
Abstract
Brasileiro de Hipismo (BH) represents an open population that permits genetic input from breeds recognised by the World Breeding Federation for Sport Horses. The aim of this study was the genetic characterisation of the population as the first of a series aiming at the consolidating the current selection programme. The Brazilian Association of the Brazilian Sport Horse Breeders provided genealogical data from 1977 to 2011. A data bank containing 34,393 horses was analysed for population genetic parameters including: individual inbreeding (F), effective population size (Ne), effective number of founders (fe), effective number of ancestors (fa), and number of founder genome equivalents (fg). Analysis were run twice: for animals born until 1995 (PREVIOUS) and for those born in the last 15 years (15YEARS). Ne estimated via paired increase in coancestry consisted of 188.59 (±3.24) animals. fe was 466 and 222 for PREVIOUS and 15YEARS, respectively. Ancestors were represented by fa of 274 and 129. Mean F for 15YEARS was 0.6%; 1444 animals were inbred, with mean inbreeding of 3.33%. The difference found in fa and fe parameters, between PREVIOUS and 15YEARS, indicated the loss of original alleles. The increase in contribution from some founders represents the breeder’s preference for a few horses, without the negative effect of high inbreeding levels. Brasileiro de Hipismo genetic variability agrees with its large based formation history, and should allow for genetic gain of heritable traits through selection.
Introduction
Horses (Equus caballus) from the Brasileiro de Hipismo (BH) breed are selected for showjumping, dressage and eventing, and the breed is recognised by the World Breeding Federation for Sport Horses (WBFSH). The herd book represents an open population that allows genetic input from any breed recognised by the WBFSH, the Thoroughbred (THB), Arabian and their crosses. The Brazilian Association of the Brazilian Sport Horse Breeders (ABCCH) is responsible for the genealogical registry of BH horses. The breed began by crossing national base mares (BASE) and stallions from founder breeds. Base mares, animals with known genealogy or not, were chosen as they were considered to be endowed with desirable morphological and functional characteristics and able to generate horses that would fit to the breed standard (ABCCH, Citation2011). New BASE are no longer registered.
Dias et al. (Citation2000) analysed genealogical data of BH registered between July 1977 and September 1998. The authors described the breed composition in the data bank as being 23% BH horses; 22% with unknown genealogy, 15% THB; 8% Hanoverian; 5% Westfalen; 5% Holsteiner and 4% Trakehner, the latter all European (mainly German) breeds of horses known for their success in sport competitions. These authors concluded that the great number of founder breeds contributed to the estimated inbreeding coefficient being close to zero and effective population size (Ne) of 253 animals; indicating high genetic variability.
The study of genetic variability in this Brazilian equine population with great sport potential and international importance is relevant for the maintenance and genetic improvement of the national sport horse. The aim of this study was the genetic characterisation of the current BH population, by estimating indicators of genetic variability and describing the contribution of the most influential ancestors.
Materials and methods
The ABCCH provided all genealogical data from its foundation in July 1977, until June 2011. The original data bank, containing 28,599 animals, was expanded when accounting for animals registered as parents to 5850 horses. Widely known, genetic relations of some stallions with large progeny numbers from founder breeds were also included. Such information was collected from the breed’s web site (http://brasileirodehipismo.com.br/site/html/stbconsgenealogia.asp) and resulted in 163 genetic relations added to data bank. The final data bank contained 34,393 horses and included: animal identification, sire, dam, sex, breed, and stud of origin.
Aiming at a better characterisation of the recent development of this breed, analyses were run twice considering two different subpopulations: i) all BH animals born until 1996, totalising 10,588 horses and entitled PREVIOUS; ii) all BH born in last 15 years, containing 8353 horses, named 15YEARS.
Population genetic parameters estimated in the present study included: individual inbreeding (F) (Wright, Citation1931), computed following Meuwissen and Luo (Citation1992); effective population size (Ne), effective number of founders (fe), effective number of ancestors (fa) and number of founder genome equivalents (fg). In order to make inferences on genetic contribution of founder breeds to total diversity, breeds were classified according to their geographical origin, resulting in the following groups: BH (19,031 animals), GERMAN (1958), FRENCH (329), DUTCH (298), and BELGIAN (271). Base mares formed the BASE (4639) group. Thoroughbred, Arabians and their crosses formed the HOT group (1554). Animals with missing breed information were classified as NOTINFORMED (5970). Among NOTINFORMED animals, were those 5850 parents included as animals in the data bank, as mentioned above. The 50 breeds from other origins had minor individual numbers, and were grouped together under the name OTHER (343). Genetic contributions were computed following Caballero and Toro (Citation2002). Generation intervals were computed as the average age of parents at the birth of their progeny kept for reproduction according to James (Citation1972). The final data bank was formed by 55% (19,012) BH animals, 17% without breed specification (5970), 14% base animals (4639), 4% THB (1461), 2% (725) Westfalen and 2% (641) were Holsteiner horses. The remaining animals were in smaller proportion (<1%) of the other 47 founder breeds. Total number of stallions and breeding mares were 2886 and 10,372, respectively.
According to Kimura and Crow (Citation1963) the Ne is defined as the size of an idealised population with the same amount of inbreeding or random gene frequency drift as the population under study. In the present study, Ne was estimated via individual increase in inbreeding per generation (͞Ne), as proposed by Gutiérrez et al. (Citation2008); and also by average increase in coancestry for all pairs of individuals (͞Nec) (Cervantes et al., Citation2011). Other estimates of Ne were calculated by computing the regression coefficient of the individual inbreeding coefficient over: i) the number of full generations traced; ii) the maximum number of generations traced; and iii) the equivalent complete generations (Maignel et al., Citation1996). Effective number of founders (fe) is defined as the number of equally contributing founders that would be expected to produce the same genetic diversity as in the population under study and was computed as: fe = 1/Σfk-1 q2k; where qk is the probability of gene origin of the k ancestor. Parameter fa is the minimum number of ancestors, not necessarily founders, explaining the complete genetic diversity in a population, and was computed as fe = 1/Σaj-1 q2j; where qj is the marginal contribution of an ancestor j (Gutiérrez and Goyache, Citation2010). The parameter fg is defined as the number of founders that would be expected to produce the same genetic diversity as the population under study if the founders were equally represented and no loss of alleles occurred, according to Caballero and Toro (Citation2000).
Animals in the data bank were ordered and recoded using personal Fortran programmes, and data was analysed using the ENDOG software v 4.8 (Gutiérrez and Goyache, Citation2005).
Results and discussion
The number of BH horses registered annually started at 75, in 1977, and increased considerably, reaching 975 animals in 1990. Since then the number has been relatively stable until 2009, varying between 454 and 768 foals per year. During 2010 and until June 2011 a drop in registry can be noted (). This was expected, as animals born in this period may not have been registered at the time of data collection. Since breed formation, the total number of BH breeders was 794, with the average number of foals registered equal to 31.63 (±103.67). Breeders were characterised according to their contribution with stallions to the total population. demonstrates that all breeding farms (794) brought in male genetic material, and some of them never produced any stallions (394). Among those breeders that produced stallions (400), only 59 sired their own mares, in small proportions (12%). This fact might have contributed to inbreeding control. Total number of reproductive mares that foaled BH horses was 6583; averaging 2.89 (±2.40) foals per mare. In total, 991 stallions were used as sires for BH horses since 1977, with mean number of foals per sire equal to 19.16 (±37.40). Generation intervals for 15YEARS are shown in . High generation intervals were found as they were expected in horse populations (Hamann and Distl, Citation2008; Dubois, Citation2007; Valera et al. Citation2005). Dias et al. (Citation2000) reported slightly inferior intervals for male (10.40 years) and female (9.30 years) generations in BH horses born until 1998. Pedigree completeness was estimated by generation for the whole data bank, BH horses from the PREVIOUS and 15YEARS subpopulation. As expected, those animals born in the last 15 years presented higher pedigree completeness than other subpopulations (). Completeness for PREVIOUS period was equal to 99.80, 78.60, 28.10 and 6.70%; from first to fourth parental generation, respectively. Recent pedigree completeness (15YEARS) was equal to 99.99, 98.3, 69.9 and 37.8%, following the same order. A general tendency for sire lines being more complete than dam lines was observed in both periods.
Detailed population genetic parameters for BH in PREVIOUS and 15YEARS subpopulations are shown in . A large decrease in number of ancestors contributing to the 15YEARS period (1736) was seen compared to PREVIOUS (3316 ancestors). Comparing both periods, the same reducing pattern occurred in fe (466 for PREVIOUS and 222 for 15YEARS), fa (274 and 129) and number of ancestors explaining 50% of genetic variation (126 and 61), respectively. The number of founder genome equivalents (fg) also presented similar declining pattern; as for the whole BH subpopulation (PREVIOUS) it was 224.77, while for the 15YEARS subpopulation was equal to 81.07.
The mean computed inbreeding for all animals in the data bank (0.24%) was slightly higher than the expected by unbalancing of founders contribution (0.05%). When considering only PREVIOUS BH horses the mean inbreeding was 0.15%. Mean inbreeding for 15YEARS was 0.6%. In this period, 1444 animals presented inbreeding greater than to 0.5%, with average inbreeding of 3.33%. Maximum inbreeding coefficient was equal to 27%. The maximum computed inbreeding was 32.81% in animals outside the BH population (). Dias et al. (Citation2000) reported mean inbreeding for 151 inbred animals equal to 0.12% for animals born until 1998. Among animals born in the last 15 years, a greater number of inbred animals were found as well as greater inbreeding average for inbred animals. Such tendency of increasing BH mean inbreeding with time was also illustrated by and has been seen in other horse populations (Maciel et al., Citation2014). This is expected as later generations have higher pedigree completeness.
Estimates of effective population size (Ne) via increase in inbreeding by generation are presented in . The estimate based on mean equivalent generations was similar to those obtained by individual increase in inbreeding (͞Ne), equal to 205.78 (±58.18); and via paired increase in coancestry (͞Nec), equal to 188.59 (±3.24) animals. Greater effective population size was obtained for older generations of BH horses from a smaller data bank, and shallower pedigrees (Dias et al. Citation2000).
Dias et al. (Citation2000) described a data bank containing 19,303 horses. Among those, 11,508 were BH born until 1998. Those animals are mostly the same considered here in the PREVIOUS subpopulation, but a smaller data bank was included in their study. Dias et al. (Citation2000) described only generation intervals (9.90 years), effective population size (253 animals) and inbreeding. Considering the 151 inbred animals, average inbreeding was 0.12%, ranging between 0.03 and 0.25%. Maximum number of generations reported was 3.10 for one animal. Among BH horses, 59.7% were at the first generation. Completeness of pedigree information was responsible for the small differences found between Dias et al. (Citation2000) and the PREVIOUS subpopulation reported here.
Animals born in the last 15 years constitute most of the BH population acting as athletes in sport competitions today. The present analysis complements that described by Dias et al. (Citation2000) and is the first to describe detailed population genetic parameters for recent (15YEARS) and PREVIOUS BH subpopulations. Classification of Studs (), detailed generation intervals (), parameters fa, fe and fg (), recent estimates of Ne, and origin of BH genes (), constitute a more profound analysis than the anterior study.
Genetic diversity in BH population is demonstrated by its estimation of number of ancestors, fe, fa, fg and number of ancestors explaining 50% of genetic variation. According to Boichard et al. (Citation1997) such parameters are less sensitive to pedigree length than inbreeding coefficients and Ne estimated by increase in inbreeding.
The decreasing pattern observed in each parameter, when PREVIOUS and 15YEARS were compared, indicated a loss of founder alleles. Similar bottlenecks were described for Trakehner (Teegen et al., Citation2009) and Selle Français (SF) (Dubois, Citation2007). Other evidence of allelic loss in BH was revealed trough the differences found between parameters fe and fa in both moments. The ratio between them (fe/fa) equals 1.70 and 1.72 for the PREVIOUS and 15YEARS, respectively. A much higher ratio (3.15) was found for Hanoverian horse, where the loss of genetic diversity was attributed to selection (Hamann and Distl, Citation2008).
The greater number of effective founders than effective ancestors is possibly due to breeder’s preferential use of some genetic lines, leading to an unbalance in founder contribution. Such preferential reproduction was verified specially through the decrease in reproduction of THB stallions, as well as THB and BASE mares ( and ).
Geographical origin of founder breeds had their genetic contribution to total diversity of 15YEARS presented in . Recent BH population descends mainly from German Sport Horse breeds such as Holstein, Westphalia and Hanoverian. Registration date of each BASE mare was not available, but birth dates suggested the period of their acceptance for reproduction. Few (13.99%) from the total of registered BASE mares were born before 1977. Most of them (59.37%) were born during the 70’s and a smaller proportion (26.26%) during the 80’s. Only 18 (0.39%) BASE mares were born in the 90’s and the last one in 1992. Therefore, a decrease in BASE’s contribution was expected with their ageing. The data bank contained many horses not considered BH. Those animals complemented pedigree information, resulting in the higher pedigree completeness described for BH horses, especially for the 15YEARS period. Only base mares were, in fact, of unknown genealogy. Therefore, pedigree completeness could be increased with complementation of founder breed data, especially for those 4516 animals registered as parents that were initially included in the data bank as animals.
Pedigree complementation is a long and exhausting job, demanding cooperation among many breeding organisations. Still, the addition of complete pedigree information of each imported animal used for reproduction is of great relevance for population genetic studies (Thorén Hellsten et al., Citation2008; Cervantes et al., Citation2009). Brasileiro de Hipismo breeders seem to have followed the world tendency of sport horse breeding organizations in importing genetic material. Recent developments in reproductive techniques have increased this global phenomenon (Bruns et al., Citation2004; Koenen and Aldridge, Citation2002). Koenen et al. (Citation2004) illustrated such exchange studying the geographical origin of warmblood stallions used in some important European sport horse studbooks. The authors demonstrated that Germany was the main origin of the sires used in Denmark, Sweden and Netherlands. On the other hand, some countries like France and Ireland showed larger national recruitment of stallions. The breeder’s preference for foreign blood, especially German, is once more demonstrated in . The small proportion of BH stallions used as parents in the last 15 years, and the large amount of births from BH mares demonstrated that genetic input happened mainly through either imported stallions or semen.
The high proportion of mating involving stallions without breed specification () refers to imported semen. As described by Koenen et al. (Citation2004), one European sport horse stallion may be born under one studbook and be accepted for reproduction in another. In those cases the breed specification becomes a difficult task, once the horse may produce foals registered in different breeds. Such genetic input occurred mainly through stallions, probably due to reproductive facilities.
Animals from 50 founder breeds were found in the BH data bank. At least 37.52% of recent BH genes are from German sport horse breeds. A high number (4636) of base mares of unknown genealogy are registered in data bank, though their genetic contribution for 15YEARS was estimated at 7.18%, probably due to selection.
The use of BH, THB and other founder breeds as stallions during the whole period considered was illustrated in . Other breeds were represented mainly by European founder breeds recognised by the WBFSH, as each animal was approved by ABCCH. The continuous genetic income and a decrease in use of THB horses as sires were evident.
The increase in reproduction of BH mares was demonstrated in , as well as a growing use of other breeds recognised by the WBFSH, possibly due to embryo transfer. Base mares were accepted initially when few imported females were available. As time passed, those animals aged and their use decreased. Thoroughbred mare use was reduced probably by the growing number of available BH mares. In the last 15 years, foals have been registered mainly as products of BH and European warmblood mares. The change in stallion and reproductive mare pattern of reproduction were clearly responsible for those bottlenecks indicated by genetic population parameters.
Gutierrez and Goyache (Citation2010) reported that, when information is scarce, Ne estimations based on maximum and minimum generations traced could be considered as the upper and lower limits of the parameter in the analysed population. Estimates based on mean equivalent generations should be close to real value of Ne. Parameter Ne of BH population obtained via increase in inbreeding by mean equivalent generation (223.39), (͞Ne), (205.77; ±58.18) and (͞Nec), (188.59; ±3.24) represented the effective population size for BH population.
The Spanish Sport Horse (SSH) is a recent breed, founded in 2002, and many aspects in common with BH were reported regarding its genetic formation (Cervantes et al. Citation2009; Bartolomé et al. Citation2011). Cervantes et al. (Citation2009) analysed genealogy for three Spanish horse breeds using the breed studbook data and information from the parental breeds, describing important differences in parameter estimation. Spanish Sport Horse had its effective population size estimated at 47 with breed exclusive data (until 2004) and 135.5 when data from Spanish Arab, Spanish Purebreed, THB, and Hispano-Arab were included. Bartolomé et al. (Citation2011) analysed SSH animals born between 2005 and 2009, finding Ne equal to 225 (±72.4). Parameters fe (963); fa (407) and fg (245.3) were higher than those described here for BH breed.
Brasileiro de Hipismo Ne estimates agree with that described for Creole (Ne= 214.46) by Maciel et al. (Citation2014); although the parameter was inferior to Ne described for Hanoverian (Ne= 372.34) by Hamann and Distl (Citation2008). Closed herd books tend to present lower values of Ne, as reported by Cervantes et al. (Citation2009) for the Arab-derived Spanish breeds (109.5 to 135.5), and by Teegen et al. (Citation2009) for the Trakenher breed (144 to 150). Locally adapted breeds with limited environmental range such as the Brazilian Pantaneiro, whose herd book was also started in the 1970s, also show a much lower Ne (25) (McManus et al., Citation2013). Parameters fe, fa, and number of ancestors contributing to 50% of genes estimated for BH recent population (15YEARS), indicated larger genetic diversity than described for important BH founder breeds, such as SF (Dubois, Citation2007) and Hanoverian (Hamann and Distl, Citation2008). Less diversity was described for the closed herd populations: Lipizzan (Zechner et al., Citation2002), Andalusian (Valera et al., Citation2005), and Spanish Arab (Cervantes et al., Citation2008, Citation2009).
The low mean inbreeding estimated for BH horses was due to its large population size and the large number of founder animals from genetically diverse breeds. As expected, information on BH pedigree is increasing, along with mean annual inbreeding (). Recent BH animals (15YEARS) had mean equivalent generations equal to 3.25; while for PREVIOUS period it was estimated as 2.15. Deeper pedigree data have been described in older breeds. Zechner et al. (Citation2002) described an average of 15.22 complete generations for Lipizzan horses. Andalusian (Valera et al., Citation2005), Hanoverian (Hamann and Distl, Citation2008), and last decade Spanish Arab (Cervantes et al., Citation2008) were reported to have mean equivalent generations of 8.26; 8.43 and 7.9 respectively. Shallower pedigree was found in younger breeds such as SSH (4.29) by Bartolomé et al. (Citation2011) and Creole (4.62) by Maciel et al. (Citation2014). Horse breeds with small population size and closed studbooks, like Andalusian (mean inbreeding 8%) reported by Valera et al., (Citation2005); Spanish Arab (recent mean inbreeding of 10%) described by Cervantes et al. (Citation2008); and Lipizzan (11%) by Zechner et al. (Citation2002) were expected to show higher endogamy coefficients. Open populations tended to present lower mean inbreeding: Hanoverian (1.33%) described by Hamann and Distl (Citation2008); SF (1.40%) by Dubois (Citation2007) and SSH (0.66%) by Bartolomé et al. (Citation2011). The twenty most influential ancestors were listed along with their genetic contribution to the 15YEARS period (), and mentioned in the text with capital letters. Some ancestors, such as the THB LADYKILLER (recent contribution of 2.8%) and its progeny LANDGRAF I (2%) had their contribution largely increased in the last 15 years. Both of them have been represented in BH population directly by the Holstein stallions Landritter and Lorado who had 495 and 285 BH foals registered, respectively. The THB stallion RANTZAU (recent contribution of 1.98%), and the SF IBRAHIM (1.4%) were represented mainly by their respective progenies: Cor de la Bryere and Alme GIRONDINE (1.2%), dam of Alme, is also listed among the most contributing animals for recent BH population. QUENOTTE (0.9%), dam of Cor de la Bryere, brings her ancestors THB genes: FURIOSO (0.8%) and PRECIPITATION (0.8%).
Dubois (Citation2007) reported that at least 50% of SF gene pool originated in THB. The author also described the major ancestors of SF and their contribution to the breed. Although genetic contribution of French sport horses to recent BH generation was only 5.69%, some of the ancestors described for SF were found to be major contributors to BH as well. IBRAHIM, for example, contributed to 8.70% of genetic constitution of SF (Dubois, Citation2007) and was responsible for 1.44% of BH diversity. FURIOSO, contributed to 3.8% of SF and 0.81% of BH. RANTZAU, who participated in 2.3% of SF, and GIRONDINE, with 1.6% influence in SF; kept similar contribution to the most recent BH generation.
Common ancestors were also reported in the Hanoverian breed. Cor de la Bryere, LADYKILLER and Furioso II were reported to contribute with 1.31, 1.29 and 1.28% of Hanoverian stallion’s genes by Hamann and Distl (Citation2008). Ramiro contributed with 1.75% of Hanoverian stallions gene pool, and is an important son of RAIMOND, bringing RAMZES into Hanoverian breed. Some other ancient horses had their contribution increased in recent BH subpopulation. Many other ancient horses had their genetic contribution increased in BH recently. The Anglo Arabian stallion RAMZES left a major contributor to BH population, and increased in the past 15 years, from 0.57% (whole data) to 1.3% (15YEARS). Similar phenomenon occurred to PERSEUS and ST.PR.ST.DUELA were represented by their progeny the Westphalian Pilatus, who had 10 descendants in the dataset. Both increased contribution to the BH breed recently.
Considering that fifty breeds appear in BH dataset and little complementation was manually inserted, other genetic relations are certainly missing in this study. Therefore, inbreeding coefficients and genetic contributions estimated can be considered as minimum contributions of each founder for BH breed, and some of them might be underestimated by missing data effect. As demonstrated, similarities exist between BH and European sport horse breeds. Genetic flow is still happening from founder breeds to the Brazilian population. The recent increase in the contribution of ancient ancestors was accompanied by a tendency of increasing inbreeding over years, suggesting that the same lines were brought once again to BH population. These facts indicate that the breeder’s preference for a few genetic lines might lead to further increase in inbreeding.
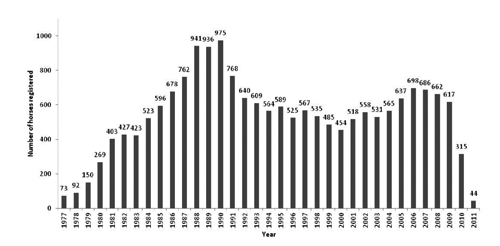
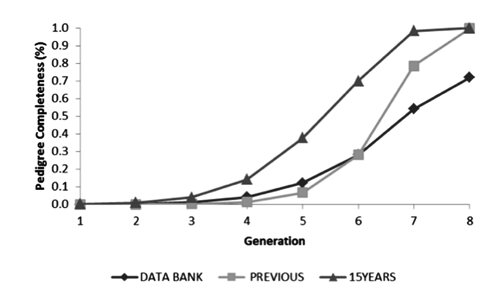
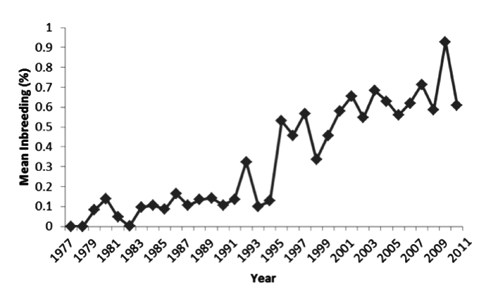
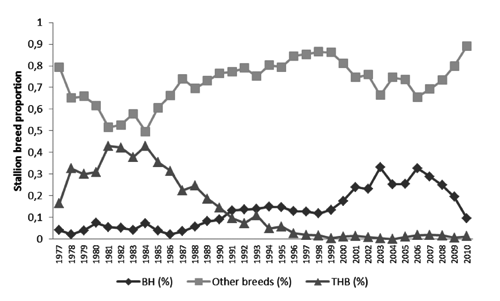
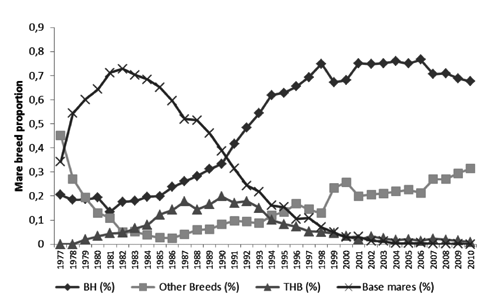
Table 1. Classification of studs, considering their contribution with stallions to the population.
Table 2. Generation intervals and mean age of parent at birth of their offspring for Brasileiro de Hipismo subpopulation born in the last 15 years.
Table 3. Population genetic parameters estimated for the Brasileiro de Hipismo population until 1995 and for animals born in the last 15 years.
Table 4. Measures of inbreeding for all horses in data bank and for those born in the last 15 years.
Table 5. Increase in inbreeding per generation and effective population size estimated via mean: maximum, complete and equivalent generations.
Table 6. Genetic contribution of founder breeds to total diversity of animals born in the last 15 years.
Table 7. Most used breeds as stallion and mare along with each proportion of total foaling of the last 15 years.
Table 8. Most influential ancestors to Brasileiro de Hipismo horses born in the last 15 years and their genetic contribution to 15YEARS and PREVIOUS subpopulations, sex and progeny of greater influence to the breed.
Conclusions
The genetic variability has decreased in BH breed, mainly due to reduction in use of THB breed and extinction of base mares. Such loss of founder alleles did not lead to dangerous inbreeding levels. On the other hand, a tendency of increasing BH inbreeding was demonstrated as well as the raising contributions of few famous ancient animals. Thus, the search for a limited number of famous families from the parental horse breeds might lead to future limitation in genetic variation.
Acknowledgements
Thanks are due to the Brazilian Association of the Brazilian Sport Horse Breeders (ABCCH) for providing the data.
Related Research Data
References
- ABCCH, 2011. Regulamento do stud book Brasileiro do cavalo de Hipismo. Available from: http://brasileirodehipismo.com.br/site/html/stbregulamento.asp
- BartoloméE. CervantesI. ValeraM. GutiérrezJ.P. 2011. Influence of foreign breeds on the genetic structure of the Spanish Sport Horse population. Livest. Sci. 142: 70-79.
- BoichardD. MaignelL. VerrierE., 1997. The value of using probabilities of gene origin to measure genetic variability in a population. Genet. Sel. Evol. 29: 5-23.
- BrunsE.M. RicardA. KoenenE., 2004. Interstallion: on the way to an international genetic evaluation of sport horses. Page 326 in Proc. 55th Ann. Meeting Eur. Assoc. Anim. Prod., Bled, Slovenia.
- CaballeroA. ToroM.A., 2000. Interrelations between effective population size and other pedigree tools for the management of conserved populations. Genet. Res. 75: 331-343.
- CervantesI. GoyacheF. MolinaA. ValeraM. GutiérrezJ.P., 2011. Estimation of effective population size from the rate of coancestry in pedigreed populations. J. Anim. Breed Genet. 128: 56-63.
- CervantesI. GutiérrezJ.P. MolinaA. GoyacheF. ValeraM., 2009. Genealogical analyses in open populations: the case of three Arab-derived Spanish horse breeds. J. Anim. Breed Genet. 126: 335-347.
- CervantesI. MolinaA. GoyacheF. GutiérrezJ.P. ValeraM., 2008. Population history and genetic variability in the Spanish Arab Horse assessed via pedigree analysis. Livest. Sci. 113: 24-33.
- DiasI.M.G. BergmannJ.A.G. RezendeA.C.C. CastroG.H.F., 2000. Formação e estrutura populacional do equino Brasileiro de Hipismo. Arq. Bras. Med. Vet. Zoo. 52: 647-654.
- DuboisC., 2007. Modélisation des programmes de sélection dans l’élevage du cheval de sport français. PhD Diss., AgroParisTech, Paris, France.
- GutiérrezJ.P. GoyacheP., 2005. A note on ENDOG: a computer program for analysing pedigree information. J. Anim. Breed Genet. 122: 172-176.
- GutiérrezJ.P. GoyacheF., 2010. ENDOG v4.8: a computer program for monitoring genetic variability of populations using pedigree information. User’s guide. Departamento de Producción Animal - Área de Genetica y Reproducción Animal, Universidad Complutense de Madrid Publ., Madrid, Spain.
- GutiérrezJ.P. CervantesI. MolinaA. ValeraM. GoyacheF., 2008. Individual increase in inbreeding allows estimating realised effective sizes from pedigrees. Genet. Sel. Evol. 40: 359-378.
- HamannH. DistlO., 2008. Genetic variability in Hanoverian warmblood horses using pedigree analysis. J. Anim. Sci. 86:1503-1513.
- JamesJ., 1972. Computation of genetic contributions from pedigrees. Theor. Appl. Genet. 42: 272-273.
- KimuraM. CrowJ.F., 1963. The measurement of effective population number. Evolution 17: 279-288.
- KoenenE.P.C. AldridgeL.I., 2002. Testing and genetic evaluation of sport horses in an international perspective. pp 1-8 in Proc. 7th World Congr. Livest. Prod., Montpellier, France.
- KoenenE.P.C. AldridgeL.I. PhilipssonJ., 2004. An overview of breeding objectives for warmblood sport horses. Livest. Prod. Sci. 88: 77-84.
- MacielF.C. BertoliC.D. Braccini NetoJ. CobuciJ.A. PaivaS.A. McManusC., 2014. Population structure and genealogical analysis of the Brazilian Creole Horse. Anim. Genet. Res. 42: 1-11.
- MaignelL. BoichardD. VerrierE., 1996. Genetic variability of French dairy breeds estimated from pedigree information. Interbull Bull. 14: 49-54.
- McManusC. SantosS.A. DallagoB.S.L. PaivaS.R. MartinsR.F.S. Braccini NetoJ. MarquesP.R. AbreuU.G.P., 2013. Evaluation of conservation program for the Pantaneiro horse in Brazil. Rev. Bras. Zootecn. 42: 404-413.
- MeuwissenT.I. LuoZ., 1992. Computing inbreeding coefficients in large populations. Genet. Sel. Evol. 24: 305-313.
- TeegenR. EdelaC. ThallerG., 2009. Population structure of the Trakehner Horse breed. Animal 3: 6-15.
- Thorén HellstenE. JorjaniH. PhilipssonJ., 2008. Connectedness among five European sport horse populations. Livest. Sci. 118: 147-156.
- ValeraM. MolinaA. GutiérrezJ.P. GómezJ. GoyacheF., 2005. Pedigree analysis in the Andalusian horse: population structure, genetic variability and influence of the Carthusian strain. Livest. Prod. Sci. 95: 57-66.
- WrightS., 1931. Evolution in mendelian populations. Genetics 16: 97-158.
- ZechnerP. SölknerJ. BodoI. DrumlT. BaumungR. AchmannR. MartiE. HabeF. BremE.G., 2002. Analysis of diversity and population structure in the Lipizzan horse breed based on pedigree information. Livest. Prod. Sci. 77: 137-146.