effect was determined for digestibility of DM, OM, NDF, TC and NFC. Eating, ruminating and total chewing times, expressed in hours/day, decreased linearly (P<0.0001) with the energy levels of experimental diets. Idle time increased linearly (P<0.0001) with the increase of energy levels of the rations. Feeding and rumination efficiencies were influenced by the energy levels (P<0.0001) when expressed in g DM/h. The number of ruminal boluses, number of chews and chews per ruminal bolus were not affected by the levels of ME. However, time spent chewing per ruminal bolus was influenced (P<0.0014). Increase in ME levels of diets influences nutrient intake and feeding behaviour of Morada Nova lambs during the growing period.
Introduction
The importance of small ruminants in undeveloped countries as a source of animal protein and income is increasing. In tropical regions such as Brazil, production systems are based on pastures, which represent the lowest cost feed resource for ruminant feeding (CitationDuarte et al., 2011). Consequently, animal growth is slowed, mainly due to distribution and seasonal variation in quantity and quality of forage in the tropical environments. Therefore, the use of feedlot animals has been increased in Brazil because, among other reasons, the rate of weight gain is greater with concentrate than with roughage-based diets, which reduces feeding time and consequently reduces the cost of interest on the capital invested in animals.
However, in all environments feed characteristics influence animals’ motivation to eat, dietary choices and, ultimately, nutrient intake (CitationBaumont et al., 2000; CitationHaddad, 2005; CitationLuo et al., 2000; CitationPimentel et al., 2011). In all production systems, it is generally economically sensible to maximize the proportion of forage in the diet to minimize feeding costs. When fed indoors, animals are usually fed twice a day. Sixty to 80% of daily intake is eaten during two main meals following distributions. The main characteristics of feeding behaviour are described in terms of the satiation process and motivation to eat (CitationProvenza, 1995).
The eating behaviour of ruminants, such as feed intake, ruminating time and number of chews, varies by feed type and physical characteristics and has an important effect on the digestive physiology of the ruminant. Moreover, it is used as an indicator of the physical and chemical characteristics of roughage (CitationLee et al., 2004b; CitationLee et al., 2008). In particular, the physical shape and feeding ration of roughage affects the eating and ruminating time (CitationGill et al., 1969; CitationCastle et al., 1979) and considerably changes the buffering capacity in the rumen by affecting the secretion of saliva (CitationBartley, 1976).
The Morada Nova breed has its origins in Northeastern Brazil. But there is still little scientific information about this breed and most studies have been based on breed crossing to produce heavier animals for meat (CitationMcManus et al., 2010). Feed cost represents the largest portion of the production cost. Therefore, differences among small ruminants and digestive efficiency are very important criteria for the selection of the most appropriate animal to be kept in any particular circumstance. The objective of this work was to study the differences in nutrient intake and feeding behaviour of Morada Nova lambs fed with different energy levels.
Materials and methods
Experiment site
The experiment was carried out at the Animal Laboratory of the Department of Animal Science of the Universidade Federal do Ceará (UFC) in Fortaleza-CE, Brazil. Humane animal care and handling procedures were followed according to the University’s animal care committee.
Animals, housing and feeding
Forty Morada Nova non-castrated male lambs, with average initial body weight (BW) of 12.20±2.05 kg at approximately 60 days old were used. First, animals were identified, wormed and placed in individual stalls with feeding troughs for food and water. The lambs were allocated randomly to five treatments that consisted of increasing levels of metabolizable energy (0.96; 1.28; 1.72; 2.18 and 2.62 Mcal/kg DM) obtained from different roughage:concentrate ratios (95:5, 80:20, 60:40, 40:60 and 20:80). Lambs were subjected to an adaption period of 20 days and after that were kept in confinement until they reached an average 25 kg of body weight, after which the group was slaughtered. On this occasion, a random 2 animals of the control group (0.96 Mcal/kg DM) were also slaughtered. This procedure was carried out for each group until all the animals had been slaughtered. Average days in confinement to obtain the predetermined weight were 98, 109, 146 and 174 days at levels of 2.62, 2.18, 1.72 and 1.28 Mcal/kg DM, respectively. The experimental model was a randomized block design with eight replications. The experimental diets were formulated according to the National Research Council (2007) and were offered as a total mixed ration. Animals had free access to water throughout the trial. Diets were formulated to be isonitrogenous with 16% CP (DM basis) to a 200 g daily gain, except for the ratio 95:5, formulated to meet the requirements for maintenance with 9% CP. Diets were composed of Tifton 85 hay as roughage and concentrates based on corn grain, soybean meal, urea, sodium chloride, calcium carbonate, dicalcium phosphate and mineral premix ( and ).
Table 1 Chemical composition of the ingredients in g/kg DM (unless otherwise stated).
Table 2 Composition of the experimental diets.
Particle size of the experimental feed
Particle sizes of the experimental feed were measured using a Penn State particle separator (PSPS), according to the method used by CitationKononoff and Heinrichs (2003). Samples of the experimental diets (approximately 100 g) were taken to determine particle sizes through manual agitation in the PSPS, which consisted of three sieves (1.18, 8, and 19 mm) that could separate feed into four different types depending on the particle size. This allowed the estimation of physically effective neutral detergent fibre (peNDF) according to CitationMertens (1997), whereby the concentration of NDF (% DM) of feed is multiplied by the percentage of particles retained in sieves above 1.18 mm. The value of the effectiveness of feed is equal to the product of this operation, according to the formula:
Intake and chemical analysis
Animals were fed individually ad libitum twice a day (at 08h00 and 16h00). Feed refusals were measured before the morning feeding and the amount of feed offered was adjusted to allow a 10% refusal. Daily dry matter intake (DMI) was determined by the difference between the weight of the diet offered and the orts. Every day, before the feed supply, diet orts of each animal were removed and weighed, and data were recorded in spreadsheets for daily control. Samples of feeds and orts were weighed, packed and frozen each day for subsequent chemical analysis.
Forage, concentrate and orts composited samples were dried at 55°C for 72 h in a forced air oven, then ground in a knife mill with a 1 mm screen (Wiley mill, Arthur H. Thomas, Philadelphia, PA, USA). The samples were analyzed for dry matter contents (DM) (CitationAOAC 1990; method no. 930.15), ash (CitationAOAC 1990; method no. 924.05), crude protein (CP) (CitationAOAC 1990; method no. 984.13), ether extract (EE) (CitationAOAC 1990; method no. 920.39) and acid detergent fibre (ADF) (CitationVan Soest et al., 1991). To analyze the neutral detergent fibre (NDF), the samples were treated with thermo-stable alpha amylase without the use of sodium sulfite, corrected for residual ash (CitationMertens, 2002) and for residual nitrogenous compounds (CitationLicitra et al., 1996).
The total carbohydrate content (TC) was calculated using the expression:
according to CitationSniffen et al. (1992). The non-fibrous carbohydrates (NFC) were calculated from the equation adapted from CitationWeiss (1999), where:
Because of the presence of urea in concentrate constitution, the NFC was calculated from the adapted equation by CitationHall (2000), where:
Digestibility trials were conducted to determine metabolizable energy (ME) of the diets. Indigestible acid detergent fibre (iADF) was used as a marker to estimate faecal DM excretion as described by CitationCasali et al. (2009). Feces were collected every 15 days for three consecutive days: at 08h00 on the first day, at 12h00 on the second day and at 16h00 on the third day. Samples of feces, feeds (Tifton 85 hay and concentrate) and orts from the digestibility trial were dried at 55°C, ground to pass through a 1 mm screen and then proportionally sub-sampled to a compound sample to be analyzed as reported previously. TDN was calculated according to CitationWeiss (1999):
where CPd, NFDd, NDFapd and EEd correspond to digestible crude protein, digestible non-fibrous carbohydrates, digestible neutral detergent fibre corrected for ash and protein and digestible ether extract, respectively. The dietary DE was estimated as 4.409 Mcal/kg of TDN, and DE was converted to ME using an
efficiency of 82% to convert DE to ME (NRC, 2000).
Feeding behaviour
To measure the ingestive behaviour variables, animals were submitted to visual observation in the 95th and 96th days of the experiment. First day observations were registered at 5-min intervals for 24 h to determine the time spent eating, ruminating and idle, according to the methodology proposed by CitationJohnson and Combs (1991). The shed was kept under artificial lighting at night throughout the experimental period.
On the second day, animals were observed during 3 periods of 2 h (08h00 to 10h00, 14h00 to 16h00, and 18h00 to 20h00). These data were used to estimate the number of chews per ruminal bolus and time spent chewing per ruminal bolus, using a chronometer. The variables of ingestive behaviour were obtained by the equations:
where:
FE (g DM/h, g NDF/h) is the feeding efficiency; DMI (g DM/day) is the dry matter intake; ET (h/day) is the eating time; RE (g DM/h, g NDF/h) is the ruminating efficiency;
RT (h/day) is the ruminating time; TCT (h/day) is the total chewing time; Nrb (no./day) is the number of ruminal boluses;
TCrb (sec/bolus) is the time spent chewing ruminal bolus;
NCrb (no./bolus) is the number of chews per ruminal bolus; and NCd (no./day) is the number of chews per day (CitationPolli et al., 1996).
Statistical analysis
The experimental design was a randomized block (initial body weight), with five treatments, according to the mathematical model:
where:
Yij = value observed in the plot that received the treatment i in the block j;
µ = general average of the population;
αi = effect of treatment i = 1, 2, 3, 4, 5;
βi = effect of the block j = 1, 2, 3, 4, 5, 6, 7, 8;
eij = random error.
The statistical analyses were performed using PROC GLM of the SAS version 9.0 (CitationSAS, 2003). An orthogonal partition of the sum of the square of treatments into linear, quadratic and cubic degree effects was obtained by analysis of variance. Regression equation was adjusted when 0.05 significance was observed, using PROC REG SAS (9.0).
Results and discussion
Nutrient intake
A linear effect (P<0.0001) was observed for DM, OM, CP, TC, NFC and TDN intakes (g/day) with increased diet ME levels. Increased ME levels promoted a quadratic effect for NDF (P<0.0170) and FC (P<0.0030) intakes (g/day), with critical points (maximum response) at ME levels of 1.56 and 1.44 Mcal/kg DM, respectively. A similar pattern was also observed for FC (P<0.0001) intake, when expressed in g/kgBW0.75 with critical points (maximum response) at ME levels of 1.79 Mcal/kg DM ().
Table 3 Intake of diet constituents by sheep fed with different levels of metabolizable energy.
CitationMedeiros et al. (2007) reported linear increase in intake with values of 925, 964, 1.003 and 1.124 g/day for sheep fed diets containing 20, 40, 60 and 80% concentrate, values similar to those reported by CitationMahgoub et al. (2000), when three energy levels (2.4, 2.5 and 2.7 Mcal/kg DM) were assessed in Omani growing lambs’ diets. NDF intake, when expressed in g/day, showed quadratic behaviour with the ME increase in the diets, indicating the existence of an inflection point or transition between the physical and biological control. According to CitationMertens (1987), when the energy density of the diet is high (low NDF), in relation to requirements of the animal, intake may be limited by energy demand, not rumen fill. For diets of low energy density (high content of NDF), the intake can be limited by filling the rumen reticulum.
The increase in particle size in the experimental diets with lower ME levels may also have influenced the filling effect and, consequently, the reduction of DM intake. Similarly, CitationKato et al. (1989) and CitationJeon et al. (2001) observed that the small particle size of the feed induced an increased dry matter intake.
Low-quality tropical forages are deficient not only in nutrients for animal performance, but also in substrates for microbial metabolism, mainly nitrogenous compounds (CitationDetmann et al., 2009). Thus, inclusion of protein and/or energy sources in diets would be beneficial to the rumen environment and would increase the microbial growth on the fibrous carbohydrates (CitationCosta et al., 2008). Increase in intake could be a reflection of increases in the digestibility of the fibrous compounds (CitationLazzarini et al., 2009) which exert a high rumen fill effect. Increases in the voluntary low-quality forage intake as a result of protein sources is frequently associated with higher forage passage and digestion rates, which accelerate the removal of the indigestible fibre compounds from the rumen, resulting in a higher rumen turnover (CitationPaulino et al., 2008). Particle size of diets may also influence intake. According to CitationPoppi et al. (1985), the particle size of 1.18 mm is that which determines whether a particle has a rapid (<1.18 mm) or slow (> 1.18 mm) passage by the rumen, this value being for both sheep and cattle. Thus, the larger particle size in diets with high levels of ME () allowed rumen to escape slowly and a higher filling effect.
Table 4 Particle size distribution in percentage of feeds and diets.
Thus, if daily intake is the primary goal, theories related to the regulation of long-term consumption are the most suitable as a starting point for derivation of the equations. Therefore, the optimum content of NDF in the diet should not be fixed but should vary depending on the net energy requirement of the animal. The diet (0.96 Mcal/kg DM) containing high NDF content (722.5 g/kg DM) promoted low total DMI due to restrictions caused by the filling of the animals’ rumen-reticulum. On the other hand, the diet containing a high level of energy (2.62 Mcal/kg DM) and low fibre (267.4 g/kg DM) also resulted in lower total DM intake, indicating that animal energy requirements have been reached at lower levels of consumption. Satiety as the physiological factor limiting intake may be related to diets with high caloric density (high amount of concentrate) and, in this case, animal requirements control intake.
Ingestive behaviour
Eating time and ruminating time (h/day) decreased linearly (P<0.0001) with the energy levels of the experimental diets (). A linear effect was observed for idle animals (P<0.0001). These results corroborate those obtained by CitationDulphy et al. (1980) who reported a decrease in time spent in the physiological process of rumination with increasing inclusion of dietary concentrate. CitationFrança et al. (2009) and CitationMendes et al. (2010) registered a reduction in time spent on rumination activity in Morada Nova sheep fed with different ME levels (2.2, 2.8 and 3.4 Mcal/kg DM).
Table 5 Ingestive behaviour of Morada Nova lambs fed rations with different metabolizable energy levels.
A linear effect was registered for rumination and feeding efficiency (g DM/h) at the levels of ME (P<0.0001). When expressed in g NDF/h, feeding efficiency had a significant effect (P<0.0016), meanwhile rumination efficiency had no significant effect for the diet ME levels (). CitationKim et al. (1994) reported that the shorter the length of the feed, the higher the feeding efficiency. This was also observed by CitationJeon et al. (1997) who reported that feeding efficiency was higher with roughage of a smaller particle size.
The total chewing time (min/day) was influenced by the ME level of experimental diets, with a decreasing linear effect (P<0.0001). According to CitationDulphy et al. (1980), when the cell wall constituents of the diet decreases, increasing the starch content, the total chewing time decreases, a fact detected by the linear decrease observed in the data obtained in this study (). CitationTurino (2003) observed that diets with higher NDF and physically effective neutral detergent fibre (peNDF) promote an increase in chewing activity in sheep.
The number of ruminal boluses, number of chews per day and chews per ruminal bolus were not affected by ME. However, time spent chewing by ruminal bolus was influenced (P<0.0014) by ME in the diets (). CitationLee et al. (2004a) and CitationJeon et al. (1997) reported that the shorter the cutting length (particle size) of the roughage, the lower the number of boluses.
Table 6 Ingestive behaviour of Morada Nova lambs fed rations with different metabolizable energy levels.
The values of peNDF increased with increasing fibre in the diets (), which may be indicative of longer rumination and chewing times since the concept of peNDF is related to the physical characteristics of fibre to stimulate chewing activity, because of a high correlation between time spent chewing and fibre effectiveness (CitationMertens, 1997). CitationBeauchemin and Yang (2005) reported that by decreasing the peNDF content of diets, the number of chews per day during intake was linearly reduced and tended to reduce the number of chews during rumination. Consequently, total chewing activity was reduced. The authors reported that eating time (min/day) and rumination time (min/day) were linearly affected by diet peNDF, as well as the total chewing time (min/day).
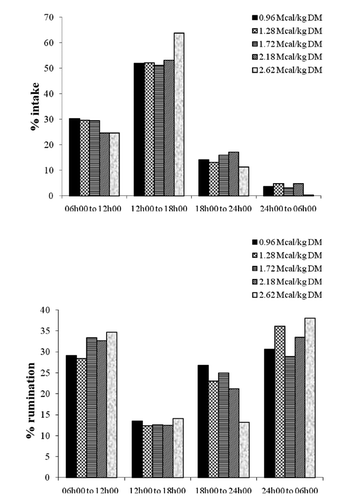
shows the distribution of consumption and ruminating activities in four periods of the day: (1) 06h00 to 12h00; (2) 12h00 to 18h00; (3) 18h00 to 24h00; and (4) 24h00 to 06h00. The sum of periods 1 and 2 corresponded to the longer time spent on consumption (82.21% of total consumption), so the consumption was concentrated during the day. These observations were also registered by CitationPereira et al. (2009), CitationDado and Allen (1995), and CitationMacedo et al. (2007), who obtained an average of 57% of time spent feeding adding the periods 1 and 2. CitationDado and Allen (1995) reported that DMI is increased after feeding, when feed is still fresh. According to CitationForbes (1995), as ruminants are diurnal animals, their feeding activity is more frequent during the day than at night; however, this can vary. Rumination occurred mainly at night, usually the time when air temperature is milder. Rumination prevailed between periods 3 and 4 (). The daily pattern of rumination activity was high after 10 h of daily feed supply (period 3) and remained active during the subsequent 6 h (period 4). CitationPolli et al. (1996) reported that the distribution of rumination activity is influenced by diet, since rumination takes place soon after the feeding periods when the animal is quiet. CitationMacedo et al. (2007) observed a percentage of 54.54% for the time spent in rumination.
Conclusions
The increase in ME levels in diets influences nutrient intake and feeding behaviour of Morada Nova lambs during the growing period.
Acknowledgments:
the authors wish to thank the Conselho Nacional de Pesquisa e Desenvolvimento Científico e Tecnológico (CNPq) and Banco Nordeste do Brasil for financial support.
References
- AOAC 1990 Official Methods of Analysis 15th ed Association of Official Analytical Chemists Washington, DC, USA
- BartleyE.E. 1976 Bovine saliva: production and function of buffers in ruminant physiology and metabolism Church and Dwight Co., Inc. New York, NY, USA
- BaumontR. PracheS. MeuretM. Morhand-FehrP. 2000 How forage characteristics influence behaviour and intake in small ruminants: a review Livest. Prod. Sci 61 15 28
- BeaucheminK.A. YangW.Z. 2005 Effects of physically effective fiber on intake, chewing activity, and ruminal acidosis for dairy cows fed diets based on corn silage J. Dairy Sci 88 2117 2129
- CasaliA.O. DetmannE. Valadares FilhoS.C. PereiraJ.C. CunhaM. DetmannK.S.C. PaulinoM.F. 2009 Estimação de teores de componentes fibrosos em alimentos para ruminantes em sacos de diferentes tecidos Rev. Bras. Zootecn 38 130 138
- CastleM.E. RetterW.C. WatsonJ.N. 1979 Silage and milk production: comparisons between three silages of different chop lengths Grass Forage Sci 34 293 301
- CostaV.A.C. DetmannE. Valadares FilhoS.C. PaulinoM.F. HenriquesL.T. MantovaniH.C. 2008 Degradação in vitro da fibra em detergente neutro de forragem tropical de baixa qualidade em função de suplementação com proteína e/ou carboidratos Rev. Bras. Zootecn 37 494 503
- DadoR.G. AllenM.S. 1995 Intake limitation, feeding behaviour and rumen function of cows challenger with rumen filt from dietary fiber or inert bulk J. Anim. Sci 78 118 133
- DetmannE. PaulinoM.F. MantovaniH.C. Valadares FilhoS.C. SampaioC. B. SouzaM.A. LazzariniI. DetmannK.S.C. 2009 Parameterization of ruminal fiber degradation in lowquality tropical forage using Michaelis-Menten kinetics Livest. Sci 126 136 146
- DuarteM.S. PaulinoP.V.R. Valadares FilhoS.C. PaulinoM.F. DetmannE. ZervoudakisJ.T. MonneratJ.P.I.S. VianaG.S. SilvaL.H.P. SerãoN.V.L. 2011 Performance and meat quality traits of beef heifers fed with two levels of concentrate and ruminally undegradable protein Trop. Anim. Health Prod 43 877 886
- DulphyJ.P. RemondB. TheriezM. 1980 Ingestive behaviour and related activities in ruminants RuckebushY. ThivendP. Digestive physiology and metabolism in ruminants MTP Press Lancaster, UK 103 122
- ForbesJ.M. 1995 Voluntary food intake and diet selection in farm animals CAB Int. Publ. Wallingford, UK
- FrançaS.R.L. Gonzaga NetoS. Pimenta FilhoE.C. MedeirosA.N. TorreãoJ.N.C. MarizT.M.A. CostaR. G. 2009 Comportamento ingestivo de ovelhas Morada Nova no terço final de gestação com níveis de energia metabolizável na dieta Rev. Bras. Saúde Prod. An 10 73 84
- GillS.S. ConradH.R. HibbsJ.W. 1969 Relative rate of in vitro cellulose disappearance as a possible estimator of digestible dry matter intake J. Dairy Sci 52 1687 1690
- HaddadS.G. 2005 Effect of dietary forage:concentrate ratio on growth performance and carcass characteristics of growing Baladi kids Small Ruminant Res 57 43 49
- HallM.B. 2000 Calculation of non-structural carbohydrate content of feeds that contain non-protein nitrogen. Bulletin No. 339 University of Florida Publ. Gainesville, FL, USA
- JeonB.T. MoonS.H. KwonY.J. KwakW.S. 2001 Effect of supplementary level of fermented broiler litter on the dry matter intake, digestibility and nitrogen balance in female spotted deer (Cervus Nippon) Korean J. Anim. Sci 43 730 731
- JeonB.T. ParkI.H. LeeS.M. KimK.H. KimJ.S. SonJ.C. 1997 The effect of different fiber sources on chewing behavior of Korean native cattle Korean J. Anim. Sci 39 385 388
- JohnsonT.R. CombsD.K. 1991 Effects of prepartum diet, inert rumen bulk, and dietary polythylene glicol on dry matter intake of lactating dairy cows J. Dairy Sci 74 933 944
- KatoK. KajimaY. OdashimaM. LeeL.S. NamK.T. ChigaH. ShojiY. OthaM. SasakiY. 1989 Feed passage and digestibility in Japanese deer and sheep. Research report of Kawatabi Experimental Station 5 59 62
- KimC.M. LeeB.S. ChungT.Y. 1994 Influence of cutting length of ammoniated barley straw on the eating and ruminating behavior of Korean native cattle Korean J. Anim. Sci 36 487 493
- KononoffP.J. HeinrichsA.J. 2003 The effect of reducing alfafa haylage particle size on cows in early lactation J. Dairy Sci 86 1445 1457
- LazzariniI. DetmannE. SampaioC.B. PaulinoM.F. Valadares FilhoS.C. SouzaM.A. OliveiraF.A. 2009 Intake and digestibility in cattle fed low-quality tropical forage and supplemented with nitrogenous compounds Rev. Bras. Zootecn 38 2021 2030
- LeeS.M. HwangJ.H. YoonY.B. KwakW.S. KimY.I. MoonS.H. JeonB.T. 2008 Effects of spent mushroom substrates addition on eating behavior of growing Hanwoo Korean J. Grassl. Forage Sci 28 107 118
- LeeW.S. LeeB.S. LeeS.C. LeeS.S. LeeS.Y. LeeD.Y. HaJ.K. 2004a Effects of rice straw and rice hull supplement on rumination and chewing behavior in hanwoo steers Korean J. Anim. Sci 46 49 54
- LeeW.S. LeeB.S. OhY.K. KangS.W. LeeS.S. HaJ.K. 2004b Effects of concentrate to roughage ratios on duration and frequencies of rumination and chewing in Hanwoo steers Korean J. Anim. Sci 46 55 60
- LicitraG. HernandesT.M. Van SoestP. J. 1996 Standardization of procedures for nitrogen fractionation of ruminants feeds Anim. Feed Sci. Tech 57 347 358
- LuoJ. SahluT. CameronM. GoetschetA.L. 2000 Growth of Spanish, Boer x Angora and Boer x Spanish goat kids fed milk replacer Small Ruminant Res 36 189 194
- MacedoC.A.B. MizubutiI.Y. MoreiraF.B. PereiraE.S. RibeiroE.L.A. RochaM.A.R. RamosB.M.O. MoriR.M. PintoA.P. AlvesT.C. CasimiroT.R. 2007 Comportamento ingestivo de ovinos recebendo dietas com diferentes níveis de bagaço de laranja em substituição à silagem de sorgo na ração Rev. Bras. Zootecn 36 1910 1916
- MahgoubO. LuC.D. EarlyR.J. 2000 Effects of dietary energy density on feed intake, body weight gain and carcass chemical composition of Omani growing lambs Small Ruminant Res 37 35 42
- McManusC. PaivaS.R. AraújoR.O. 2010 Genetics and breeding of sheep in Brazil Rev. Bras. Zootecn 39 236 246
- MedeirosG.R. CarvalhoF.F.R. FerreiraM.A. BatistaA.M.V. AlvesK.S. Souto Maior JuniorR.J. AlmeidaS.C. 2007 Efeito dos níveis de concentrado sobre o desempenho de ovinos Morada Nova em confinmento Rev. Bras. Zootecn 36 1162 1171
- MendesC.Q. TurinoV.F. SusinI. PiresA.V. MoraisJ.B. GentilR.S. 2010 Comportamento ingestivo de cordeiros e digestibilidade dos nutrientes de dietas contendo alta proporção de concentrado e diferentes fontes de fibra em detergente neutro Rev. Bras. Zootecn 39 594 600
- MertensD.R. 1987 Predicting intake and digestibility using mathematical models of ruminal function J. Anim. Sci 64 1548 1558
- MertensD.R. 1997 Creating a system for meeting the fiber requirements of dairy cows J. Dairy Sci 80 1463 1481
- MertensD.R. 2002 Gravimetric determination of amylase-treated neutral detergent fibre in feeds with refluxing beakers or crucibles: collaborative study J. AOAC Int 85 1217 1240
- National Research Council 2000 Nutrient Requirements of Beef Cattle 8th ed National Academy Press Washington, DC, USA
- National Research Council 2007 Nutrient Requirements of Sheep National Academic Press Washington, DC, USA
- PaulinoM.F. DetmannE. ValenteE.E.L. BarrosL.V. 2008 Nutrição de bovinos em pastejo 131 170 Proc. 4th Nat. Symp. on Strategic Management of Pasture Viçosa, Brazil
- PereiraE.S. MizubutiI.Y. RibeiroE.L.A. VillarroelA.B.S. PimentelP.G. 2009 Consumo, digestibilidade aparente dos nutrientes e comportamento ingestivo de bovinos da raça Holandesa alimentados com dietas contendo feno de capim-tifton 85 com diversos tamanhos de partícula Rev. Bras. Zootecn 38 190 195
- PimentelP.G. PereiraE.S. QueirozA.C. MizubutiI.Y. Regadas FilhoJ.G.L. MaiaI.S.G. 2011 Intake, apparent nutrient digestibility and ingestive behavior of sheep fed cashew nut meal Rev. Bras. Zootecn 40 1128 1133
- PolliV.A. RestleJ. SennaD.B. AlmeidaS.R.S. 1996 Aspectos relativos à ruminação de bovinos e bubalinos em regime de confinamento Rev. Bras. Zootecn 25 987 993
- PoppiD.P. HendricksenR.E. MinsonD.J. 1985 The relative resistance to escape of leaf and stem particles from the rumen of cattle and sheep J. Agr. Sci 105 9 14
- ProvenzaF.D. 1995 Role of learning in food preferences of ruminants: Greenhalgh and Reid revisited 233 247 Proc. 8th Int. Symp. on Ruminant Physiology Logan, UT, USA
- SAS 2003 SAS System for Windows, Release 9.1 SAS Inst., Inc. Cary, NC, USA
- SniffenC.J. O'ConnorD.J. Van SoestP.J. FoxD.G. RussellJ.B. 1992 A Net Carbohydrate and Protein System for Evaluating Cattle Diets: Carbohydrate and Protein Availability J. Anim. Sci 70 3562 3577
- TurinoV.F. 2003 Substituição da fibra em detergente neutro (FDN) do bagaço de cana-de-açúcar in natura pela FDN de casca de soja em dietas contendo alta proporção de concentrados para cordeiros confinados Degree Diss Universidade de São Paulo Brazil
- Van SoestP.J. RobertsonJ.B. LewisB.A. 1991 Methods for dietary fiber, neutral-detergent fiber and nonstarch polysaccharides in relation to animal nutrition J. Dairy Sci 74 3583 3597
- WeissW.P. 1999 Energy prediction equations for ruminant feeds 176 185 Proc. Cornell Nutrition Conf. for Feed Manufacturers, Cornell University Ithaca, NY, USA