Abstract
In spite of attractive attributes, diatoms are underrepresented in research and literature related to the development of microalgal biofuels. Diatoms are highly diverse and have substantial evolutionarily-based differences in cellular organization and metabolic processes relative to chlorophytes. Diatoms have tremendous ecological success, with typically higher productivity than other algal classes, which may relate to cellular factors discussed in this review. Diatoms can accumulate lipid equivalently or to a greater extent than other algal classes, and can rapidly induce triacylglycerol under Si limitation, avoiding the detrimental effects on photosynthesis, gene expression and protein content associated with N limitation. Diatoms have been grown on production scales for aquaculture for decades, produce value-added products and are amenable to omic and genetic manipulation approaches. In this article, we highlight beneficial attributes and address potential concerns of diatoms as biofuels research and production organisms, and encourage a greater emphasis on their development in the biofuels arena.
(A) Thalassiosira pseudonana, (B)Cocconeis sp., (C)Lampriscus sp., (D)Gyrosigma balticum, (E)Cyclotella cryptica, (F)Nitzschia sp., (G)Thalassiosira weissflogii and (H)Achnanthes sp.

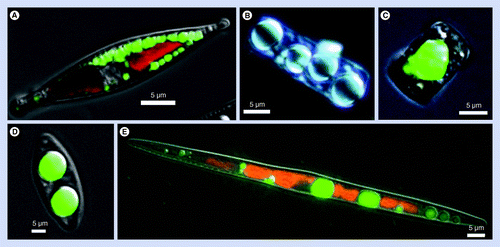
(A) Nitzschia curvilineata, (B)Cyclotella cryptica, (C)C. cryptica, (D)Nitzschia alba and (E)Nitzschia filiformis. Green fluorescence is BODIPY staining of neutral lipids, red is chlorophyll autofluorescence. (B) is a differential interference contrast image with no fluorescence.
Branch lengths do not correspond to phylogenetic distances. Red arrow locates diatoms and green arrow locates chlorophytes.
Reproduced with permission from Citation[176].
![Figure 3. Unrooted consensus phylogeny of photosynthetic eukaryotes and derived taxa.Branch lengths do not correspond to phylogenetic distances. Red arrow locates diatoms and green arrow locates chlorophytes.Reproduced with permission from Citation[176].](/cms/asset/fdffc15d-9467-4781-9ecb-768acad02eee/tbfu_a_10815964_f0003.gif)
Cultures were harvested from exponential phase and inoculated into fresh medium for the various treatments. The nocodazole treatment was done in a separate experiment from Si and N.
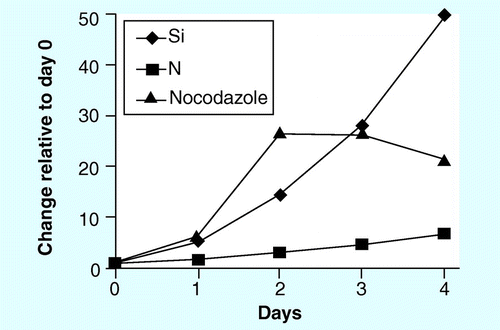
Incremental amounts of silicic acid were added to the cultures at day 1 and growth followed thereafter. On day 7 when all cultures were limited, the ratio of cell numbers to starting concentration were in exact correspondence.
A frequently stated reason for choosing unicellular microalgae as biofuel-production organisms is that they are more efficient than vascular plants at turning light and nutrients into valuable products. The economics of algal biofuels is critically dependent on productivity, and one can apply the same criterion and ask the question: among the microalgae, are certain classes more productive? Before answering that question, we need to acknowledge that within a given class of algae, there will be differences in measured productivity values when comparing individual species. These differences can stem from intrinsic factors related to their cell biology or from growth conditions at the time of sampling, which can involve both environmental and ecological variables. Comparing productivity in controlled culture conditions has value, because it reflects what is possible, but it also encompasses biases due to the fact that conditions are never exactly the same in two culture systems. Indications are that algal biofuels will be developed on a regional basis in a wide variety of environments Citation[1]. Another consideration is the utility of comparing maximum productivity values. In a production system operating for a good portion of the year, it would be far more desirable to have strains that are consistently productive under a variety of environmental conditions than strains that have higher productivity under optimal conditions for a shorter period of time. A final consideration in productivity comparisons is that if the number of studies is relatively small, which is usually the case, poor statistical comparisons result. An alternative to measuring productivity values in culture is to evaluate an algal class’ productivity on the basis of its abundance in nature in a variety of environments. This approach may directly relate to biofuel production systems that would be deployed in different environments and provide robust statistics, since the sample size is large.
Diatoms are among the most productive and environmentally flexible eukaryotic microalgae on the planet. They are estimated to be responsible for 20% of global carbon fixation and have become the dominant primary producers in the ocean Citation[2,3]. In addition to being highly abundant, they are also highly diverse, with estimates of over 100,000 species Citation[4]. Their most distinctive cellular feature is their cell wall made of nanostructured silica , which is reproduced with fidelity through generations by genetically controlled assembly processes. Typical diatoms range in size from a few to a few hundred microns, with the majority of species between 10 and 50 µm.
Diatoms have characteristics, both conceptual and proven, which make them amenable to large-scale biofuel cultivation. In 1998, the Aquatic Species Program recommended a list of 50 microalgal strains, selected from a pool of over 3000 candidates, which held the most promise as biofuel-production organisms Citation[201]. Sixty percent of the selected strains were diatoms, which were chosen based on criteria such as high growth rates and lipid yields, tolerance of harsh environmental conditions, and performance in large-scale cultures Citation[201]. Diatoms are excellent lipid accumulators, and we frequently see cells similar to the ones shown in where a substantial portion of the cells’ volume is occupied by lipid droplets that accumulate rapidly, especially under Si limitation.
There is a puzzling underrepresentation of diatoms in the microalgal biofuels arena. Based on our survey of the literature and research experience, this is not because they are inferior to other classes of algae. The purpose of this article is to highlight attributes of diatoms that make them highly desirable as organisms for biofuel research and production, and to address potential concerns and misconceptions about diatoms that we have encountered in the literature and during dialog with colleagues over the past several years.
Evolutionary aspects of diatoms
Diatoms are members of the Stramenopile or heterokont class of algae , which are highly different from, and have a more complex evolutionary history than, green algae and vascular plants Citation[5]. Common to all photosynthetic eukaryotes was a primary endosymbiosis event where a heterotrophic eukaryote engulfed or was invaded by a cyanobacterium. Most of the cyanobacterial genes were transferred to the nucleus as the cyanobacterium evolved into the chloroplast. The progenitor plant cell resulting from primary endosymbiosis gave rise to the glaucophytes, red algae, green algae and plants. A secondary endosymbiosis later occurred between a different heterotrophic eukaryote and a red alga. The red algal endosymbiont was the progenitor of the plastids found in Stramenopiles, a group that includes diatoms, brown macroalgae and plant parasites . In addition to diatoms, prospective biofuel production organisms such as Nannochloropsis are in the Stramenopiles.
The evolutionary story is a bit more complex for diatoms; evidence suggests that at some stage in the development of the progenitor plant cell, a Chlamydial invasion occurred Citation[6]. There is also evidence that green algal nuclear genes are present in the secondary endosymbiont Citation[7], suggesting an endosymbiotic episode with a green alga. Diatom genomes also contain abundant numbers (up to 5%) of genes derived from bacteria of various classes, with more than half of these genes being shared between two evolutionarily diverse diatoms, Thalassiosira pseudonana and Phaeodactylum tricornutumCitation[8]. Thus diatom genomes, and their resultant metabolomes, are a complex mixture of components derived from highly divergent sources Citation[8,9]. This is reflected in a unique combination of metabolic processes in diatoms, which combine both plant- and animal-like characteristics Citation[10]. Fundamental aspects of diatom metabolism differ substantially from other classes of algae Citation[11], and caution should be taken when making generalizations about algae based on studies on one organism.
The four major groups of diatoms (based on the features of their silica cell walls) are the radial centrics, the bipolar and multipolar centrics, the araphid pennates, and the raphid pennates; each group arose and diversified sequentially under decreasing CO2 levels during the Mesozoic era Citation[5]. Thus, their carbon concentrating and fixation systems evolved under substantially different conditions; for example, CO2 levels were, at one point, eight-times higher than present Citation[12]. It is reasonable to expect that changing environmental CO2 levels were drivers (at least in part) of diatom diversification and that differences in the physiology of the major groups may reflect this adaptation. Diatoms also adapted to grow and dominate in low-nutrient open ocean waters resulting from changing global geochemical conditions Citation[5]. Conversely, diatoms may have affected environmental conditions themselves; a rapid rise in diatom abundance during the Eocene may have resulted in a massive drawdown of atmospheric CO2 that facilitated global cooling Citation[13].
The fossil record indicates that over the Mesozoic era larger eukaryotic phytoplankton of the red algal lineage, which includes dinoflagellates, coccolithophorids and diatoms, displaced a large portion of other algae in the ocean, which were predominantly cyanobacteria and very small green algae Citation[14]. This suggests fundamental productivity advantages to the red algal line Citation[14], which will be discussed later. Enhanced sinking rates of the larger phytoplankton increased the burial of organic carbon in the continental margins and shallow seas, which generated most of our known petroleum reserves Citation[15,16], and reduced atmospheric CO2 levels. While some early reports suggested that diatoms were the major source of carbon for fossil fuels, dinoflagellates and coccolithophorids were the dominant types of phytoplankton during the major carbon export period Citation[14]. Currently, diatoms are responsible for a large fraction of the organic carbon buried on continental margins Citation[17] and are major contributors to nascent petroleum reserves.
Diatom productivity
Currently, in the oceans, diatoms are one of the most productive classes of microalgae. They are responsible for a substantial portion (40%) of marine primary productivity Citation[3] and most of the CO2 export (drawdown and sinking) from the atmosphere, of the order of 50% Citation[18]. The intrinsically high productivity of diatoms not only makes them major players in the global carbon cycle, but also suggests their utility as biofuel production organisms.
▪ Natural productivity
Diatoms are efficient at nutrient utilization and are entirely responsible for the initial nitrate uptake in the equatorial upwelling zone of the eastern Pacific Ocean Citation[18]. Other eukaryotic phytoplankton obtain their N through the action of grazers, which allows N from diatoms to be recycled Citation[18]. One of the contributing factors to diatom dominance is that they possess a large nutrient storage vacuole, which dinoflagellates and coccolithophorids lack Citation[19–21]. Vacuolization has important consequences in terms of productivity. Nutrient utilization efficiency is generally related to surface to volume ratio, and smaller cells tend to be more efficient for this reason. One way to compensate for a larger surface area is to store nutrients inside the cell in a vacuole, which in essence reduces the cytoplasmic volume and increases the surface area from which nutrients can be utilized by the cytoplasm Citation[19]. As long as extracellular nutrients are present at levels that minimize diffusive limitation to the cell surface, the gains in productivity in vacuolated cells are quite substantial Citation[19]. Thus, in nutrient-replete conditions, diatoms will hoard excess nutrients, storing enough nitrate to enable several cell divisions, and preventing the growth of other phytoplankton Citation[15]. In a survey of in situ growth rates of marine phytoplankton, maximum doubling rates between 2 and 4 per day were repeatedly measured for both pennate and centric diatoms, which were higher than those for other algae with corresponding sizes Citation[22]. Diatoms can even dominate under nutrient-limiting conditions; in one example a diatom species was shown to outcompete non-N-fixing cyanobacteria under low nitrate concentrations in a eutrophic lake Citation[23]. Diatoms consistently outcompete other eukaryotic phytoplankton under conditions of mixing and high turbulence, in which pulses of nutrients occur between short periods of quiescence Citation[24,25]. Furthermore, the relative dominance of diatoms in the oceans relates to geologic time periods of generally greater ocean turbulence Citation[26–29].
Diatoms also have a greater carbon fixing ability than other co-existing groups of microalgae, as shown from calculations of productivity per unit of crop carbon Citation[30]. This has not only been repeatedly demonstrated in laboratory cultures under optimal conditions Citation[31–33], but in environmental samples from different locations Citation[30]. In a comparative study, diatoms grew faster under low light conditions than competing algae Citation[34]. Higher photosynthetic efficiency for diatoms not only relates to other Stramenopiles, but also in comparison to green algae (molecular mechanisms will be discussed below). A comparison between P. tricornutum and Chlorella vulgaris indicated that under nonfluctuating light conditions, efficiencies of converting light into biomass were equivalent, but under fluctuating light conditions, the diatom was nearly twice as efficient as C. vulgarisCitation[35]. Efficient carbon fixation ability along with nutrient utilization could increase the appeal of diatoms for co-processes such as CO2 abatement and waste water remediation.
▪ The relevance of natural productivity to biofuels production
Several of the concepts presented in the previous paragraphs are of relevance to biofuels production. Diatoms naturally bloom under nutrient-replete conditions, meaning maximal growth rate to highest density is obtained, which are ideal characteristics for a biofuel production system. Diatom blooms also tend to crash precipitously, which is a tendency of all bloom species. Crashes can result from a number of factors including nutrient exhaustion, grazing, sedimentation, or viral lysis and remains a fundamentally poorly understood process in all microalgae Citation[36,37]. Environmental studies have shown that once chlorophyll levels begin to decrease (signaling the beginning of the end of the bloom), triacylglycerol (TAG) content increases Citation[38,39]. Translating that into a biofuels production scenario, TAG accumulation precedes cell death and thus crashing would not necessarily be detrimental to production. Diatoms also grow best under highly mixed conditions, which are desirable for large-scale cultivation. Their large contribution to carbon export in the oceans relates to their relatively rapid sinking rates, which could translate into enhanced harvesting ability in a production system (see below). Their ability to sequester nutrients could be advantageous for crop management; supplying a nutrient pulse in which most of a particular nutrient is rapidly taken up and removed from the culture medium could reduce nutrient availability for contaminating organisms. Vacuole size and nutrient storage capacity should affect different aspects of production schemes. Chlorophytes, including Chlorella species, Chlamydomonas reinhardtii and Dunaliella salina, have small vacuoles (3–14% of their protoplast volume) compared with diatoms (45–90% of their protoplast volume), and should have a greater dependence on external nutrient conditions including consistency of supply Citation[19]. In terms of lipid productivity, cells with larger vacuoles may take longer than cells with smaller ones to become nutrient limited, due to the enhanced nutrient storage capability.
▪ Diatom productivity in large-scale culture systems
Diatoms have been cultivated in large-scale outdoor systems for purposes other than biofuel production, such as aquaculture, for decades Citation[40]. Widely ranging values for algal biomass productivity are reported in the literature and are due in large part to differences in cultivation parameters. Such parameters include, but are not limited to, light source and intensity, depth of culture, mixing rate, temperature, nutrients and supplementary carbon additions in the form of sugars or CO2. In an attempt to limit some of the variability for the purpose of comparison, only values obtained from microalgae grown in outdoor ponds, raceways or open photobioreactors illuminated with natural light are included here. Diatom biomass productivity values range from 5 to 25 g m-2 day-1Citation[30,41–43], and are in line with values obtained for other types of microalgae including the Eustigmatophyte Nannochloropsis salinaCitation[44], the cyanobacterium SpirulinaplatensisCitation[45] and the Chlorophyte Chlorella sp. Citation[46], whose biomass productivity values range from 10–25 g m-2 day-1. Cyclotella cryptica, a model diatom species from the Aquatic Species Program, was shown to have consistent productivity levels of 20 g m-2 day-1 (erroneously listed as 12 g m-2 day-1 in the text of this paper Citation[41]).
What cellular factors contribute to diatom productivity?
As discussed previously, the evolution of diatoms is more complex and unique than that of green algae and terrestrial plants. As a result, there are fundamental differences in the biology (i.e. metabolism, physiology and subcellular organization) of the different algal classes Citation[11]. Acknowledging diversity within a class, it is still reasonable to expect there will be advantages or disadvantages to certain classes in a large-scale biomass production setting. It is useful to outline some of the variation between classes so as to better understand the biology of the organisms that we propose to cultivate for fuel. Since chlorophytes are commonly considered as algal biofuels production strains and are fundamentally different than diatoms, we will focus on comparing these two classes. We acknowledge that there are other classes of photosynthetic organisms (Nannochloropsis, cyanobacteria) being considered for large-scale cultivation that are not included in this discussion.
▪ Differences in photosynthesis & carbon fixation comparing diatoms & green algae
Dissipation of excess light in photosynthesis either occurs at the level of photosystem II (PSII) and especially light-harvesting complex II (LHCII) via thermal dissipation or adjustment of the absorption cross section; or by redirecting electrons into pathways other than CO2 fixation. Diatoms possess an additional xanthophyll-based cycle involved in energy dissipation relative to green algae and higher plants Citation[47]. In a comparative study, the diatom P. tricornutum favored modulation at PSII/LHCII, with a substantial increase in de-epoxidation of diadinoxanthin to diatoxanthin, which reduced the flow of electrons into secondary processes. This was more efficient than C. vulgaris, where thermal dissipation at LHCII was less Citation[48]. Green algae undergo state transitions where reorganization of antenna proteins shifts primary activity between PSII and PSI, especially under fluctuating light conditions Citation[49]. State transitions increase photosynthetic efficiency, as has been demonstrated by mutants in this process, which have reduced capacity to make ATP via cyclic-photophosphorylation and have impaired growth Citation[49]. No evidence for state transitions or a reaction center-based quenching mechanism has been found in diatoms Citation[50], yet they are inherently more efficient than chlorophytes in balancing photosynthetic electron flow Citation[48]. Perhaps the lower efficiency of chlorophytes involves the need for physical movements of antenna proteins over 500 nm Citation[49], as compared with localized quenching by additional xanthophyll as occurs in diatoms. Diatom thylakoid membranes are arranged in groups of three and are not differentiated into grana and stroma lamellae Citation[51,52], which affects redox signaling. Diatoms lack the α-carotene biosynthetic pathway, which means that both photoprotective and light harvesting pigments are synthesized from the same precursors, in contrast to the chlorophytes Citation[48]. In addition, in contrast to the chlorophytes, diatom light-harvesting proteins are not differentiated into major and minor complexes Citation[53,54], the Calvin–Benson cycle is not controlled by light via the redox state of thiol groups Citation[55] and the oxidative pentose phosphate cycle is apparently not present in the chloroplast Citation[55]. It is reasonable to assume that these substantial differences in photosynthesis, and carbon fixation and flux, contribute to diatoms’ environmental success.
▪ Carbon fixation efficiency & concentrating mechanisms
Both diatoms and chlorophytes possess carbon concentrating mechanisms (CCMs), which effectively increase CO2 concentration in the proximity of RuBisCO. It is not clear whether there are advantages in CCM efficiency intrinsic to either class; however, there are significant differences in the ability of RuBisCO to discriminate between CO2 and O2. The chlorophytes have Form1B RuBisCO, with selectivity (Srel) values ranging from 54 to 83, whereas diatoms have Form1D RuBisCO, with a much higher Srel of 106–114 Citation[56]. CCMs can involve two different mechanisms: biochemical (e.g., C4 metabolism), in which carbon is fixed by an enzyme other than RuBisCO and the fixation product or a derivative is decarboxylated in the vicinity of RuBisCO; or biophysical, which involves localized enhancement of CO2 via equilibrium shifts involving pH changes or active transport of inorganic carbon across membranes Citation[57]. The latter involves carbonic anhydrase (CA) and bicarbonate transporters. In diatoms, evidence for both C3 and C4 biochemical fixation exists, and the preference can apparently differ in relatively closely related species Citation[58], which has led to confusion in attempts to make generalizations. P. tricornutum and T. pseudonana encode 9 and 13 CAs, respectively, but only P. tricornutum has β-CAs that are targeted to the chloroplast Citation[59]. It is as yet unclear whether differences in CCMs relate to the different evolutionary time periods under which different classes of diatoms arose in relation to CO2 levels. The documented differences may either reflect an organization that maximizes efficiency under a given CO2 condition or, alternatively, that a variety of organizational schemes provide acceptable efficiency under current conditions. More research, especially localization studies, is required to clarify the situation. Variation in CCM mechanisms may not be unexpected; according to a modeling study, the advantage of a CCM would be best suited for an intermediate-sized algal cell, and there should be a high degree of co-adaptation in different uptake systems Citation[60].
▪ Compartmentation & carbon metabolism in diatoms
Subcellular compartmentation enables incompatible metabolic processes to occur by physically separating them, and many biochemical processes (lipid metabolism, oxidative phosphorylation, photosynthesis) take place on the membranes of subcellular compartments, which also provide a greater area for these processes to occur Citation[61]. Stramenopiles have additional intracellular compartmentation relative to chlorophytes as a result of their secondary endosymbiotic origin. In particular, the chloroplast is surrounded by two additional membranes, the chloroplast endoplasmic reticulum and the periplastidic membrane Citation[62,63]. These two membranes are closely associated, but there is a noticeable space between the periplastidic membrane and the outer chloroplast membrane Citation[64]. This periplastid compartment appears to be functionally significant, as several enzymes involved in myriad of cellular processes, including lipid transport and metabolism, have been shown to be targeted there Citation[65]. Though the specific roles of this compartment are far from resolved, it is clear that it plays a part in regulating metabolism and overall cellular function.
Similar to green algae and terrestrial plants, diatom genomes contain genes for the essential enzymes of the conserved eukaryotic central carbon metabolic pathways of carbon fixation, glycolysis/gluconeogenesis and the citric acid cycle Citation[8,10]. One difference in the glycolysis pathway is that neither T. pseudonana nor P. tricornutum encode a gene for a glucokinase Citation[66]. Perhaps more surprising is the localization of the complete lower half of the glycolysis pathway from triose phosphate isomerase to pyruvate kinase to the mitochondria Citation[66]. Diatoms apparently have the unique ability to convert glyceraldehyde-3-phosphate into pyruvate, along with the attendant intermediates, entirely in the mitochondria. The role of this partial pathway in the mitochondrion is unclear, though it could be a strategy to channel substrates towards the citric acid cycle. In terrestrial plants, an increased association between cytosolic glycolytic enzymes and the mitochondrial surface occurs during higher respiration, with attendant-increased efficiency through substrate channeling Citation[67]. A mitochondrial-localized glycolysis pathway could also avoid the need for the malate-aspartate shuttle to import the reducing equivalents needed to generate ATP through oxidative phosphorylation.
Diatoms contain a complete urea cycle Citation[10], which is not found in chlorophytes, and several metazoan-like urea cycle components are localized to the diatom mitochondria. In metazoans, the ornithine-urea cycle is only known for its essential role in the removal of fixed N; in diatoms, however, it apparently serves as a distribution and repackaging hub for inorganic carbon and N Citation[68]. Experimental manipulation of the cycle indicates that it contributes significantly to the metabolic response of diatoms to episodic N availability Citation[68].
Diatoms also differ from other classes of algae in terms of carbohydrate storage. Instead of starch, diatoms store fixed carbon as a complex carbohydrate called chrysolaminarin, which contains β(1→3) and β(1→6) linked glucose units in a ratio of 11:1 Citation[69]. Chrysolaminarin is soluble and is stored in the chrysolaminarin vacuole (CV). The CV is quite large Citation[70], and recent imaging work in our laboratory in collaboration with Robyn Roth at Washington University (MO, USA) indicates that it can comprise as much as 50% of the cell volume. Diatoms are not necessarily better at storing carbohydrates than green algae. For example, C. reinhardtii has quite an impressive ability to accumulate starch Citation[71]. However, it may be important to consider the relative energetics involved in accessing a soluble carbohydrate (chrysolaminarin) stored in a vacuole versus an insoluble form (starch) stored in the chloroplast.
▪ The silica cell wall
The most conspicuous feature of diatoms is their silica-based cell wall, also called the frustule . The frustule consists of two halves that overlap like a petri dish, the epi- and hypo-theca. The ‘top’ and ‘bottom’ of the dish are made of structures called valves, and the sides are most typically composed of a series of thin siliceous bands that encircle the cell, overlap each other, and provide the overlap between the theca, called the girdle bands. During vegetative division, diatoms make new valves in an intracellular compartment called the silica deposition vesicle (SDV), and once complete, the entire structure is exocytosed Citation[72]. After the two daughter cells separate, individual cells can sequentially synthesize new girdle bands, which enable elongation of the cell. The source of Si for diatoms is dissolved silicic acid, Si(OH)4Citation[73], which is taken up at low concentrations by specific silicic acid transport proteins, and at high concentration by diffusion across the plasma membrane Citation[74].
The silica-based cell wall not only serves a protective function, but can contribute to diatoms’ overall productivity. Silica is polymerized from soluble silicic acid precursors in an energetically favorable autocatalytic process when silicic acid concentrations are sufficiently high (above 2 mM) and/or the pH is lowered Citation[75]. In diatoms there are organic components that speed up this process; however, they are present in, at most, a few percent relative to the silica that is formed. The energetic needs for silica polymerization are thus much lower than an organically based cell wall, and calculations done for silicified plant cell walls indicated that silicification required only 3.7% of the energy compared with a lignin-based wall, and 6.7% for a polysaccharide wall Citation[76]. In addition to the energetic savings, there should be savings in carbon, since a large percentage of the carbonaceous cell wall components are replaced with silica and the carbon can be used for other cellular purposes. Another feature of the cell wall that may relate to biofuels and specifically TAG accumulation, is that very different cell shapes can result, which may impose constraints on the size and number of lipid droplets that can be accommodated in the cell . There may actually be species-specific geometrical features that relate to optimal productivity.
The silica cell wall also apparently plays an important role in carbon concentrating mechanisms. Data suggest that diatom biosilica is an effective pH buffer, which increases carbonic anhydrase activity near the cell surface to enable the conversion of bicarbonate to CO2Citation[77].
Attributes of diatoms for biofuel production
In addition to efficient photosynthesis and nutrient utilization, diatoms possess several other attributes that relate directly or indirectly to useful characteristics for a biofuels production system.
▪ Environmental & trophic flexibility of diatoms
It is envisioned that algal biofuels development will have a regional character, and there will be a need to develop strains optimally adapted to different environments Citation[1]. Further, the development of different types of culture strategies, ranging from outdoor open systems to enclosed photobioreactors to hybrid systems, and different trophic strategies, from auto- to hetero- to mixo-trophic, must be considered Citation[78]. Thus algae with diverse culturing characteristics are desirable.
On this note, diatoms are found in a wide variety of environments, including general environments such as marine, freshwater and soil, but also specific extreme or variable environments of salinity, temperature (ice or hot springs) or pH. The most intensively studied diatoms are marine, and all of the species with sequenced genomes fall into that category. This does not preclude freshwater diatoms from being developed for biofuels production, but considering the issue of water supply, saline-tolerant species may be the most appropriate for development. Diatoms isolated from thalassic hypersaline marine environments include both centric and pennate species that grow well at salinities ranging from 0.5 to 15%, and three strains of Pleurosigma strigosum cannot grow in salinities less than 5% total salts, representing truly halophilic diatoms Citation[79]. Different strains of P. tricornutum were shown to have near-linear growth rates (divisions per day) in salinities ranging from 5 to 70 psu Citation[80]. Numerous species grow under acidic conditions (pH 3.3–4.4 Citation[81]) and diatoms have been found to grow at pHs consistently maintained between 1 and 2.5 Citation[82]. Conversely, a wide array of both pennate and centric species have been found in alkaline lakes, such as those in East Africa, where the pH can reach 10.6 Citation[83]. Diatoms can grow at 60–65°C in hot springs Citation[84] and at 40°C in thermal muds, which are a 5 g l-1 colloidal aluminum silicate suspension mixed with thermomineral water, rich in sodium chloride and sulfates Citation[85]. Psychrophilic diatoms grow on polar ice and in hypersaline brine channels at -1–2°C Citation[86], with photosynthetic activities optimized between 4 and 7°C Citation[87]. Diatoms can form biomasses in ice with chlorophyll concentrations that are matched only in hypertrophic waters or in extremely eutrophicated sediments Citation[88]. The Fragilariopsis genus may be the most prevalent biomass producer among polar plankton Citation[89,90], serving as the base of the entire polar food web. The range of different environments where diatoms can dominate sheds light on their diversity; in terms of biofuels production, growth in extreme conditions, such as salinity or pH can be utilized to keep pond contaminants and predators at low levels.
Diatoms exhibit trophic flexibility. The majority of species are autotrophic, but mixotrophy and obligate heterotrophy occur. C. cryptica, a candidate biofuels production strain, is capable of slow growth (10% of autotrophic) on glucose only in the dark, likely due to its glucose transport mechanism being induced in the absence of light Citation[91]. The mixotrophs and obligate heterotrophs are generally able to use simple sugars as carbon sources, but some species are able to break down complex carbohydrates. There are two forms of mixotrophs: those that can grow strictly auto- and hetero-trophically, or those whose growth is only enhanced in the presence of an extracellular carbon source Citation[92]. Several Nitzschia species can switch between strict auto- and hetero-trophic growth Citation[93] and have been observed to burrow into solid medium containing agar, presumably breaking it down as a carbon source. In the natural environment, heterotrophic diatoms can be found on macroalgae (kelp), from which they obtain their carbon Citation[94]. Nitzschia alba is an obligate heterotroph that has lost its chloroplasts, and can grow on both simple and complex carbohydrates Citation[94–96], demonstrates growth rates in the field appreciably higher than other heterotrophic protists, and doubles in 6 h when cultured in the presence of glucose Citation[95]. The ability of diatoms to breakdown complex carbohydrates suggests possible applications with different carbon feedstocks, such as cellulose.
▪ Lipid amount & composition
A primary target for biofuels production is the accumulation of lipid, either in the form of total lipid or TAG, which provide carbon for conversion to a variety of types of fuel Citation[1]. Diatoms are excellent lipid accumulators, which partly may be due to extra buoyancy needed to counteract the effect of their relatively heavy silica cell walls. It is vitally important when comparing lipid content numbers to appreciate the variability in conditions in different studies; strict comparisons cannot generally be made. However, by comparing data from a sufficiently large number of studies some general conclusions can be formed.
In terms of total lipids, Shifrin and Chisholm Citation[97] reported values of 12–45% of dry cell weight (DCW) for diatoms under conditions of N limitation. A comparative analysis of several species in different algal classes by Hu et al.Citation[98] showed that on a DCW basis, chlorophytes had an average total lipid content of 25.5% under normal growth and 45.7% under nutrient stress. Regarding diatoms, lipid content was 22.7% DCW during normal growth and 44.6% during stress as indicated in the text, but this value was mistakenly listed as 37.8% in their Figure 1, as the line depicting the average is clearly above 40% Citation[98]. A literature survey by Griffiths and Harrison Citation[99] of 55 microalgal species in various classes showed that diatoms as a class performed above average at 27.4% (19% better than average) for total lipid content under nutrient-replete conditions and 36.1% (13% better than average) under N-deficient conditions. Diatoms also accumulate lipid as a result of Si limitation, and the average lipid content for diatoms limited for Si relative to N deficient conditions was 41%, which was 28% higher compared with all species and 13% higher when compared within the diatoms Citation[99].
In terms of neutral lipid (TAG), Si-deficient cultures of C. cryptica were reported to contain 64% on an ash-free dry weight (AFDW) basis, compared with 32% in Si-replete cultures, and Cylindrotheca fusiformis contained 57% in Si-deficient cultures and 17–20% in Si-replete cultures Citation[201]. A wide variety of TAG composition has been identified in diatoms, with fatty acid (FA) side-chains ranging from 14:0/16:0/16:1 to 16:1/20:5/22:6; however, there were ten predominant species, which were enriched in C14 and C16 Citation[100].
Diatom FAs are among the most highly enriched in C14 chain lengths compared with other classes of algae Citation[98], especially chlorophytes, which appear to lack this particular length Citation[101–103]. Reports range from 4 to 32% C14:0 of the total FA composition for diatoms Citation[98,103,104]. C16 FAs are highly represented in diatoms and C16:0 and C16:1 combined can represent 65–70% of the total FA, with C16:1 being predominant. C18 lengths are poorly represented in diatoms, but can be a predominant form in chlorophytes, especially polyunsaturated forms Citation[98,102]. In general, shorter chain lengths are desirable to improve cold-flow properties of biofuels Citation[105] and saturated FAs are more desirable because they increase the ignition quality (cetane number) of the fuel Citation[106].
A critical point to make regarding measurement of total lipid or TAG in diatoms is that DCW measurements will underestimate the actual amount of lipid in terms of percent cellular carbon when compared with a similar measurement in other classes of algae because of the contribution of the noncarbonaceous silica cell wall. Comparison of eight different diatom species indicated ash contents from 49 to 59% Citation[107]; further, diatoms had an average of twice the ash content of a variety of nonsilicified algae and four times the content of two Chlamydomonas species examined Citation[97]. The silica content of three diatom species was measured at 22–47% of the dry weight Citation[108]. Factoring this into DCW measurements, this suggests that lipid content in diatoms on a per cell carbon basis is substantially higher than usually reported. Illustrating this point is a comparison of Si-induced lipid accumulation in C. cryptica, in which total lipid based on DCW was 42% Citation[97] compared with 54.1% when calculated on an AFDW basis Citation[109]. The data suggest that diatoms are at least on par with, if not superior to, other classes of algae in terms of their ability to accumulate lipids.
▪ Other valuable hydrocarbons, EPA, DHA & isoprenoids
Diatoms are unusually highly enriched for eicosapentaenoic acid (EPA; C20:5, ω-3), which is not prevalent in chlorophytes Citation[98,103,104]. Fish do not naturally produce EPA, but obtain it from algae they ingest, to which diatoms should contribute significantly. Docosahexaenoic acid (DHA) levels in diatoms are less than EPA, on the order of 5% of total lipid Citation[98,103,104]. Diatoms contain a rich variety of carotenoids in the form of β-carotene, as well as a number of oxygenated derivatives called xanthophylls. These include fucoxanthin, neofucoxanthin, diadinoxanthin and diatoxanthin, some of which play important roles as accessory pigments and photoprotective agents in the light-harvesting complex of the photosystems Citation[110]. Reports of diatom carotenoid content ranges between 0.07 and 0.6% of the dry weight (0.7–6.3 mg pigment/g dry wt), with pennates exhibiting a higher content than the centrics that were examined Citation[108]. Some diatom xanthophylls are used commercially for their color and antioxidant properties Citation[111], and it should be possible through recombinant techniques to dramatically increase production of specific desired xanthophylls in microalgae that naturally produce these pigments. Diatoms possess both the 2-C-methyl-D-erythritol 4-phosphate (MEP) or nonmevalonate pathway for carotenoid synthesis, and the mevalonate pathway (MEV) for synthesis of sterols, sesquiterpenes and triterpenoids Citation[112,113].
▪ Antipathogenic or medical applications of extracted diatom components
Although not a major field of research, added-value chemical components of diatoms have been described. These include antibacterial and antifungal compounds Citation[114–116], which are present in FA extracts Citation[117,118]. Antibacterial activity extracted from Skeletonema costatum was shown to inhibit the growth of Vibrio, a pathogen of fish and shellfish Citation[118]. Other extracts of S. costatum and Haslea ostrearia demonstrated anti-tumoral activities against human lung cancer and also had anti-HIV effects Citation[119–121]. Highly branched C25 isoprenoidpolyenes (polyunsaturated sesterpenes oils orhaslenes) were identified as being responsible for these activities Citation[122]. A recent report documented that marine benthic diatoms contain compounds able to induce leukemia cell death and modulate blood platelet activity Citation[123] suggesting that diatoms may represent a richer source of interesting bioactive compounds than previously recognized.
▪ The advantages of Si limitation-based lipid accumulation induction
As far as has been documented, diatoms accumulate TAG only under limitation conditions. Published literature has shown that TAG accumulates under either Si or N limitation Citation[98,109], and in we present a comparison of these conditions with a treatment where the cells were exposed to the microtubule-based cell cycle inhibitor nocodazole. There are several important points to consider from this. First is that only cell cycle arrest, and not nutrient limitation, is required for induction of TAG accumulation in diatoms . This has been substantiated by other experiments in our laboratory in which cell cycle stage is directly measured, and the results demonstrate that TAG does not begin to accumulate until arrest has occurred [Smith SR et al. Unpublished Data]. Often, it is stated in the literature that TAG accumulation occurs only under nutrient stress conditions; however, experiments in our laboratory indicate that, for diatoms stopping the cell cycle, even under nutrient-replete conditions is sufficient . A second point is that TAG accumulation under Si limitation lags slightly behind nocodazole treatment, but reaches and eventually exceeds the maximum level . The difference in induction start points may be related to a longer time required to halt the cell cycle under Si limitation. The third point is that TAG accumulation under N limitation lags substantially in timing and level of induction compared with nocodazole and Si. After a very long limitation period, abundant lipid droplets will form under N limitation, but only several days after similar sized droplets are seen under Si limitation [Traller JC, Unpublished Data]. The reason for the differential response comparing Si with N limitation is likely due to the differences in extent of intracellular storage of the two elements. N is stored intracellularly in a variety of forms and can be accessed over time, resulting in continued growth even after cells are placed in a N-free medium. Si is not appreciably stored in diatom cells, and hence placement of cells into a Si-free medium results in relatively rapid arrest and, therefore, a faster onset of TAG accumulation.
In terms of a production scenario, a lag time for induction of TAG accumulation would be unfavorable – increased time means decreased overall productivity. Thus, N limitation would not be desirable for diatoms, especially when Si limitation is an option. For this reason, it is a valid question whether diatoms without a Si requirement, such as P. tricornutum, would be competitive in a production system. The rate of TAG accumulation in chlorophytes resulting from N limitation varies among species. In C. reinhardtii, experiments on wild-type, starchless and complemented mutants showed that TAG levels increased on the order of twofold within 24 hours Citation[124,125], suggesting a more rapid response under N limitation than with diatoms. Consistent with this, growth arrest in C. reinhardtii under N limitation appears to be fairly immediate Citation[126]. In C. vulgaris, growth arrest was not immediate, and increased cell number was evident 2–4 days after placement in a N-free medium Citation[126]. To our knowledge, no correlation between TAG accumulation and cell cycle arrest has been reported in chlorophytes, although correlation has been made between life cycle stage and TAG accumulation Citation[101].
▪ The detrimental effects of N limitation
In addition to the time consideration, there are other drawbacks of having to rely on N limitation to induce TAG accumulation. There are numerous detrimental effects of N limitation on photosynthesis Citation[127,128], which include decreased chlorophyll, alteration of thylakoid stacking, reduction in PSII cross-section and decreased energy transfer from the LHC to reaction centers. N limitation reduced photosynthetic rates and acetate uptake in ChlamydomonasCitation[125] and LHC protein content decreased 10–40-fold, contributing to chlorosis Citation[129]. Chlorophyll levels typically decrease under N limitation, which does not occur under Si limitation in diatoms Citation[127,130]. N limitation also dramatically reduces ribosome content in C. reinhardtiiCitation[131]. N-deprived microalgae typically have reduced protein content; for P. tricornutum, N starvation led to a decrease in abundance of RuBisCO, especially the small subunit Citation[132]. De novo protein synthesis is necessary for an increase in acetyl CoA carboxylase activity, the first committed step in FA biosynthesis Citation[100,133]. General protease activity was increased in a diatom and Dunaliella tertiolecta under N limitation Citation[99,134]. In terms of transgenic approaches to improve productivity, because of the overall decrease in protein synthesis under N limitation Citation[127], any expressed proteins are likely to be less effective than desired. The combined effects suggest that N limitation is truly stressful to the cells, which raises the question of how cell ‘health’ is related to its ability to synthesize and accumulate TAG. In diatoms, Si limitation has little direct effect on other aspects of cellular metabolism than the cell cycle Citation[135]; for example, protein and RNA synthesis are not appreciably affected Citation[136].
▪ The ease of extraction of lipid from diatoms
On occasion we have heard comments that it must be more difficult to extract lipid from diatoms because of their solid cell walls. This is not true. Although cell wall components are solid, the overall structure is composed of discrete siliceous substructures that are joined together by organic molecules. The individual parts are relatively easily disrupted, and organic solvents readily extract lipid and pigments Citation[137,138]. The diatom cell wall is actually easier to break open than the wall of some green algae; for example, RNA isolation procedures for diatoms involve simply vortexing cells in the presence of Tri Reagent Citation[139], whereas for some green algae frozen cells have to be ground in a mortar and pestle prior to extraction. Furthermore, pigments are easily extracted from diatoms and blue-green algae using 90% acetone, whereas harsher solvents are required for the extraction of pigments from coccoid green algae Citation[140]. In a biofuels production setting, where the energetic cost of extracting lipid is high, diatoms and other more easily extractible algae may provide a significant economic benefit.
▪ Settling rates, flocculation, harvesting & shear stress
One of the most expensive aspects of microalgal biofuels production is harvesting and dewatering; the need to concentrate a small percentage of solids from a much larger volume of water Citation[141]. Factors that would reduce these costs include increasing settling rates or the ability of microalgal cells to flocculate. As stated previously, diatoms are major exporters of organic carbon from the ocean surface due to their increased settling rates relative to other phytoplankton Citation[142]. The efficiency of this settling rate is likely to be at least partly dependent on the shape of the diatom. One report documented that elongated pennate diatoms were problematic in a water treatment facility because they did not settle well in the facility’s sedimentation tanks, causing clogs in the downstream sand filters Citation[143]. However, diatom cell walls are covered with organic material having properties similar to other algal cell walls, and respond well to both cationic and anionic chemical flocculation treatments Citation[144]. Further, diatoms are known to flocculate naturally and chain-forming marine diatoms can aggregate into centimeter-sized flocs in as little as 24 h Citation[145]. The ability of diatoms to naturally flocculate is species-specific Citation[146], facilitated by a variety of mechanisms ranging from direct adhesion to the cell surface production of exopolymers Citation[147]. Some species may even produce anti-flocculation compounds Citation[147]. Increased exopolymer secretion in conjunction with cellular protuberances that trap other particles is a common sinking mechanism in marine species, which is often triggered near the end of a bloom and generates sinking rates of 100 m day-1 or more Citation[148]. These data indicate that understanding diatom self-flocculation strategies may be of value in the context of biofuels production.
Diatoms are well adapted to turbulence, and in fact are more productive in high turbulence conditions than other classes of marine microalgae Citation[149]. However, there is some apparent contradiction in the literature concerning this point, in that some studies indicate that diatoms are especially sensitive to shear during cultivation Citation[150,151]. In one report, the authors concluded that micro-eddies on the scale of the cell size were responsible for shear stress Citation[150]. A suggestion was made that diatoms may be especially sensitive to shear stress because of their rigid silica walls Citation[152]; however, the particular species used in this study was predominantly unsilicified. Another study examined the passage of two diatom species through pumps and valves and concluded that the extent of cellular damage was related to operational parameters, such as the pressure drop and the rotational speed of the pump Citation[153]. Cultivation in our laboratory has revealed that some species are impervious to highly turbulent mixing, whereas others are sensitive to modest agitation, suggesting that species-specific differences need to be taken into account with regard to shear stress; such differences have also been reported in the literature Citation[150,154]. The data suggests that the issue is not with diatoms as a class, but particular diatom species subjected to particular turbulence conditions.
▪ Susceptibility to predators or pathogens & resistance to contamination
In spite of the protective cell wall, diatoms are not immune to predators and pathogens. A major class of diatom predator in the ocean is copepods, which are small crustaceans Citation[155]. Interestingly, diatoms have apparently developed both physical and chemical defenses against copepods. A study demonstrated that diatom cell wall architectures were optimized to resist crushing Citation[156], in this context by copepod jaws, and diatoms also produce aldehyde derivatives that have been implicated in reducing copepod reproductive viability Citation[155]. Apofucoxanthinoid compounds (derived from fucoxanthin) produced by diatoms act as copepod feeding deterrents at concentrations of 2.2–20.2 ppm Citation[157]. Small flagellates also feed selectively on diatoms Citation[158]. In general, diatoms represent a bulky food for ingestors and their cell walls must be compromised during ingestion or else the intracellular contents are not completely accessible. Chytrid fungi infect diverse prokaryotic and eukaryotic algae, including diatoms Citation[159]. Phytomyxea (plasmodiophorids) parasites have also been shown to parasitize diatoms Citation[160]. Diatom viruses were first described in 2004 Citation[161] and are either ssRNA or ssDNA viruses that infect the genera Rhizosolenia or ChaetocerosCitation[162]. It is likely that the measures to deal with predation and pathogens that are being considered for other classes of algae will need to be applied to diatoms as well for biofuels production. In terms of cross-contamination of cultivation systems or environmental contaminants, one advantage of the defined shapes and sizes of diatoms is that it is generally easy to use microscopy to identify differences between diatom species, as well as to distinguish diatoms from other microalgae. The generally higher nutrient uptake rate of diatoms compared with other classes of algae (as mentioned previously) could also contribute to reduced contamination.
▪ Genomic & transgenic aspects of diatoms
Genetic modification is likely to play an important role in the development of algal biofuels, both as a research tool and in possible field applications once appropriate regulations are in place. Diatoms are well established in terms of genomic and transgenic capabilities. There are currently four available diatom genome sequences, for T. pseudonana, P. tricornutum, Fragilariopsis cylindrus and Pseudo-nitzschia multiseries. Of the four, F. cylindrus and Pseudo-nitzschia multiseries may not have biofuels relevance, because neither grows at a fast enough rate for a production system. Sequenced genome sizes in diatoms vary from compact at 27 Mbp to larger on the order of 300 Mbp, and GC content is close to 50% Citation[8,10]. The latter point is important because many genes already developed for mammalian or yeast purposes (e.g., eGFP) can be used directly in diatoms with no modification, such as codon optimization Citation[163]. Another important point is that there is no evidence for gene silencing in diatoms, as occurs in some chlorophytes Citation[164]. In one study, a transgenic diatom was cultured in the absence of antibiotic selection for 1 year and retained resistance Citation[165].
Currently diatom transformation is limited to the biolistic approach Citation[165,166], which is effective, but inefficient. Higher efficiency methods would be desirable; for example, to enable saturation screening of insertional mutants or to complement mutations by introducing a library of genes. Integration of introduced DNA occurs randomly in diatom genomes Citation[162], which can result in different levels of expression depending on the site of integration. This is not necessarily negative; on the one hand, more involved screening is required to identify useful transformants, but the variation in expression can be useful in obtaining a graded response. Homologous recombination has not been reported in diatoms, so gene replacement is not yet an option. Promoters that drive overexpression of genes have been identified Citation[163,165,166], as well as a regulated promoter that can be turned on or off by changing the N source in the growth medium Citation[167]. Gene knockdowns have also been accomplished using RNAi approaches Citation[168]. Although there is room for improvement, a complete set of basic genetic manipulation tools are available for diatoms.
▪ Diatom metabolic diversity could provide abundant targets for bio-engineering
The evolutionary divergence of centric and pennate diatoms occurred at least 90 million years ago (based on the fossil record), yet their genomes differ from one another to the same extent as those of fish and mammals, which diverged approximately 550 million years ago Citation[8]. The relatively rapid rate of diatom evolution has been at least partially attributed to the acquisition of horizontally acquired genes, which are thought to confer novel cellular and metabolic functions, and are drivers of diversification Citation[8]. However, there are also substantial differences between diatoms in the targeting and homology of core genes involved in highly conserved eukaryotic pathways and processes. These include differences in the homology and distribution in enzymes of carbohydrate metabolism, the pentose phosphate pathway and carbonic anhydrases Citation[59,68,169–171]. Differences in these core pathways are likely to affect metabolism and/or cellular energetics. These differences indicate that the trophic and environmental flexibility found within the diatoms has a genetic basis. Diversity in the core metabolism of diatoms means that there are multiple possibilities for manipulation of enzymes and pathways to improve growth and lipid accumulation characteristics.
Perceived disadvantages of diatoms in large-scale cultivation
The vast majority of diatoms require Si for growth, and thus a major difference between cultivating diatoms and other classes of algae is the requirement to add soluble Si to the medium to support growth. The presence of an additional nutrient can be perceived as a drawback in a culture system because of increased cost and, as in the case of Si, ancillary problems that could arise because of its aqueous chemistry. Complete details of the aqueous chemistry of Si can be found in Iler Citation[75]. As a brief summary, soluble Si is present in aqueous solution as silicic acid, Si(OH)4, which is the predominant form at pH 8.0, with a small fraction (3%) of the monoanion Si(OH)3O-. The solubility of silicic acid is limited at neutral pH to approximately 2 mM, above that concentration it begins to autopolymerize to first form multimers, which eventually coalesce into solid silica, SiO2, which can precipitate from solution Citation[75]. Silicic acid can form complexes with metal ions, and magnesium silicates are especially prone to precipitation Citation[75]. Another perceived problem with silica relates to the important distinction between the amorphous silica found in diatoms and crystalline silica found in asbestos. Although chemically identical, their toxic effects are completely different. Needle-shaped crystalline silica is a causative agent in silicosis, in which scarring occurs in the upper lobes of the lungs. Amorphous silica is harmless to all organisms other than insects (see below) when breathed, ingested or contacted; thus, large-scale diatom cultivation will not produce a hazardous silica product.
There are advantages to the Si requirement for diatoms in cultivation. As discussed previously , Si limitation results in faster TAG accumulation than N limitation, with attendant reduction in production time. An interesting concept is whether a Si limitation lipid strategy actually requires more N than a species that accumulates under N limitation. There are two considerations regarding this: increased neutral lipid productivity under Si limitation may outweigh the extra cost of N, and N could be included during the Si-limitation phase at levels much lower than used for growth to prevent the detrimental effects of N limitation as discussed previously, but not in excess of that, thus minimizing cost. Another benefit of the diatom’s Si requirement is that biomass accumulation can be precisely regulated by the amount of Si added to batch cultures. Many diatom species have well-defined silica content in their cell walls; thus, there is a strict dependency on how many cell divisions can occur with a given starting amount of Si in the medium. This is illustrated in , where batch cultures of C. cryptica were grown with different starting concentrations of Si. Growth arrested in each culture on day 7, and the resulting culture density was precisely in the ratio of the starting amount of Si in the media. Such control could be highly valuable in a large-scale culture system because biomass attainment would both be predictable and controllable. Similar control may not be likely with other nutrients, such as N, given the issue of intracellular storage and the abundant presence of N sources in the environment (including N fixers). There are scenarios where Si limitation may be problematic in a production setting; for example, Si-containing dust could blow into an open system and perturb the control. However, the effects of this type of contamination should also be predictable once the rate of dissolution of airborne Si into the medium is determined. It should be noted that it is not likely that all diatom species will exhibit such precise control of biomass relative to Si concentration. Heavily silicified diatoms are known to reduce in size with succeeding generations (eventually triggering the sexual cycle), which leads to a mixed size population of cells with varying amounts of silica in their walls. An important point is that size reduction is not universal in diatoms, and in most laboratory species there is no evidence of size reduction, and have documented reasons why it does not have to occur Citation[172].
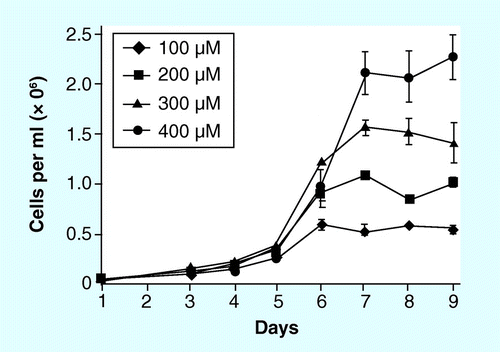
Given its limited solubility (0.07% in water), the method of delivery of Si to a culture system deserves some consideration. Most production systems are maintained at or above pH 8, which is favorable for silicic acid solubility; however, there are still limits. A maximum of 2 mM Si could be added to a batch culture, but in addition to pushing the solubility limit, such high concentrations could affect the growth of diatoms. Environmental concentrations of silicic acid are rarely above 100 µM, and we have observed detrimental effects on some species in laboratory cultures at 400 µM. Fortunately, the amount of free silicic acid in a culture can be readily measured, either by estimating the amount based on biomass accumulated , or by direct chemical measurement using a colorimetric assay Citation[173]. By regular monitoring, one could add Si when needed to promote high biomass accumulation without detrimental side effects.
A valid concern is what would be the added cost of including Si as a component of the growth medium. There is a lack of publically available information on Si’s projected costs; in nine cost analysis publications over the past 2 years, only one found one that mentioned the words Si or silica, and there were no cost figures for Si included Citation[141]. Our discussion is therefore limited to general concepts. First is the availability of precursor raw material; Si is the second most abundant element on the earth’s surface, and so supplies are not limited. The true cost will be in the conversion of precursors to silicic acid. A variety of types of silica ranging from crystalline quartz (e.g., sand) to amorphous diatomaceous earth will dissolve in water, with amorphous forms dissolving faster Citation[75]. Second is the cost related to the recycling of silica from spent medium or biomass. Because diatom silica readily dissolves at high pH Citation[75], recovery of silica from lipid-extracted diatoms should be feasible, without detrimental effects on other cellular components, such as protein. Therefore, recovered Si could be recycled repeatedly, since the chemical form of dissolved Si will not be altered, as might happen with other nutrients such as N. A third factor is the possibility of using diatom silica as a value-added product, which is discussed below. The overall cost of adding Si to a production system will depend on these factors and has to be balanced against potential increases in productivity using diatoms in a Si limitation approach. To date, no studies have addressed these issues in a quantitative way, which obviously needs to be done to evaluate the possibilities and to provide impetus for potentially improving processing methods to reduce costs.
Diatom silica generated in a production system can be considered as a marketable product. Diatomaceous earth is already widely used in commercial processes and products. Due to the porous nature of diatom frustules, they are excellent filtering materials for very fine particles, and are primarily used in pool filters and for filtration of beer and wine. Diatom silica is also highly hygroscopic, and it is used as an insecticide because its contact with the cuticle stimulates desiccation of the insect in conjunction with perforating the cuticle with sharp-edged small particles. It is also used as a neutral anthelmintic (dewormer) in mammals. The abrasive nature of microscopic diatom particles is taken advantage of in metal polishes and toothpastes. Diatom silica, which is essentially glass, has optical properties, and when added to paint, makes it flat. Diatomaceous earth is added to plastic wrap to separate the layers slightly to prevent them sticking tightly to each other. Dynamite is a stabilized form of nitroglycerin, which is absorbed into diatomaceous earth.
Mined diatomaceous earth is usually a mixture of diatom species, and so can be relatively heterogeneous. It is possible that diatom silica isolated from a monoculture pond may have more consistent properties. One possible future application of diatom silica is in nanotechnology. Diatoms make nanostructured silica in a variety of shapes that cannot be achieved by current synthetic materials approaches. Silica has limited properties as a material but several approaches have been developed by which diatom silica can either be directly converted or used as a template to generate a completely different chemistry while maintaining the nanoscale structure Citation[174].
Diatoms are likely to have other added-value applications. They are currently used as small percentage supplements to animal feed, and we are aware of studies evaluating the use of diatoms as a larger component of animal feed, ranging from fish to cattle. EPA from cultivated diatoms is less expensive than from fish sources Citation[175]. Other applications, such as amino acids for cosmetics, antibiotics and antiproliferative agents, are at early stages of development Citation[175].
Conclusion
A question commonly asked is, “If diatoms have such desirable properties and potential advantages, then why aren’t they more highly represented in biofuels research and production projects?” After considering their attributes as described in this article, it is puzzling. Two possible explanations come to mind. One is the long history of the study of chlorophytes due to their intimate relationship with terrestrial plants (especially related to photosynthesis) and our food supply. Another is related to the fact that diatoms dominate marine environments, whereas chlorophytes and plants dominate terrestrial environments. The ocean is a difficult system to study, and there are fewer research institutions with a marine focus than those dedicated to terrestrial environments; this imbalance may be an additional reason why the virtues of diatoms have been under appreciated. Generalized statements about the properties of ‘algae’ are also often heard. It is hoped that this review has highlighted the substantial differences not only between different classes of microalgae (specifically between diatoms and chlorophytes), but between different species within a class. Generalizations are not only inaccurate, but obscure the rich diversity of microalgae, which is a ‘value-added product’ in itself. At this early stage in the development of algal biofuels, little is known about the fundamental mechanisms involved in carbon fixation, flux and partitioning in microalgal cells, nor are scientists well versed in how to grow microalgae at scale. The deployment of algal biofuels on a production scale will involve largely uncontrollable and only partially predictable environmental conditions; thus, development of strains with diverse environmentally related productivity attributes will be necessary. For these reasons, it is valuable to investigate diverse classes of algae as research models and as potential production organisms. In a practical sense, this will define what is possible in terms of environmental adaptability and productivity, and allow a survey of the gene pool, which could be used to improve production species. As a class of algae, diatoms have all of the requisite attributes to be included as important contributors to the development of algal biofuels technology.
Future perspective
There has been substantial progress in the development of algal biofuels in the past few years. This may not yet be reflected in the publication record, because of the lag time in getting data published and corporate policies to protect data; however, at algal biofuels meetings it is clear that there is an undercurrent of excitement about things moving forward. The two major challenges in the development of this technology are the development of the “biology” to understand and improve factors that result in consistently high productivity of microalgal strains and development of large-scale culture systems that can grow algae and produce fuel precursors economically and at predictable production levels. Of the two challenges, understanding the biology is likely to be more straightforward, given the powerful tools already available to investigate and manipulate cellular metabolism. After an initial phase, over the next several years, of understanding and manipulating metabolic factors involved in fuel precursor synthesis in model species and initial production strains, a transition into applying these tools to a wider variety of production strains, which will be adapted to specific environmental conditions, is anticipated. This may also include multispecies cultivation, in which one or the other species is the primary producer under given conditions. This approach will require greater understanding of the ecological aspects of microalgal growth. Throughout the development of the organismal basis for biofuels production, an intimate interaction with the engineering and production aspects of large-scale cultivation will be needed.
The ability of a species to grow rapidly to high density and accumulate fuel precursor molecules.
Unicellular microalgae of the heterokont class, characterized by silica-based cell walls and high productivity.
The process of triacylglycerol accumulation in cells leading to the formation of lipid droplets.
The need by diatoms for an additional nutrient (silicon for cell wall synthesis) relative to other classes of algae. Silicon limitation will induce lipid accumulation in most diatoms.
The ability of a microalgal species to adapt and grow in different environmental conditions, which is relevant to the development of biofuels production systems.
Evolutionary aspects of diatoms
▪ Diatoms have a different evolutionary history than chlorophytes, which resulted in fundamental differences in cellular organization and metabolic processes. | |||||
▪ On geologic timescales diatoms have become the predominant carbon fixers in the ocean. | |||||
Diatom productivity
▪ Diatoms out-compete other classes of algae for growth, which may result from superior nutrient acquisition and storage and photosynthetic efficiency. | |||||
▪ Diatoms are environmentally adaptable and trophically flexible. | |||||
What cellular factors contribute to diatom productivity?
▪ Diatoms exhibit more efficient photosystem electron flux and increased selectivity for CO2 compared with other classes of algae. | |||||
▪ Diatoms have substantial differences in intracellular compartmentation, the presence and localization of metabolic pathways and forms of carbohydrate stored compared with chlorophytes. | |||||
▪ The rate of lipid accumulation is substantially improved under silicon limitation relative to nitrogen limitation, and numerous detrimental effects of nitrogen limitation are avoided. | |||||
▪ The silica cell wall may reduce carbon requirements and energy for cell wall synthesis. | |||||
Attributes of diatoms for biofuels production
▪ Diatoms have been cultivated for decades in outdoor open systems for aquaculture. | |||||
▪ Diatoms naturally grow under bloom conditions – maximum growth to high density. | |||||
▪ Diatoms accumulate abundant lipids, and lipid content may be underestimated unless based on an ash-free dry weight basis. | |||||
▪ Diatoms produce added-value products. | |||||
▪ The diatom silica cell wall may contribute to improved production through precise control of biomass accumulation and increased settling rates. | |||||
Perceived disadvantages of diatoms in large-scale cultivation
▪ Diatoms require silicon to grow, but the costs of supply, recycling and value-added products relative to productivity have not been determined. | |||||
▪ Abundant silicon supply exists, and recycling should be feasible. | |||||
Financial & competing interests disclosure
Diatom biofuels research in the Hildebrand laboratory is supported by AFOSR grant FA9550-08-1-0178, DOE grant DE-EE0001222 and NSF grant CBET-0903712. SRS was supported by the Department of Defense through the National Defense Science and Engineering Graduate Fellowship Program. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed.
No writing assistance was utilized in the production of this manuscript.
References
- Pienkos PT, Darzins A. The promise and challenges of microalgal-derived biofuels. Biofuels Bioprod. Bioref. Bioref.3,431–440 (2009).
- Trequer P, Nelson DM, Van Bennekom AJ, DeMaster DJ, Leynaert A, Queguiner B. The silica balance in the world ocean: a reestimate. Science268,375–379 (1995).
- Field CB, Behrenfeld MJ, Randerson JT, Falkowski P. Primary production of the biosphere, integrating terrestrial and oceanic components. Science281,237–240 (1998).
- Kooistra WHCF, Gersonde R, Medlin LK, Mann DG. The origin and evolution of the diatoms. In: Evolution of Primary Producers in the Sea. Falkowski PG, Knoll AH (Eds). Elsevier, Boston, MA, USA, 207–249 (2007).
- Armbrust EV. The life of diatoms in the world’s oceans. Nature459,185–192 (2009).
- Becker B, Hoef-Emden K, Melkonian M. Chlamydial genes shed light on the evolution of photoautotrophic eukaryotes. BMC Evol. Biol.8,203–220 (2008).
- Moustafa A, Beszteri B, Maier UG, Bowler C, Valentin K, Bhattacharya D. Genomic footprints of a cryptic plastid endosymbiosis in diatoms. Science324,1724–1726 (2009).
- Bowler C, Allen AE, Badger JH et al. The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature456,239–244 (2008).
- Finazzi G, Moreau H, Bowler C. Genomic insights into photosynthesis in eukaryotic phytoplankton. Trends Plant Sci.15,565–572 (2010).
- Armbrust E, Berges J, Bowler C et al. The genome of the diatom Thalassiosira pseudonana, ecology, evolution, and metabolism. Science306(5693),79–86 (2004).
- Wilhelm C, Buchel C, Fisahn J et al. The regulation of carbon and nutrient assimilation in diatoms is significantly different from green algae. Protist157,91–124 (2006).
- Guidry MW, Arvidson RS, MacKenzie FT. Biological and geochemical forcings to phanerozoic change in seawater, atmosphere, and carbonate precipitate composition. In: Evolution of Primary Producers in the Sea. Falkowski PG, Knoll AH (Eds). Elsevier, Boston, MA, USA, 377–403 (2007).
- Rabosky DL, Sorhannus U. Diversity dynamics of marine planktonic diatoms across the Cenozoic. Nature457,183–186 (2009).
- Falkowski PG, Katz ME, Knoll AH et al. The evolution of modern eukaryotic phytoplankton. Science305,354–360 (2004).
- Falkowski PG, Katz ME, Milligan AJ et al. The rise of oxygen over the past 205 million years and the evolution of large placental mammals. Science309,2202–2204 (2005).
- Emery KO, Uchupi E. Western North Atlantic ocean. Am. Assoc. Pet. Geol. Mem.17,532 (1972).
- Smetacek V. Diatoms and the ocean carbon cycle. Protist150,25–32 (1999).
- Dugdale RC, Wilkerson FP. Silicate regulation of new production in the equatorial pacific upwelling. Nature391,270–273 (1998).
- Raven JA. The role of vacuoles. New Phytol.106,357–422 (1987).
- Sicko-Goad L, Stoermer EF, Ladewski BG. A morphometric method for correcting phytoplankton cell volume estimates. Protoplasma93,147–163 (1977).
- Eppley RW, Rogers J. Inorganic nitrogen assimilation of Ditylumbrightwellii, a marine plankton diatom. J. Phycol.6,344–350 (1970).
- Furnas MJ. In situ growth rates of marine phytoplankton – approaches to measurement, community, and species growth rates. J. Plankton Res.12,1117–1151 (1990).
- Amano Y, Takahashi K, Machida M. Competition between the cyanobacterium Microcystis aeruginosa and the diatom Cyclotella sp. under nitrogen-limited condition caused by dilution in eutrophic lake. J. Appl. Phycol. doi:10.1007/s10811–011–9718–8 (2011) (Epub ahead of print).
- Tozzi S, Schofield O, Falkowski P. Historical climate change and ocean turbulence as selective agents for two key phytoplankton functional groups. Mar. Ecol. Prog. Ser.274,123–132 (2004).
- Grover JP. Dynamics of competition in a variable environment – experiments with 2 diatom species. Ecology69,408–417 (1988).
- Crowley TJ, North GR. Paleoclimatology. Oxford University Press, NY, USA, 349 (1991).
- Prauss M. The oceanographic and climatic interpretation of marine palynomorph phytoplankton distribution from Mesozoic, Cenozoic and Recent sections. Göttinger Arb.Geol. Paläont.76,1–235 (2000).
- Huber BT, Hodell DA, Hamilton CP. Middle–late cretaceous climate of the southern high-lattitudes – stable isotopic evidence for minimal equator-to-pole thermal-gradients. Geol. Soc. Am. Bull.107,1164–1191 (1995).
- Falkowski PG, Oliver MJ. Mix and max, how climate selects phytoplankton. Nat. Rev. Microbiol.5,813–819 (2007).
- Thomas WH, Dodson AN, Reid FMH. Diatom productivity compared with other algae in natural marine phytoplankton assemblages. J. Phycol.14,250–253 (1978).
- Thomas WH, Seibert DLR, Dodson AN. Phytoplankton enrichment experiments and bioassays in natural coastal seawater and in sewage outfall receiving waters off Southern California. Estuar. Coast Mar. Sci.2,191–206 (1974).
- Jitts HR, McAllister CD, Stephens K, Strickland JDH. The cell division rates of some marine phytoplankton as a function of light and temperature. J. Fish. Res. Board Can.21,139–157 (1964).
- McAllister CD, Shah N, Strickland JDH. Marine phytoplankton photosynthesis as a function of light intensity, a comparison of methods. J. Fish. Res. Board Can.21,159–181 (1964).
- Smetacek V. Diatoms and the ocean carbon cycle. Protist150,25–32 (1999).
- Wagner H, Jakob T, Wilhelm C. Balancing the energy flow from captured light to biomass under fluctuating light. New Phytol.169,95–108 (2006).
- Franklin DJ, Brussaard CPD, Berges JA. What is the role and nature of programmed cell death in phytoplankton ecology? Eur. J. Phycol.41,1–14 (2006).
- Kirchman DL. Phytoplankton death in the sea. Nature398,293–294 (1999).
- Parrish CC. Time series of particulate and dissolved lipid classes during spring phytoplankton blooms in Bedford Basin, a marine inlet. Mar. Ecol. Prog. Ser.35,129–139 (1987).
- Nichols PD, Palmisano AC, Rayner MS, Smith GA, White DC. Changes in the lipid composition of Antarctic sea-ice diatom communities during a spring bloom, an indication of community physiological status. Antarct. Sci.1,133–140 (1989).
- Lebeau T, Robert JM. Diatom cultivation and biotechnologically relevant products. Part 1: cultivation at various scales. Appl. Microbiol. Biotech.60,612–623 (2003).
- Huesemann MH, Hausmann TS, Bartha R, Aksoy M, Weissman JC, Benemann JR. Biomass productivities in wild type and pigment mutant of Cyclotella sp. (Diatom). Appl. Biochem. Biotech.157,507–526 (2009).
- Goldman JC, Ryther JH, Williams LD. Mass production of marine algae in outdoor cultures. Nature245,594–595 (1975).
- D’Elia CF, Ryther JH, Losordo TM. Productivity and nitrogen balance in large scale phytoplankton cultures. Water Res.11,1031–1040 (1977).
- Boussiba S, Vonshak A, Cohen Z, Avissar Y, Richmond A. Lipid and biomass production by the halotolerant microalga Nannochloropsis salina.Biomass12,37–47 (1987).
- Richmond A, Lichtenberg E, Stahl B, Vonshak A. Quantitative assessment of the major limitations on productivity of Spirulina platensis in open raceways. J. Appl. Phycol.2,195–206 (1990).
- Doucha J, Livansky K. Productivity, CO2/O2 exchange and hydraulics in outdoor open high density microalgal (Chlorella sp.) photobioreactors operated in a middle and southern European climate. J. Appl. Phycol.18,811–826 (2006).
- Lohr M, Wilhelm C. Algae displaying the diadinoxanthin cycle also possess the violaxanthin cycle. Proc. Natl Acad. Sci. USA96,8784–8789 (1999).
- Wagner H, Jakob T, Wilhelm C. Balancing the energy flow from captured light to biomass under fluctuating light conditions. New Phytol.169,95–108 (2006).
- Wollman FA. State transitions reveal the dynamics and flexibility of the photosynthetic apparatus. EMBO J.20,3623–3630 (2001).
- Ting CS, Owens TG. Photochemical and non-photochemical fluorescence quenching processes in the diatom Phaeodactylum tricornutum.Plant Physiol.101,1323–1330 (1986).
- Berkaloff C, Caron L, Rousseau B. Subunit organization of PS I particles from brown algae and diatoms, polypeptide and pigment analysis. Photosynth. Res.23,181–193 (1990).
- Owens TG. Light-harvesting antenna systems in the chlorophyll a/c-containing algae. In: Light-Energy Transduction in Photosynthesis. Stevens CLR, Bryant DA (Eds). American Society of Plant Physiologists, Rockville, MA, USA, 122–136 (1988).
- Larkum AWD, Vesk M. Algal plastids, their fine structure and properties. Photosynthesis Algae. Larkum AW, Douglas SE, Raven JA (Eds). Kluwer Academic Publishers, Dordrecht, The Netherlands, 11–28 (2003).
- Westermann M, Rhiel E. Localisation of fucoxanthin chlorophyll a/c-binding polypeptides of the centric diatom Cyclotella cryptica by immunoelectron microscopy. Protoplasma225,217–223 (2005).
- Michels A, Wedel N, Kroth PG. Diatom plastids possess a phosphoribulokinase with an altered regulation and no oxidative pentose phosphate pathway. Plant Physiol.137,911–920 (2005).
- Badger MR, Andrews TJ, Whitney SM et al. The diversity and coevolution of RuBisCO, plastids, pyrenoids, and chloroplast-based CO2-concentrating mechanisms in algae. Can. J. Bot.76,1052–1071 (1998).
- Giordano M, Beardall J, Raven JA. CO2 concentrating mechanisms in algae. Mechanisms, environmental modulation, and evolution. Annu. Rev. Plant Biol.56,99–131 (2005).
- Roberts K, Granum E, Leegood RC, Raven JA. Carbon acquisition by diatoms. Photosynth. Res.93,79–88 (2007).
- Tachibana M, Allen AE, Kikutani S, Endo Y, Bowler C, Matsuda Y. Localization of putative carbonic anhydrases in two marine diatoms, Phaeodactylum tricornutum and Thalassiosira pseudonana.Photosynth. Res.109,205–221 (2011).
- Wirtz KW. Non-uniform scaling in phytoplankton growth rate due to intracellular light and CO2 decline. J. Plankton Res.33,1325–1341 (2011).
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular Biology of the Cell (4th Edition). Garland Science, NY, USA (2002).
- Lang M, Apt KE, Kroth PG. Protein transport into “complex” diatom plastids utilizes two different targeting signals. J. Biol. Chem.273,30973–30978 (1998).
- Gibbs SJ. The route of entry of cytoplasmically synthesized proteins into chloroplasts of algae possessing chloroplast ER. Cell Sci.35,253–266 (1979).
- Gruber A, Vugrinec S, Hempel F, Gould SB, Maier U, Kroth PG. Protein targeting into complex diatom plastids, function and characterization of a specific targeting motif. Plant Mol. Biol.64,519–530 (2007).
- Moog D, Stork S, Zauner S, Maier UG. In silico and in vivo investigations of proteins of a minimized eukaryotic cytoplasm. Genome Biol. Evol.3,375–382 (2011).
- Kroth PG, Chiovitti A, Gruber A et al. A model for carbohydrate metabolism in the diatom Phaeodactylum tricornutum deduced from comparative whole genome analysis. PLoS ONE3,e1426 (2008).
- Graham JWA, Williams TCR, Morgan M, Fernie AR, Ratcliffe RG, Sweetlove LJ. Glycolytic enzymes associate dynamically with mitochondria in response to respiratory demand and support substrate channeling. Plant Cell19,3723–3738 (2007).
- Allen AE, Dupont CL, Obornik M et al. Evolution and metabolic significance of the urea cycle in photosynthetic diatoms. Nature473,203–209 (2011).
- Beattie A, Percival E, Hirst EL. Studies on metabolism of chrysophyceae – comparative structural investigations on leucosin. (chyrsolaminarin) separated from diatoms and laminarin from brown algae. Biochem. J.79,531–537 (1961).
- Chiovitti A, Molino P, Crawford SA, Teng RW, Spurck T, Wetherbee R. The glucans extracted with warm water from diatoms are mainly derived from intracellular chrysolaminaran and not extracellular polysaccharides. Eur. J. Phycol.39,117–128 (2004).
- Libessart N, Maddelein ML, Van den Koornhuyse N et al. Storage, photosynthesis, and growth – the conditional nature of mutations affecting starch synthesis and structure in Chlamydomonas.Plant Cell7,1117–1127 (1995).
- Pickett-Heaps J, Schmid AMM, Edgar LA. The cell biology of diatom valve formation. Progress in Phycological Research. Round FE, Chapman DJ (Eds). Biopress Ltd., Bristol, UK, 1–168 (1990).
- Del Amo Y, Brzezinski MA. The chemical form of dissolved Si taken up by marine diatoms. J. Phycol.35,1162–1170 (1999).
- Thamatrakoln K, Hildebrand M. Silicon uptake in diatoms revisited, a model for saturable and nonsaturable uptake kinetics and the role of silicon transporters. Plant Physiol.146,1397–1407 (2008).
- Iler RK. The chemistry of silica, solubility, polymerization, colloid and surface properties, and biochemistry. Wiley-Interscience, NY, USA 866 (1979).
- Raven JA. The transport and function of silicon in plants. Biol. Rev.58,179–207 (1983).
- Milligan AJ, Morel FMM. A proton buffering role for silica in diatoms. Science297,1848–1850 (2002).
- Brennan L, Owende P. Biofuels from microalgae – a review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev.14,557–577 (2010).
- Clavero E, Hernandez-Marine M, Grimalt JO, Garcia-Pichel F. Salinity tolerance of diatoms from thalassic hypersaline environments. J. Phycol.36,1021–1034 (2000).
- Kraebs G, Buechel C. Temperature and salinity tolerances of geographically separated Phaeodactylum tricornutum Bohlin strains, maximum quantum yield of primary photochemistry, pigmentation, proline content and growth. Bot. Mar.54,231–241 (2011).
- Gerhardt A, de Bisthoven LJ, Guhr K, Soares AMVM, Pereira MJ. Phytoassessment of acid mine drainage, Lemna gibba bioassay and diatom community structure. Ecotoxicology17,47–58 (2008).
- Souza-Egipsy V, Altamirano M, Amils R, Aguilera A. Photosynthetic performance of phototrophic biofilms in extreme acidic environments. Env. Microbiol.13,2351–2358 (2011).
- Hecky RE, Kilham P. Diatoms in alkaline, saline lakes – ecology and geochemical implications. Limnol. Oceanog.18,53–71 (1973).
- Bryanskaya AV, Namsaraev ZB, Kalashnikova OM, Barkhutova DD, Namsaraev BB, Gorlenko VM. Biogeochemical processes in algal-bacterial mats of the Urinskii alkaline hot spring. Micrbiology75,611–620 (2006).
- Tolomio C, Ceschi-Berrini C, Moschin E, Galzigna L. Colonization by diatoms and antirheumatic activity of thermal mud. Cell Biochem. Funct.17,29–33 (1999).
- Choi HG, Joo HM, Jung W, Hong SS, Kang JS, Kang SH. Morphology and phylogenetic relationships of some psychrophilic polar diatoms (Bacillariophyta). Nova Hedw.133,7–30 (2008).
- Neori A, Holm-Hansen O. Effect of temperature on rate of photosynthesis in Antarctic phytoplankton. Polar Biol.1,33–38 (1982).
- Thomas DN, Dieckmann GS. Ocean science – Antarctic Sea ice – a habitat for extremophites. Science295,641–644 (2002).
- Zielinski U, Gersonde R. Diatom distribution in southern ocean surface sediments. (Atlantic sector), implications for paleoenvironmental reconstructions. Paleogeo. Paleoclimat. Paleoecol.129,213–250 (1997).
- Mock T, Hoch N. Long-term temperature acclimation of photosynthesis in steady-state cultures of the polar diatom Fragilariopsis cylindrus.Photosynth. Res.85,307–317 (2005).
- Hellebust JA. Glucose uptake by Cyclotella cryptica – dark induction and light inactivation of transport system. J. Phycol.7,345–349 (1971).
- Ceron Garcia M, Garcia Camacho F, Sanchez Miron A, Fernandez Sevilla JM, Molina Grima E. Mixotrophic production of marine microalga Phaeodactylum tricornutum on various carbon sources. J. Microbiol. Biotechnol.16,689–694 (2006).
- Lewin JC, Lewin RA. Auxotrophy and heterotrophy in marine littoral diatoms. Can. J. Microbiol.6,127–137 (1960).
- Lewin JC, Lewin RA. Culture and nutrition of some apochlorotic diatoms of the genus Nitzschia.J. Gen. Microbiol.46,361–367 (1967).
- Armstrong E, Rogerson A, Weftley JW. Utilisation of seaweed carbon by three surface associated heterotrophic protists, Stereomyxaramosa, Nitzschia alba and Labyrinthula sp. Aquat. Microb. Ecol.200,49–57 (2000).
- Linkins AE. Uptake and utilization of glucose and acetate by a marine chemoorganotrophic diatom Nitzschia alba, clone link 001. PhD Thesis. University of Massachusetts, MA, USA (1973).
- Shifrin NS, Chisholm SW. Phytoplankton lipids, interspecific differences and effects of nitrate, silicate, and light-dark cycles. J. Phycol.17,374–384 (1981).
- Hu Q, Sommerfeld M, Jarvis E et al. Microalgal triacylglycerols as feedstocks for biofuel production, perspectives and advances. Plant J.54,621–639 (2008).
- Griffiths MJ, Harrison STL. Lipid productivity as a key characteristic for choosing algal species for biodiesel production. J. Appl. Phycol.21,493–507 (2009).
- Yu ET, Zendejas FJ, Lane PD, Gaucher S, Simmons BA, Lane TW. Triacylglycerol accumulation and profiling in the model diatoms Thalassiosira pseudonana and Phaeodactylum tricornutum (Baccilariophyceae) during starvation. J. Appl. Phycol.21,669–681 (2009).
- Guckert JB, Cooksey KE. Triglyceride accumulation and fatty acid profile changes in Chlorella (Chlorophyta) during high pH-induced cell cycle inhibition. J. Phycol.26,72–79 (1990).
- Thompson GA. Lipids and membrane function in green algae. Biochim. Biophys. Acta1302,17–45 (1996).
- Chen GQ, Jiang Y, Chen F. Salt-induced alterations in lipid composition of diatom Nitzschia laevis under heterotrophic culture condition. J. Phycol.44,1309–1314 (2008).
- Mansour MP, Frampton DMF, Nichols PD, Volkman JK, Blackburn SI. Lipid and fatty acid yield of nine stationary-phase microalgae, applications and unusual C-24-C-28 polyunsaturated fatty acids. J. Appl. Phycol.17,287–300 (2005).
- Radakovits R, Eduafo PM, Posewitz MC. Genetic engineering of fatty acid chain length in Phaeodactylum tricornutum.Metab. Eng.13,89–95 (2011).
- Knothe G. “Designer” biodiesel, optimizing fatty ester composition to improve fuel properties. Energy Fuels22,1358–1364 (2008).
- Correa-Reyes JG, del Pilar Sanchez-Saavedra M, Viana Maria T, Flores-Acevedo N, Vasquez-Pelaez C. Effect of eight benthic diatoms as feed on the growth of red abalone (Haliotisrufescens) postlarvae. J. Appl. Phycol.21,387–393 (2009).
- Lewin JC, Guillard RR. Diatoms. Ann. Rev. Microbiol.17,373–414 (1963).
- Roessler PG. Effects of silicon deficiency on lipid composition and metabolism in the diatom Cyclotella cryptica.J. Phycol.24,394–400 (1988).
- Bertrand M. Carotenoid biosynthesis in diatoms. Photosynth. Res.106,89–102 (2010).
- Jin E, Polle JEW, Lee HK, Hyun SM, Chang M. Xanthophylls in microalgae, from biosynthesis to biotechnological mass production and application. J. Microbiol. Biotechnol.13,165–174 (2003).
- Lichtenthaler HK. The 1-deoxy-D-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants. Annu. Rev. Plant Phys. Plant Mol. Biol.50,47–65 (1999).
- Rohmer M. The discovery of a mevalonate-independent pathway for isoprenoid biosynthesis in bacteria, algae and higher plants. Nat. Prod. Rep.16,565–574 (1999).
- Lincoln RA, Strupinski K, Walker JM. Biologically active compounds from diatoms. Diatom. Res.5,337–349 (1990).
- Pesando D. Antibacterial and antifungical activities of marine algae. Introduction to Applied Phycology. Akatsuka I (Ed.). Academic Publishing, NY, USA 3–26 (1990).
- Viso AC, Pesando D, Baby C. Antibacterial and antifungical properties of some marine diatoms in culture. Bot. Mar.30,41–45 (1987).
- Imada N, Kobayashi K, Isdmura K et al. Isolation and identification of an autoinhibitor produced by Skeletonema costatum.Nippon Suisan Gakkaishi58,1687–1692 (1992).
- Naviner M, Berge JP, Durand P, Le Bris H. Antibacterial activity of the marine diatom Skeletonema costatum against aquacultural pathogens. Aquaculture174,15–24 (1999).
- Berge JP, Bourgougnon N, Carbonnelle D et al. Antiproliferative effects of an organic extract from the marine diatom Skeletonema costatum (Grev.) Cleve against a non-small-cell bronchopulmonary carcinoma line (NSCLC-N6). Anticancer Res.17,2115–2120 (1997).
- Berge JP, Bourgougnon N, Pojer F et al. Antiviral and anticoagulant activities of a water-soluble fraction of the marine diatom Hasleaostrearia.Plant Med.65,604–609 (1999).
- Rowland SJ, Belt ST, Wraige EJ, Masse G, Roussakis C, Robert JM. Effects of temperature on polyunsaturation in cytostatic lipids of Hasleaostrearia.Phytochemistry56,597–602 (2001).
- Wraige EJ, Johns L, Belt SJ, Masse G, Robert JM, Rowland SJ. Highly branched C25 isoprenoids in axenic cultures of Hasleaostrearia.Phytochemistry51,69–73 (1999).
- Prestegard SK, Oftedal L, Coyne RT et al. Marine benthic diatoms contain compounds able to induce leukemia cell death and modulate blood platelet activity. Mar. Drugs7,605–623 (2009).
- Wang ZT, Ullrich N, Joo S, Waffenschmidt S, Goodenough U. Algal lipid bodies, stress induction, purification, and biochemical characterization in wild-type and starchless Chlamydomonas reinhardtii.Eukaryot. Cell8,1856–1868 (2009).
- Work VH, Ullrich N, Joo S, Waffenschmidt S, Goodenough U. Increased lipid accumulation in the Chlamydomonas reinhardtii sta7–10 starchless isoamylase mutant and increased carbohydrate synthesis in complemented strains. Eukaryot. Cell9,1251–1261 (2010).
- Deng X, Fei X, Li Y. The effects of nutritional restriction on neutral lipid accumulation in Chlamydomonas and Chlorella.Afr. J. Microbiol. Res.5,260–270 (2011).
- Turpin DH. Effects of inorganic N availability on algal photosynthesis and carbon metabolism. J. Phycol.27,14–20 (1991).
- Berges JA, Charlebois DO, Mauzerall DC, Falkowski PG. Differential effects of nitrogen limitation on photosynthetic efficiency of photosystems I and II in microalgae. Plant Physiol.110,689–696 (1996).
- Plumley FG, Schmidt GW. Nitrogen-dependent regulation of photosynthetic gene expresssion. Proc. Natl Acad. Sci. USA86,2678–2682 (1989).
- Healy FP, Coombs J, Volcani BE. Changes in pigment content of the diatom Navicula pelliculosa (Bréb.) Hilse in silicon-starvation synchrony. Arch. Mikrobiol.59,131–142 (1967).
- Martin NC, Chiang KS, Goodenough UW. Turnover of chloroplast and cytoplasmic ribosomes during gametogenesis in Chlamydomonas reinhardtii.Dev. Biol.51,190–201 (1976).
- Geider RJ, LaRoche J, Greene R, Molaizola M. Response of the photosynthetic apparatus of Phaeodactylum tricornutum to nitrate, phosphate or iron starvation. J. Phycol.29,755–766 (1993).
- Roessler PG. Changes in the activities of various lipid and carbohydrate biosynthetic enzymes in the diatom Cyclotella cryptica in response to silicon deficiency. Arch. Biochem. Biophys.267,521–528 (1988).
- Berges JA, Falkowski PG. Physiological stress and cell death in marine phytoplankton, induction of proteases in response to nitrogen or light limitation. Limnol. Oceanog.43,129–135 (1998).
- Claquin P, Martin-Jezequel V. Uncoupling of silicon compared with carbon and nitrogen metabolisms and the role of the cell cycle in continuous cultures of Thalassiosira pseudonana (Bacillariophyceae) under light, nitrogen, and phosphorus control. J. Phycol.38,922–930 (2002).
- Darley WM, Volcani BE. Role of silicon in diatom metabolism, a silicon requirement for deoxyribonucleic acid synthesis in the diatom Cylindrothecafusiformis Reimann and Lewin. Exp. Cell Res.58,334–342 (1969).
- McGinnis KM, Dempster TA, Sommerfeld MR. Characterization of the growth and lipid content of the diatom Chaetoceros muelleri.J. Appl. Phycol.9,19–24 (1997).
- Schlechtriem C, Focken U, Becker K. Effect of different lipid extraction methods on δ C-13 of lipid and lipid-free fractions of fish and different fish feeds. Isotopes Environ. Health Stud.39,135–140 (2003).
- Hildebrand M, Dahlin K. Nitrate transporter genes from the diatom Cylindrothecafusiformis. (Bacillariophyceae), mRNA levels controlled by nitrogen source and by the cell cycle. J. Phycol.36,702–713 (2000).
- Shoaf WT, Lium BW. Improved extraction of chlorophyll a and b from algae using dimethyl sulfoxide. Limnol. Oceanog.21,926–928 (1976).
- Greenwell HC, Laurens LML, Shields RJ, Lovitt RW, Flynn KJ. Placing microalgae on the biofuels priority list, a review of the technological challenges. J. R. Soc. Interface7,703–726 (2010).
- Scharek R, Tupas LM, Karl DM. Diatom fluxes to the deep sea in the oligotrophic North Pacific gyre at Station ALOHA. Mar. Ecol. Prog. Ser.182,55–67 (1999).
- Joh G, Choi YS, Shin JK, Lee J. Problematic algae in the sedimentation and filtration process of water treatment plants. J. Water Suppl. Res. Technol.60,219–230 (2011).
- Borges L, Moron-Villarreyes JA, Montes D’Oca MG, Abreu PC. Effects of flocculants on lipid extraction and fatty acid composition of the microalgae Nannochloropsisoculata and Thalassiosira weissflogii.Biomass Bioenerg.35,4449–4454 (2011).
- Alldredge AL, Gotschalk CC. Direct observations of the mass flocculation of diatom blooms, characteristics, settling velocities and formation of diatom aggregates. Deep Sea Res. A Oceanogr. Res. Pap.36,159–171 (1989).
- Logan BE, Passow U, Alldredge AL. Variable retention of diatoms on screens during size separations. Limnol. Oceanog.39,390–395 (1994).
- Kiorboe T, Andersen KP, Dam HG. Coagulation efficiency and aggregate formation in marine phytoplankton. Mar. Biol.107,235–245 (1990).
- Smetacek VS. Role of sinking diatom life history cycles, ecological, evolutionary, and geological significance. Mar. Biol.84,239–251 (1985).
- Margalef R. Turbulence and marine life. Scient. Mar.61(Supp. 1),109–123 (1997).
- Contreras A, Garcia F, Molina E, Merchuk JC. Interaction between CO2-mass transfer, light availability, and hydrodynamic stress in the growth of Phaeodactylum tricornutum in a concentric tube airlift photobioreactor. Biotechnol. Bioeng.60,317–325 (1998).
- Camacho FG, Grima EM, Miron AS, Pascual VG, Chisti Y. Carboxymethyl cellulose protects algal cells against hydrodynamic stress. Enzyme Microb. Technol.29,602–610 (2001).
- Lebeau T, Robert JM. Diatom cultivation and biotechnologically relevant products. Part 1, cultivation at various scales. Appl. Microbiol. Biotechnol.60,612–623 (2003).
- Vandanjon L, Rossignol N, Jaouen P, Robert JM, Quemeneur F. Effects of shear on two microalgae species. Contribution of pumps and valves in tangential flow filtration systems. Biotechnol. Bioeng.63,1–9 (1999).
- Michels MHA, van der Goot AJ, Norsker NH, Wijffels RH. Effects of shear stress on the microalgae Chaetoceros muelleri.Bioproc. Biosys. Engin.33,921–927 (2010).
- Pohnert G. Diatom/copepod interactions in plankton, the indirect chemical defense of unicellular algae. ChemBioChem6,946–959 (2005).
- Hamm CE, Merkel R, Springer O et al. Architecture and material properties of diatom shells provide effective mechanical protection. Nature421,841–843 (2003).
- Shaw BA, Harrison PJ, Andersen RJ. Feeding deterrence properties of apo-fucoxanthinoids from marine diatoms. 2: physiology of production of apo-fucoxanthinoids by the marine diatoms Phaeodactylum tricornutum and Thalassiosira pseudonana, and their feeding deterrent effects on the copepod Tigriopuscalifornicus.Mar. Biol.124,473–481 (1995).
- Kuhn SF. Infection of Coscinodiscus spp. by the parasitoid nanoflagellate Pirsonia diadema. 1: behavioural studies on the infection process. J. Plankton Res.19,791–804 (1997).
- Rasconi S, Jobard M, Sime-Ngando T. Parasitic fungi of phytoplankton, ecological roles and implications for microbial food webs. Aquat. Microb. Ecol.62,123–137 (2011).
- Neuhauser S, Kirchmair M, Gleason FH. Ecological roles of the parasitic phytomyxids (plasmodiophorids) in marine ecosystems – a review. Mar. Freshwater Res.62,365–371 (2011).
- Nagasaki K, Tomaru Y, Katanozaka N et al. Isolation and characterization of a novel single-stranded RNA virus infecting the bloom-forming diatom Rhizosoleniasetigera.Appl. Environ. Microbiol.70,704–711 (2004).
- Tomaru Y, Fujii N, Oda S, Toyoda K, Nagasaki K. Dynamics of diatom viruses on the western coast of Japan. Aquat. Microb. Ecol.63,223–230 (2011).
- Zaslavskaia LA, Lippmeier JC, Kroth PG, Grossman AR, Apt KE. Transformation of the diatom Phaeodactylum tricornutum with a variety of selectable marker and reporter genes. J. Phycol.36,379–386 (2000).
- Cerutti H, Johnson AM, Gillham NW, Boynton JE. Epigenetic silencing of a foreign gene in nuclear transformants of Chlamydomonas.Plant Cell9,925–945 (1997).
- Dunahay TG, Jarvis EE, Roessler PG. Genetic transformation of the diatoms Cyclotella cryptica and Naviculasaprophila.J. Phycol.31,1004–1012 (1995).
- Apt KE, Kroth-Pancic PG, Grossman AR. Stable nuclear transformation of the diatom Phaeodactylum tricornutum.Mol. Gen. Genet.252,572–579 (1996).
- Poulsen N, Kröger N. A new molecular tool for transgenic diatoms – control of mRNA and protein biosynthesis by an inducible promoter-terminator cassette. FEBS J.272,3413–3423 (2005).
- De Riso V, Raniello R, Maumus F, Rogato A, Bowler C, Falciatore A. Gene silencing in the marine diatom Phaeodactylum tricornutum.Nucleic Acids Res.37,e96 (2009).
- Kuhn SF, Brownlee C. Membrane organisation and dynamics in the marine diatom Coscinodiscus wailesii.Bot. Mar.48,297–305 (2005).
- Gruber A, Weber T, Bartulos CR, Vugrinec S, Kroth PG. Intracellular distribution of the reductive and oxidative pentose phosphate pathways in two diatoms. J. Basic Microbiol.49,58–72 (2009).
- Kroth PG, Schroers Y, Kilian O. The peculiar distribution of Class 1 and Class 2 aldolases in diatoms and in red algae. Curr. Genet.48,389–400 (2005).
- Hildebrand M, Frigeri L, Davis AK. Synchronized growth of Thalassiosira pseudonana. (Bacillariophyceae) provides novel insights into cell wall synthesis processes in relation to the cell cycle. J. Phycol.43,730–740 (2007).
- Strickland JDH, Parsons TR. A practical handbook of sea water analysis. Bull. Fish. Res. Board Can.167,1–311 (1968).
- Sandhage KH, Allan SM, Dickerson MB et al. Merging biological self-assembly with synthetic chemical tailoring, the potential for 3-D genetically-engineered micro/nano-devices. (3-D GEMS). Int. J. Appl. Ceram. Tech.2,317–326 (2005).
- Lebeau T, Robert JM. Diatom cultivation and biotechnologically relevant products. Part II: current and putative products. Appl. Microbiol. Biotech.60,624–632 (2003).
- Parker MS, Mock T, Armbrust EV. Genomic insights into marine microalgae. Annu. Rev. Genet.42,619–645 (2008).
▪ Website
- Aquatic species program close-out report. www.nrel.gov/docs/legosti/fy98/24190.pdf